苦蕎轉錄因子基因FtMYC的克隆及其表達與花青素積累的相關性分析
2019-03-11 05:23:26姚攀鋒呂兵兵董玘鑫王安虎
四川農業大學學報 2019年1期
姚攀鋒,呂兵兵,李 琪,董玘鑫,王安虎,吳 琦*
(1.四川農業大學生命科學學院,四川 雅安625014;2.西昌學院,四川 西昌615000)
苦蕎(Fagopyrum tataricum)屬于蓼科蕎麥屬一年生草本植物,由于其富含蘆丁、花青素等黃酮類化合物,因此,對人體具有降“三高”等多種保健功能[1]。與此同時,黃酮類化合物也有助于提高植物對逆境的耐受力,植物會通過調控黃酮化合物合成相關基因的表達對環境刺激迫作出響應,由此形成一種自我保護機制[2]。研究表明,苦蕎黃酮中的花青素含量上升常常伴隨著逆境的出現,參與抵抗紫外輻射、抗寒和抗旱等多個生理過程。Li S.等[3]對芽期苦蕎子葉和胚軸分別進行了低溫脅迫后發現,處理后苦蕎子葉和胚軸中花青素含量均顯著提高。此外,苦蕎冷處理后的轉錄組和代謝組分析顯示,4℃脅迫之后苦蕎的矢車菊素3-0-葡萄糖苷和矢車菊素3-0-蕓香糖苷相比正常條件下分別提高了6.3和11.3倍[4]。因此,花青素作為苦蕎體內一類重要的黃酮類化合物,在苦蕎的抗逆機制中起著重要的作用。
花青素生物合成途徑是類黃酮物質合成途徑的一個分支,參與其生物合成相關的基因主要包括結構基因和轉錄因子這兩大類,二者共同調控植物花青素的代謝合成[5]。參與花青素支路調控的轉錄因子眾多,主要包括MYB、bHLH和WD40等,它們單獨或者形成復合物發揮生物學作用[5]。其中,bHLH轉錄因子是植物中僅次于MYB轉錄因子的第二大轉錄因子超家族廣泛存在于植物中,在植物的生長發育與形態建成、次級代謝產物的合成及對外界環境脅迫應答中起著重要的調控作用[5-6]。例如,在番茄中持續表達ZmLc、Delila和MYC-RP/GP等bHLH類蛋白,能使花青素在地上部分(包括果實)和根中大量積累[7]。過表達Delila(bHLH)基因,在果實中特異的啟動子E8的控制下,能導致花青素在果肉和果皮中強烈的增加,果實呈暗紫色[5]。可見,bHLH轉基因能激活花青素代謝途徑中一系列結構基因,促進花青素等黃酮物質在植物中的大量積累,較單個結構基因的轉基因具有更為廣泛的作用。
本實驗根據獲得的苦蕎花期轉錄組數據,采用RT-PCR技術克隆出一條與花青素調控相關的bHLH轉錄因子MYC亞家族的基因FtMYC,并采用UV-B、4℃冷脅迫處理二葉期苦蕎,探究FtMYC基因在非生物脅迫條件下的表達量變化與花青素含量的相關性,并分析了FtMYC基因啟動子上的主要順式作用元件,初步明晰FtMYC的結構及其在苦蕎逆境脅迫中的作用。
1 材料和方法
1.1 材料及主要試劑
苦蕎“西蕎2號”由西昌學院王安虎教授惠贈,栽培在四川農業大學生命科學學院人工氣候室;植物RNAout(Trizol法)試劑盒、TaqDNA聚合酶、PrimeSTAR Max DNA Polymerase、SYBR Premix ExTaqTMII(Perfect Real Time)購自TaKaRa公司,其他試劑為國產分析純。
1.2 試驗方法
1.2.1 苦蕎FtMYC基因的克隆
采用SDS法提取苦蕎葉片總DNA,采用植物RNAout試劑盒參照說明書提取苦蕎總RNA,并反轉錄制備cDNA第一條鏈。根據本實驗室的苦蕎花期轉錄組數據,設計一對特異引物分別以苦蕎總DNA和cDNA第一鏈為模板擴增FtMYCDNA和cDNA序列,引物序列:FtMYCf(5'-ATGCAGGCAAATCTC AGAGAAC-3')和FtMYCr(5'-TCACCCACATTTCG TTATAGCT-3')。將PCR產物亞克隆至T載體,送擎科生物技術有限公司測序。
1.2.2 苦蕎FtMYC基因的生物信息學分析
通過NCBI在線工具Blast對FtMYC氨基酸序列進行同源比對;通過DNAMAN軟件FtMYC及其它同源蛋白進行多重序列比對,分析它的保守結構域;通過MEGA 5.0軟件,根據鄰接法(neighbor-joining,NJ)構建FtMYC的系統發育進化樹。
1.2.3 二葉期苦蕎的脅迫處理
UV-B處理:使用10W UV-B(308 nm)LED燈照射二葉期苦蕎,光照距離30 cm;4℃冷處理:在光照條件為16 h/8 h(光照/黑暗)、4℃的培養箱對子葉期的苦蕎進行冷處理。以上處理均在處理前0 h、處理后1、3、6、12、24和48 h分別取樣品的胚軸和子葉液氮速凍后放在-80℃超低溫冰箱保存。
1.2.4 苦蕎FtMYC基因表達量的檢測
根據FtMYC基因序列,設計一對熒光定量PCR引物:qFtMYCf(5'-AAT GCG AAT CAG GCG GAC A-3')和qFtMYCr(5'-GGC ACC CAA CTC GACA ACA C-3')。以苦蕎FtH3基因為內參基因,其引物為:qFtH3f(5'-GAA ATT CGC AAG TAC CAG AAG AG-3')和qFtH3r(5'-CCA ACA AGG TAT GCC TCA GC-3')。以cDNA為模板,采用SYBR Premix ExTaqTMII(Perfect Real Time)試劑盒檢測FtMYC基因在不同條件處理下的表達水平,通過2-ΔΔCt方法來計算它的相對表達量。
1.2.5 苦蕎花青素的提取及定量測定
準確稱取0.2 g新鮮樣品用液氮研磨后加入1 mL 1%鹽酸的甲醇提取液在25℃、100 r/min條件下震蕩浸提18 h;3 080 r/min離心15 min后取500μL上清,并加入等體積的雙蒸水和300μL氯仿混勻;2 268 r/min離心5 min后取上清,即為花青素提取液。最后通過紫外分光光度計檢測530 nm和657 nm處的吸光值。采用如下公式計算花青素的量:花青素(mg/g)=(A530-0.25×A657)/鮮質量。
1.2.6 苦蕎FtMYC基因啟動子克隆及的生物信息學分析
根據苦蕎基因組數據(http://gsa.big.ac.cn/index.jsp),設計一對特異引物PFtMYCf(5'-CGG GAA TAT AAT GAT TCG AGA CAA-3')和PFtMYCr(5'-CAT TAT GCT TTG ACA AGC AAG C-3'),以苦蕎DNA為模板擴增FtMYC基因啟動子序列,PCR產物亞克隆至T載體后,送擎科生物技術有限公司測序。通過Plant CARE數據庫(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)和PLACE(http://www.dna.affrc.go.jp/PLACE/)數據庫對基因5’側翼序列所含元件進行分析預測,利用啟動子核心分析數據庫BDGP(http://www.fruitfly.org/seq-tools/promoter.html)對FtMYC基因啟動子的核心元件進行預測。
1.3 數據的分析和處理
采用IBM SPASS Statistics 20.0統計軟件對苦蕎FtMYC基因的表達量與各樣本花青素的量進行相關性分析。
2 結果與分析
2.1 FtMYC基因cDNA序列的克隆
分別以苦蕎cDNA和總DNA為模版擴增得到2條大小約為1 300 bp的特異條帶,見圖1。測序結果顯示該基因的cDNA和DNA片段分別為1 287和1 309 bp。根據已獲得的苦蕎FtMYC基因的cDNA和DNA序列,利用DNAMAN軟件比對確定其全長DNA序列為1 309 bp,有1個外顯子和1個內含子構成(見圖2),符合標準的GT-AG剪切原則。Blastp比對結果表明,苦蕎FtMYC基因屬于bHLH-MYC亞家族。選取bHLH家族中擬南芥AtGL3、葡萄VvMYCA1、紫蘇PfMYC-RP和金魚草AmDELILA進行多重序列比對,發現苦蕎FtMYC在N-端包含一個由191個氨基酸組成的bHLH-MYC-N結構域,該結構域屬于bHLH轉錄因子家族中MYC蛋白的特殊結構域[8],見圖3。在C-端的bHLH結構域處也具有較高的保守性。以上結果表明,本文獲得的FtMYC基因屬于bHLH轉錄因子中的MYC亞家族,其在GenBank中的登錄號為:KU162971。
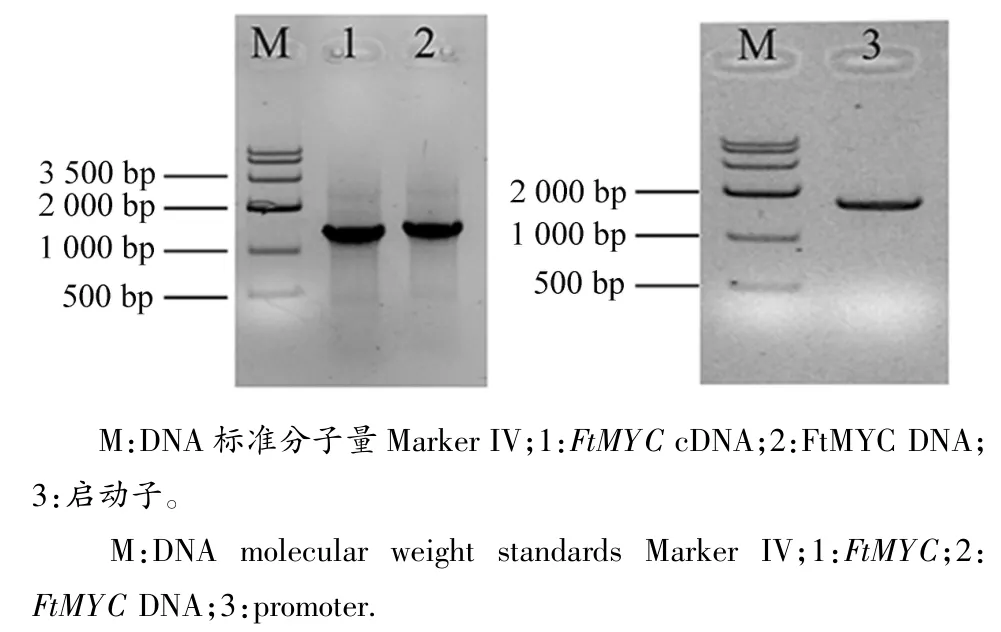
圖1 苦蕎FtMYC基因cDNA和啟動子的擴增Figure 1 Amplification of FtMYC and promoter from tartary buckwheat

圖2 苦蕎FtMYC基因結構示意圖Figure 2 Structure diagram of FtMYC

圖3 不同植物bHLH氨基酸序列比對Figure 3 Alignment of amino acid sequences of bHLH from different plants
2.2 苦蕎FtMYC系統進化分析
采用MEGA 5.0鄰接法構建基于FtMYC氨基酸序列的系統發育進化樹(見圖4)。選擇均已有功能相關性研究的植物MYC蛋白序列,結合M.Heim等[9]根據bHLH保守域氨基酸組成特點對擬南芥133個bHLH蛋白的結構解析及分類,初步分析苦蕎該FtMYC可能具有的功能。圖4中的進化樹分析表明,這些MYC蛋白可明顯分為4個大簇,由于這4簇之間氨基酸序列間差異較大,推測其具有不同的進化模式,表現出功能分化。苦蕎(Fagopyrum tataricum)FtMYC與擬南芥(Arabidopsis thaliana)AtGL3、葡萄(Vitis vinifera)VvMYCA1、紫蘇(Perilla frutescens)PfMYC-RP和金魚草(Antirrhinum majus)AmDELILA歸于一簇,而這些蛋白均參與花青素代謝的有關調控,推測苦蕎FtMYC具有相似的功能。
2.3 非生物逆境脅迫下苦蕎FtMYC的表達與花青素含量分析
2.3.1 UV-B對苦蕎FtMYC表達和花青素含量的影響
使用熒光定量PCR分析FtMYC在UV-B處理下芽期苦蕎的轉錄水平,結果表明:在胚軸中,FtMYC的表達量從處理1 h開始上升,每隔3 h呈現明顯的上升趨勢,處理2 h后顯著上升(P<0.05),為處理前(0 h)的1.83倍(見圖5);子葉中,FtMYC在的表達量變化則更為明顯,處理6 h后極顯著上升(P<0.01),為對照組的13.29倍,隨后下降,48 h后接近處理前水平(見圖6)。同時,花青素含量分析結果表明:胚軸中的花青素含量隨著處理時間不斷升高,12 h后為對照組的1.88倍并趨于穩定(見圖5);子葉中花青素含量的變化更為明顯,處理6 h后顯著性提高(P<0.05),對照組的1.44倍(見圖6)。
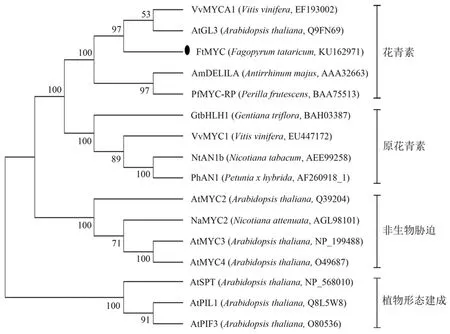
圖4 苦蕎FtMYC與其他植物來源MYC蛋白序列的系統進化樹Figure 4 Phylogenetic tree based on amino acids equences of FtMYC and other plants

圖5 UV-B對苦蕎胚軸FtMYC表達量和花青素含量的影響Figure 5 Expression levels FtMYC gene and anthocyanins content in hypocotyl under UV-B treatment condition
2.3.2 冷脅迫對苦蕎FtMYC表達和花青素含量的影響
使用熒光定量PCR分析FtMYC在冷處理下芽期苦蕎的轉錄水平,結果表明:FtMYC在胚軸中的表達量從處理1 h開始上升,但上升趨勢不明顯,12 h后達到最大為對照組的1.67倍,之后下降接近對照水平(見圖7);而FtMYC在子葉中的表達量變化卻很明顯,1 h后就發生極顯著性變化(P<0.01),12 h后達到對照組的15.3倍,之后降低為對照組的1.83倍(見圖8)。花青素含量測定結果顯示:在該條件下苦蕎胚軸、子葉中花青素含量的變化趨勢基本一致,均在12 h后極顯著提高(P<0.01),分別為對照組的2.29倍和1.94倍。

圖6 UV-B對苦蕎子葉FtMYC表達量和花青素含量的影響Figure 6 Expression levels FtMYC gene and anthocyanins content in cotyledon under UV-B treatment condition
2.3.3 苦蕎FtMYC基因表達與花青素含量的相關性分析
使用IBM SPSS Statistics 20統計軟件分析不同脅迫條件下苦蕎胚軸和子葉中FtMYC基因的相對表達量與花青素含量的相關性(見表1)。以相關系數(r)絕對值大于0.75位閾值時,UV-B處理下FtMYC表達量與苦蕎胚軸中花青素含量變化顯著性相關(r=0.798,P<0.05),與子葉中花青素含量變化極顯著相關(r=0.887,P<0.01)。4℃冷處理下FtMYC表達量與苦蕎胚軸中花青素含量變化沒有明顯的相關性(r=0.744),但與子葉中花青素含量變化顯著性相關(r=0.814,P<0.05)。結果表明,苦蕎FtMYC基因表達量與花青素含量具有較強的正相關。

圖7 冷脅迫對苦蕎胚軸FtMYC表達量和花青素含量的影響Figure 7 Expression levels FtMYC gene and anthocyanins content in hypocotyl under cold treatment condition
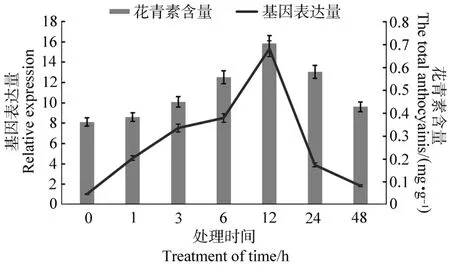
圖8 冷脅迫對苦蕎子葉FtMYC表達量和花青素含量的影響Figure 8 Expression levels FtMYC gene and anthocyanins content in cotyledon under cold treatment condition

表1 苦蕎子葉和胚軸中花青素含量與FtMYC基因表達量的相關性Table 1 Correlation coefficients between FtMYC expression levels and total anthocyanins contents in cotyledons and hypocotyl of Tartary buckwheat
2.4 FtMYC基因啟動子元件分析
以苦蕎DNA為模版,PCR擴增得到一條1684bp的特異條帶,見圖1。測序結果與苦蕎基因組數據完全一致,表明獲得了FtMYC基因的啟動子序列,將其命名為pFtMYC。本研究進一步采用DNAMAN軟件對pFtMYC的序列進行了生物信息學分析,結果顯示:pFtMYC序列包含了63.7%的A/T堿基,符合植物啟動子的基本特征。通過在線網站對該啟動子序列的分析結果顯示,pFtMYC中包含有大量的順式作用元件,比如啟動子核心元件、環境和激素類響應元件等(見表2)。其中,與環境響應相關的元件所占比例較大,例如低溫應答元件、光響應元件、真菌誘導調節元件、熱脅迫響應元件等等。此外,pFtMYC中還存在一些其他類型的元件,如激素應答元件、蛋白結合位點以及分生組織表達相關應答元件。以上表明,多種環境因子均可調控苦蕎FtMYC基因的表達。
3 討論與結論
MYC蛋白屬于bHLH家族的轉錄因子,該類轉錄因子在動植物體內具有多種生物學功能。MYC蛋白的N-端含有一個bHLH-MYC-N結構域,C-端含有一個結合目的基因啟動子中E-box的bHLH型DNA結合域,MYC蛋白主要通過結合靶標基因啟動子序列中E-box元件的保守序列CAC(G/A)TG來調節基因的表達[8]。本研究利用苦蕎轉錄組數據成功克隆得到1個bHLH家族的轉錄因子基因FtMYC,因其編碼的氨基酸序列N-端和C-端區域分別包含有MYC轉錄因子典型的保守結構域,這表明FtMYC屬于bHLH轉錄因子中MYC家族的一個成員。
植物bHLH轉錄因子能夠參與調控多種生理途徑,如花器官發育、光形態建成、植物抗逆等,其中調節類黃酮和花青素合成是植物bHLH轉錄因子最重要功能之一[10]。系統進化樹分析顯示(見圖4),FtMYC和VvMYCA1等參與花青素調控的轉錄因子共聚一簇,推測FtMYC可能具有類似的調控功能。在UV-B和4℃冷脅迫處理條件下,FtMYC在苦蕎二葉期胚軸和子葉中的表達量均表現出一定程度上調趨勢,相比之下在子葉中變化趨勢更為顯著,這種結果表明FtMYC在苦蕎二葉期更傾向于在子葉中發揮調控作用。與此同時,當FtMYC在兩個組織中的表達量達到最大值后在都出現了不同程度下降的趨勢,可能是因為植物本身存在自我修復機制,在遭到外界脅迫一定時間后通過該機制來維持機體的代謝平衡[11]。眾所周知,花青素對UV-B波段存在特定吸收波長,能有效降低UV-B輻射在表皮層的透過率,從而減輕UV-B對植物器官與組織的傷害[12]。而當植物處于極端溫度逆境下,主要通過影響細胞的膜結構從而產生氧化脅迫[13],由于花青素本身具有較強的抗氧化作用,因此花青素在植物組織中的積累可以增強其對于極端環境的耐受性[14]。例如,蘋果MdbHLH3轉錄因子能在低溫條件下會特異性結合MdDFR和MdUFGT基因啟動子序列,通過上調花青素支路的關鍵基因的表達促進蘋果中花青素的積累[15]。本研究中除了4℃冷處理下FtMYC表達量與苦蕎胚軸中花青素含量變化沒有明顯的相關性(r=0.744)外,其他條件下均顯著性相關。由此可知,FtMYC也可能通過類似的調控機制實現了苦蕎體內花青素的積累,以此來應對外界的非生物逆境脅迫。
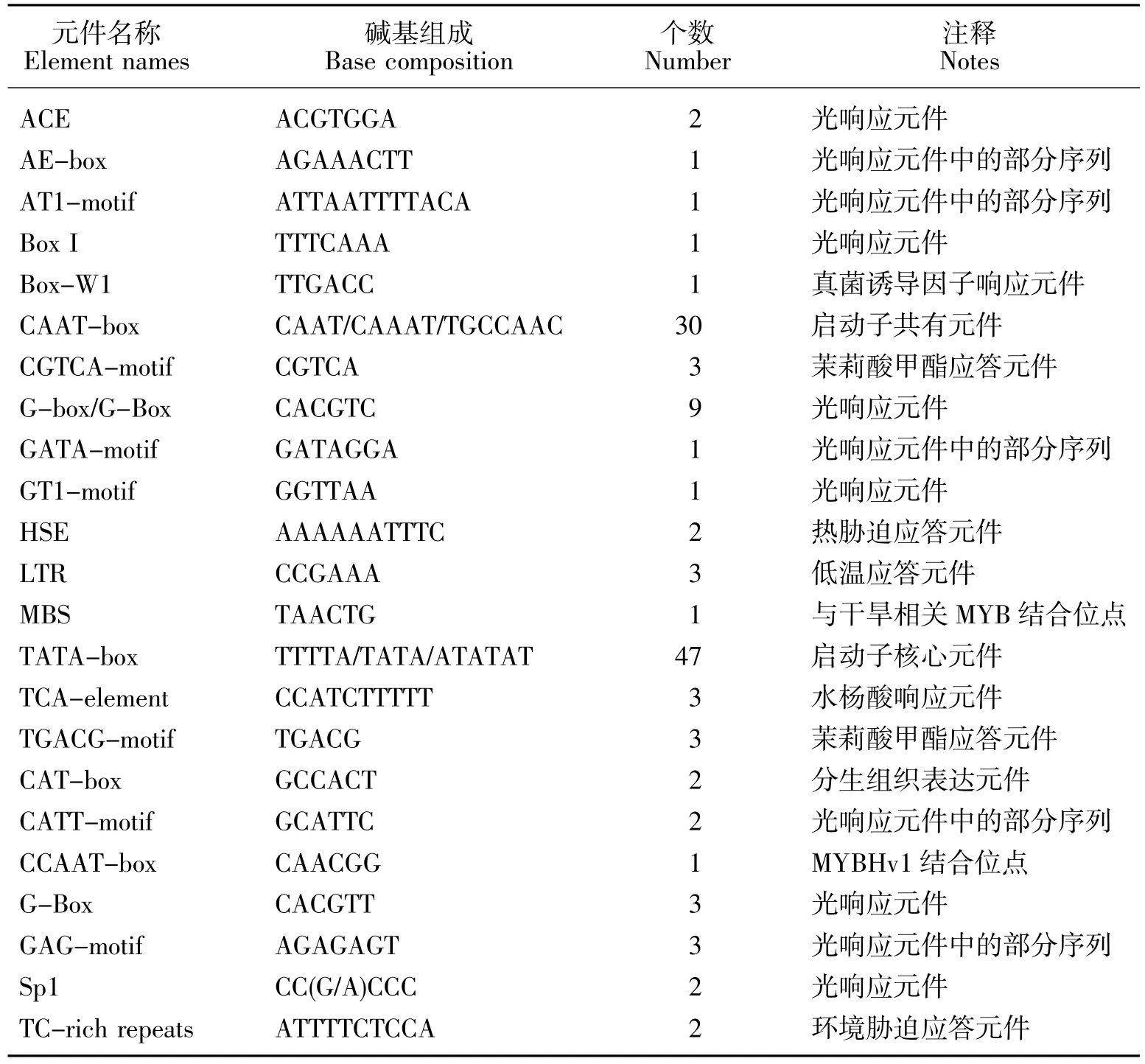
表2 pFtMYC所含主要元件分析Table 2 Component analysis of pFtMYC
轉錄水平的調控是植物基因表達調控的一種重要方式,而啟動子作為執行轉錄水平調控的主要順式作用元件,能應答多種環境的變化[16]。轉錄因子對環境刺激的響應主要是通過其啟動子上各類響應元件實現的[17],Yao P.F.等[18]對pFtbHLH3研究結果顯示,干旱脅迫處理會提高pFtbHLH3下游報告基因的表達量。本研究在對FtMYC啟動子的分析中發現,pFtMYC中除了含有基本的啟動子轉錄元件外,還包含有多個與光、低溫脅迫應答相關的元件,我們推測FtMYC的表達可能通過這些元件響應外界環境的刺激。本課題組后續也將對FtMYC的功能展開進一步研究,以期明晰FtMYC轉錄因子在苦蕎花青素合成的調控機制,為提高苦蕎黃酮含量的研究奠定一定的理論基礎。
猜你喜歡
農產品加工(2023年14期)2023-08-17 10:32:28
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
青年歌聲(2018年5期)2018-10-29 03:18:40
山東工業技術(2016年15期)2016-12-01 05:31:22
廣東飼料(2016年3期)2016-12-01 03:43:12
家庭百事通·健康一點通(2016年4期)2016-04-22 09:16:57
林業與生態(2016年2期)2016-02-27 14:24:01
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
