固醇類物質對鵝顆粒細胞膽固醇轉運相關基因表達的影響
2019-03-11 05:23:54榮玉靜胡深強王繼文
四川農業大學學報 2019年1期
榮玉靜,夏 露,胡深強,王繼文
(四川農業大學動物科技學院,成都611130)
禽類卵巢中卵泡數量眾多,根據發育階段和功能不同形成了嚴格的等級體系。產蛋期家禽卵泡按排卵順序可分為等級卵泡和等級前卵泡,其中等級卵泡按大小和排卵順序分為F1、F2、F3、F4、F5[1],F1最接近排卵。顆粒細胞在卵泡發育過程中發揮重要作用,廣泛參與營養攝入[2],卵泡膜細胞分化[3]和類固醇激素生成[4]。類固醇激素是卵母細胞和顆粒細胞存活或凋亡的重要調控因子,和家禽繁殖性能相關的類固醇激素主要有孕激素、雌激素和雄激素。對卵巢起調控作用的主要孕激素是孕酮,當卵泡發育接近成熟時,顆粒細胞分泌大量孕酮促進排卵[5]。睪酮屬于雄激素,是雌二醇合成的必須底物,而雌二醇是家禽卵巢中生物活性最強的雌激素,可促進卵泡及卵母細胞發育成熟。
外部環境和禽體內分泌的刺激使下丘腦分泌促性腺激素釋放激素(GnRH),GnRH促進垂體分泌卵泡刺激素(FSH)和促黃體生成素(LH)作用于卵巢和卵泡上相應受體,啟動細胞內類固醇激素的合成,此過程由多個基因共同參與。膽固醇是類固醇激素合成的前體物質,它在細胞內的分布情況對膽固醇穩態以及類固醇激素的合成非常重要。固醇調節元件結合蛋白(sterol regulatory element-binding protein,SREBPs)屬于膜結合轉錄因子家族,成員SREBP-2主要調控膽固醇合成,是維持膽固醇平衡必需基因[6],其活性主要受細胞固醇含量調控。類固醇激素急性調節脂質轉運結構域4(START domain containing 4,STARD4)屬于START家族,主要受膽固醇和SREBP-2特異性調控[7]。STARD4與膽固醇在細胞內的跨細胞質轉運有關,在將膽固醇遞送至內質網的過程中發揮重要作用[8]。類固醇激素合成急性蛋白(steroidogenic acute regulatory protein,STAR)位于線粒體膜,是膽固醇從線粒體外膜轉運到內膜的限速酶,隨后膽固醇側鏈裂解酶(cholesterol side-chain cleavageenzyme,CYP11A1)將內膜上的膽固醇轉化為孕烯醇酮,作為合成孕酮的底物。
作為調控機體膽固醇轉運與類固醇激素合成的關鍵基因,SREBP-2、STADR4、STAR和CYP11A1在鵝卵泡發育過程中的作用及其調控機制尚不清楚。因此,本研究選取四川白鵝為研究對象,分析四個基因在鵝卵泡發育過程中的表達模式和不同固醇類物質對其轉錄水平的調控,以期為揭示鵝卵泡發育機制和提高產蛋量提供理論參考。
1 材料和方法
1.1 樣品采集
實驗動物來自四川農業大學家禽育種場。挑選4只同期孵化,相同飼養環境并處于產蛋高峰期的天府肉鵝母系母鵝。放血法處死后迅速取出卵巢,按以下標準分離出各個發育階段卵泡:等級卵泡F1、F2、F3、F4、F5;等級前卵泡(10 mm以下)按每1 mm為一個等級(即9~10 mm、8~9 mm、7~8 mm、6~7 mm、5~6 mm、4~5 mm、3~4 mm、2~3 mm、<2 mm);閉鎖卵泡(AF);排卵后卵泡(POF)。卵泡樣品經液氮速凍后轉移至-80℃冰箱保存,用于RNA提取。本研究中實驗動物使用程序按照四川農業大學動物護理與使用指南進行。
1.2 顆粒細胞分離培養
選擇符合1.1條件的母鵝,取出卵巢后按照本課題組的方法[9]分離并培養F1卵泡顆粒細胞,將細胞分別接種至96孔和6孔細胞培養板,分別用于細胞活性檢測和RNA提取,然后將細胞放入37℃,5%CO2培養箱中培養。
1.3 細胞處理
分別將5 mg膽固醇(Sigma)、10 mg 25-羥膽固醇(Sigma)和10 mg洛伐他汀(Sigma)粉末溶于1 mL DMSO(Omega)配置母液。待細胞培養24 h,用無血清的DMEM/F12培養基饑餓4 h,然后換為含不同濃度膽固醇(5、10、15、20μg/mL)、25-羥膽固醇(1、5、10μg/mL)和洛伐他汀(0.1、1、10μg/mL)[10-12]的無血清DMEM/F12培養基處理細胞,另設不加任何處理的空白對照組。放入37℃,5% CO2培養箱中培養24 h。
1.4 細胞活性檢測
取出96孔培養板,吸去培養基,加入25μL MTT(2 mg/mL,Omega)和75μL PBS后重新放入培養箱孵育。4 h后吸出PBS和MTT,每孔加入150μL DMSO,輕微震蕩10 min;490 nm處檢測吸光值。設不加細胞的空白對照孔,其他步驟一致,檢測時用空白對照孔校準。
1.5 RNA提取與反轉錄
取出6孔培養板,吸去培養基,PBS清洗一次。用Trizol(Invitrogen,USA)裂解細胞,按Trizol試劑標準操作提取總RNA。提取的總RNA經瓊脂糖凝膠電泳檢測質量合格后,按照PrimeScriptTMII 1st Strand cDNA Synthesis Kit(TaKaRa)說明書合成cDNA。
1.6 目的基因相對表達量檢測
采用qPCR技術檢測目的基因mRNA表達量,根據PrimeScriptTMreagent KIT Perfect Real Time試劑盒(TaKaRa)說明書操作。反應體系為25μL,其中SYBR Premix ExTaqTM(2x)12.5μL,上下游引物各0.5μL,cDNA模板2μL,Rnase Free H2O 9.5μL。PCR程序:95℃30 s;95℃0.5 s,60℃30 s,45個循環。每個樣品3個重復,相關引物參數見表1。

表1 本研究中所用引物序列Table 1 Sequences of primer used in this study
1.7 數據統計分析
利用2-△△Ct法計算基因相對表達量[13],細胞活性及基因相對表達量結果使用SPSS Statistics 22(IBM,USA)軟件進行單因素方差分析確定顯著性(P<0.05),使用Excel 2010軟件繪圖。
2 結果與分析
2.1 膽固醇轉運相關基因在鵝不同發育階段卵泡中的表達模式
膽固醇轉運與類固醇激素合成相關基因在鵝不同發育階段卵泡中的相對表達量見圖1。分析發現四個基因在各個發育階段卵泡中均有表達。隨著卵泡發育,4個基因的表達量在等級前階段先上升后下降,等級階段逐漸上升,均在排卵前(F1~F2)和直徑3~4 mm卵泡中有較高表達量,整體表達模式基本一致。除CYP11A1外,其他3個基因均在F1階段表達量最高;4個基因在POF階段的表達量均低于AF階段,其中STAR和CYP11A1在POF和AF階段的表達量顯著低于其他發育階段(P<0.05)。
2.2 不同濃度膽固醇、25-羥膽固醇和洛伐他汀對鵝顆粒細胞活性的影響
MTT法檢測不同固醇類物質處理鵝F1卵泡顆粒細胞后的細胞活性結果見圖2。隨著膽固醇處理濃度的增加,顆粒細胞活性逐漸增加,呈現劑量依賴性,在20μg/mL濃度時,細胞活性顯著增加(P<0.01);與對照組相比,添加25-羥膽固醇能在一定程度上抑制顆粒細胞活性,5μg/mL濃度時差異顯著(P=0.011);各個濃度洛伐他汀均能顯著降低顆粒細胞活性(P<0.05),但不同處理濃度之間沒有顯著差異(P>0.05)。
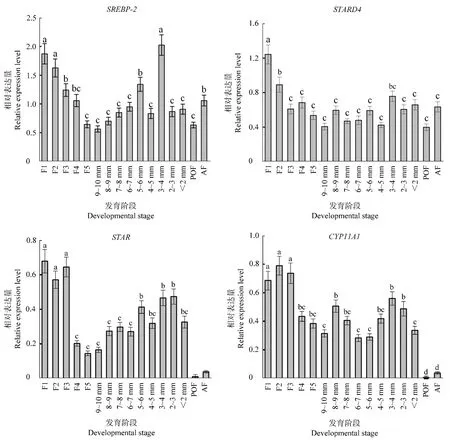
圖1 鵝不同發育階段卵泡中SREBP-2、STARD4、STAR和CYP11A1基因的表達模式Figure 1 Expression patterns of SREBP-2,STARD4,STAR and CYP11A1 genes ingeese follicles at different developmental stages
2.3 膽固醇對鵝顆粒細胞SREBP-2、STARD4、STAR和CYP11A1基因表達量的影響
不同濃度膽固醇處理鵝F1卵泡顆粒細胞后SREBP-2、STARD4、STAR和CYP11A1基因的表達量見圖3。隨著膽固醇處理濃度的增加,4個基因的表達量均表現出先上升后下降的變化趨勢,20μg/mL濃度時表達量最低。SREBP-2在15μg/mL濃度時表達量與10μg/mL濃度時相比顯著下降(P<0.001),但其他3個基因在這兩個處理濃度間沒有顯著差異(P>0.05)。
2.4 25-羥膽固醇對鵝顆粒細胞SREBP-2、STARD4、STAR和CYP11A1基因表達量的影響
不同濃度25-羥膽固醇處理鵝F1卵泡顆粒細胞后SREBP-2、STARD4、STAR和CYP11A1基因的表達量見圖4。4個基因mRNA水平在25-羥膽固醇影響下有不同的變化趨勢。隨著處理濃度的增加,SREBP-2表達量逐漸降低,且差異顯著(P<0.05),表現出劑量依賴性;低濃度25-羥膽固醇對STARD4表達量沒有影響(P>0.05),而10μg/mL時STARD4表達量顯著升高(P<0.01);添加25-羥膽固醇能顯著降低STAR表達量(P<0.001),但不同濃度之間沒有顯著差異(P>0.05);與對照組相比,低濃度25-羥膽固醇處理能顯著降低CYP11A1的表達量(P<0.05),但濃度增加至10μg/mL時,CYP11A1表達量升高,且與對照組沒有顯著差異(P>0.05)。
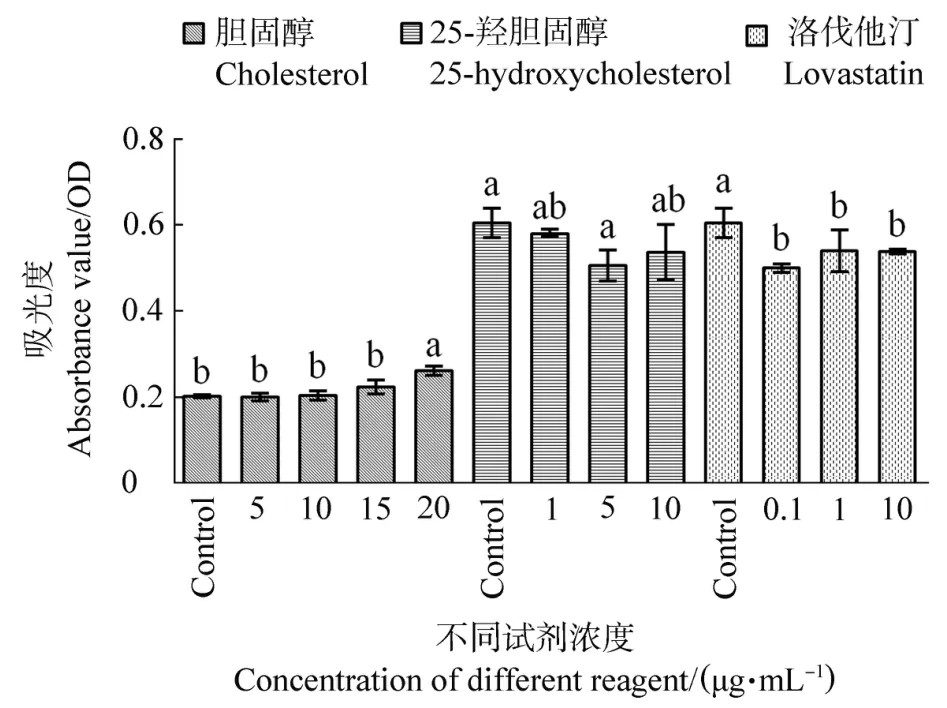
圖2 不同濃度膽固醇、25-羥膽固醇和洛伐他汀處理后鵝F1卵泡顆粒細胞活性Figure 2 Goose F1 granulosa cells activity treated with different concentration scholes terol,25-hydroxycholesterol or lovastatin
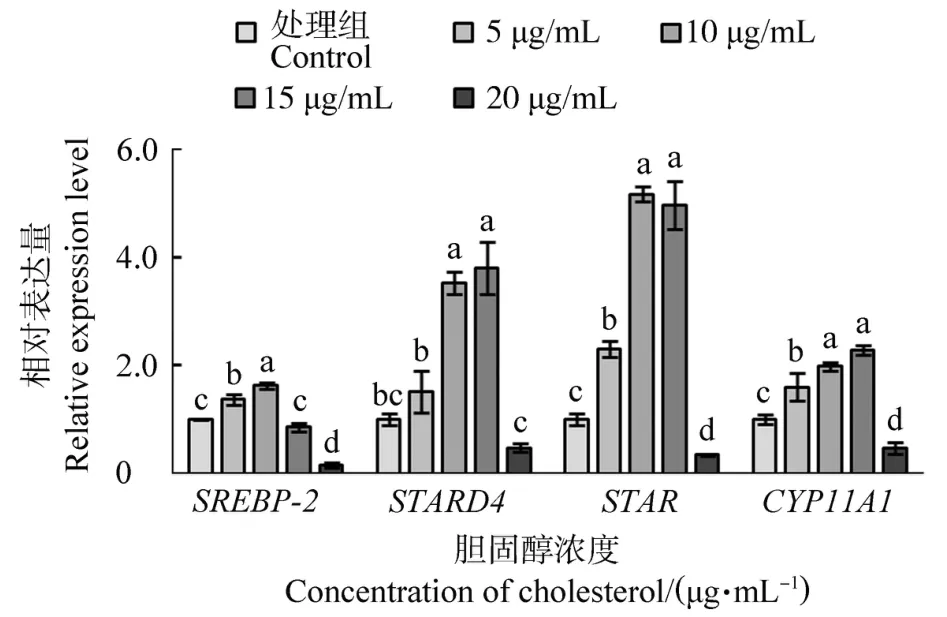
圖3 不同濃度膽固醇處理鵝F1卵泡顆粒細胞后SREBP-2、STARD4、STAR和CYP11A1基因的表達量Figure 3 The expression of SREBP-2,STADR4,STAR and CYP11A1 in goose F1 granulosa cells treated with different concentrations of cholesterol
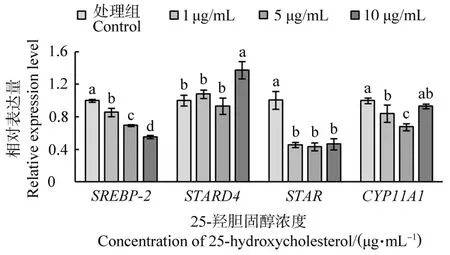
圖4 不同濃度25-羥膽固醇處理鵝F1卵泡顆粒細胞后SREBP-2、STARD4、STAR和CYP11A1基因的表達量Figure 4 The expression of SREBP-2,STADR4,STAR and CYP11A1 in goose F1 granulosa cells treated with different concentrations of 25-hydroxycholesterol
2.5 洛伐他汀對鵝顆粒細胞SREBP-2、STARD4、STAR和CYP11A1基因表達量的影響
不同濃度洛伐他汀處理鵝F1卵泡顆粒細胞后SREBP-2、STARD4、STAR和CYP11A1基因的表達量見圖5。隨著處理濃度升高,SREBP-2表達量先降低后升高,而其余3個基因均表現出先升高后降低的趨勢,與SREBP-2相反。與其余兩個處理濃度相比,1μg/mL洛伐他汀處理細胞后,SREBP-2表達量顯著下降(P<0.05),而STARD4、STAR和CYP11A1表達量顯著升高(P<0.05)。
3 討論
本實驗檢測了SREBP-2、STARD4、STAR和CYP11A1基因在鵝不同發育階段卵泡中的表達情況,分析發現4個基因表達趨勢相似,均在等級階段表達量逐漸上升,到F1~F2時達到頂峰。在禽類等級卵泡中,促性腺激素主要刺激顆粒細胞分泌孕酮以促進排卵。研究表明,雞等級卵泡中顆粒細胞合成孕酮的能力隨著卵泡逐漸成熟而增強,發育到F1卵泡時達到頂峰[14],在鵝上也有相似的報道[15]。STARD4是SREBP-2的靶基因[7],實驗結果發現STARD4和SREBP-2在鵝卵泡的不同發育階段表達模式基本一致,并和卵泡類固醇激素合成規律相同,表明二者在類固醇激素合成過程中可能通過參與膽固醇轉運發揮作用。接近排卵階段的卵泡顆粒細胞中促黃體生成素受體(LHR)表達量增加,因此LH介導的孕酮生成增加,這個過程就伴隨著STAR表達量增加[16-17],另外此時CYP11A1主要在顆粒細胞表達,隨后可以誘導LH峰出現和排卵[18-20]。綜上,本研究后續實驗選擇鵝F1階段卵泡的顆粒細胞為實驗材料,初步探討不同固醇類物質對上述4個基因的影響。
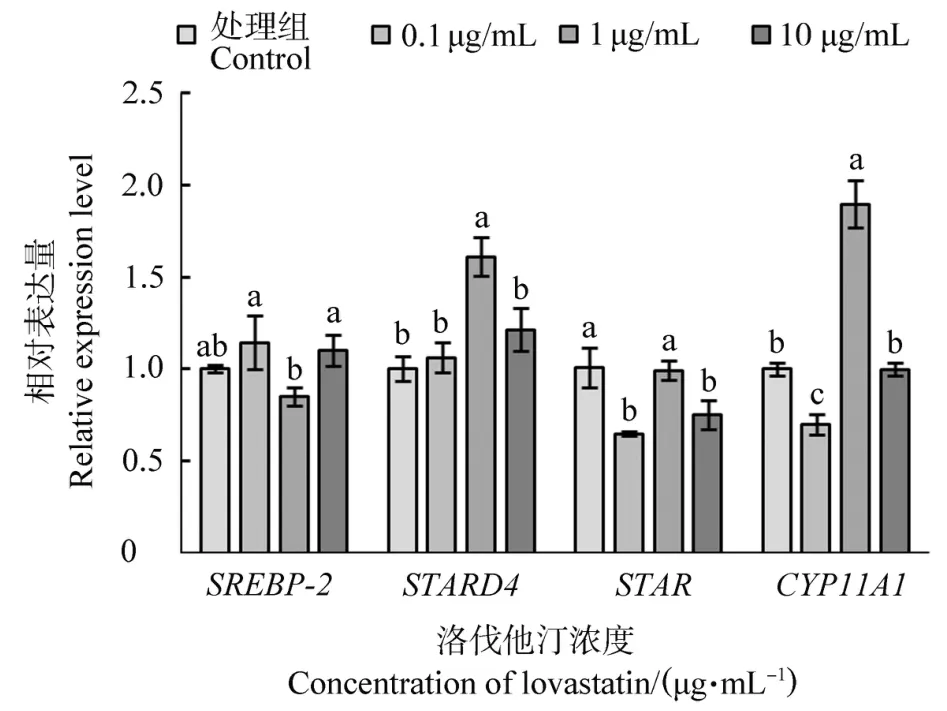
圖5 不同濃度洛伐他汀處理鵝F1卵泡顆粒細胞后SREBP-2、STARD4、STAR和CYP11A1基因的表達量Figure 5 The expression of SREBP-2,STARD4,STAR and CYP11A1 in goose F1 granulosa cells treated with different concentrations of lovastatin
細胞實驗結果顯示,不同濃度膽固醇處理顆粒細胞后,4個基因表達量隨著膽固醇濃度增加都表現出先上升后下降的趨勢,且SREBP-2在15μg/mL時表達量已顯著下降,其他3個基因則沒有。在飼喂高膽固醇飲食的小鼠肝臟中檢測到STADR4表達量降低超過2倍,并與SREBP-2表現出協調調節[21]。Ning Y.[22]等發現,膽固醇能增加小鼠腦微血管內皮細胞中STAR表達量,并存在劑量效應。推測當細胞內膽固醇濃度升高時,STADR4表達量增加,促進細胞內膽固醇代謝平衡;同時STAR從線粒體外膜轉運進內膜的膽固醇增加,引起CYP11A1表達量上升。而當膽固醇濃度過高時,抑制SREBP-2和STADR4表達,進一步降低STAR和CYP11A1表達量,此時可能通過刺激細胞內膽固醇代謝增加了細胞活性。此外,SREBP-2對膽固醇濃度可能更敏感,這有助于它及時通過調節STADR4表達確保內質網對細胞器(包括質膜)膽固醇豐度的敏感性,從而保證細胞內膽固醇水平的穩定[8]。25-羥膽固醇是膽固醇氧化產物,它可以通過抑制SREBP-2裂解來抑制其下游基因轉錄激活,最終降低細胞內膽固醇合成[23]。25-羥膽固醇還可以激活膽固醇酯化,此時細胞內膽固醇轉運蛋白STADR4的表達量會有所上升,這與本實驗結果相符。羥甲基戊二酸單酰輔酶A還原酶(HMGCR)是合成膽固醇的限速酶,洛伐他汀是一種HMGCR競爭性抑制劑,可以抑制細胞內膽固醇合成[24]。洛伐他汀處理小鼠巨噬細胞源性泡沫細胞,可明顯減少細胞內總膽固醇和膽固醇酯含量[25]。實驗結果表明,不同濃度洛伐他汀處理后STARD4、STAR和CYP11A1表達趨勢相似,推測洛伐他汀通過影響胞內STARD4來調節膽固醇平衡,從而影響STAR和CYP11A1表達量。25-羥膽固醇和洛伐他汀都能降低細胞內膽固醇含量,但其作用方式不同,可能因此對相關基因的影響也不同。另外,二者都可以誘導細胞凋亡[26-27],從本實驗結果來看,洛伐他汀對顆粒細胞活性的抑制效應強于25-羥膽固醇。
4 結論
SREBP-2、STARD4、STAR和CYP11A1基因在鵝產蛋期不同發育階段卵泡中表達模式基本一致;高濃度膽固醇能增加鵝F1卵泡顆粒細胞活性,25-羥膽固醇和洛伐他汀與之相反;不同固醇類物質通過不同方式影響細胞內膽固醇水平,從而可能對膽固醇轉運和類固醇合成相關基因產生不同影響。本實驗結果可以為進一步揭示鵝卵泡發育機制和提高產蛋量提供理論參考。
