弓形蟲沉默信號調節子2(TgSir2)的克隆表達與多克隆抗體制備
2019-03-07 08:40:30張芳菲曹佳欣王晨紅毛懿杰華倩倩劉淑賢
中國人獸共患病學報 2019年2期
關鍵詞:血清
張芳菲, 曹佳欣, 王晨紅, 毛懿杰, 華倩倩, 劉淑賢, 譚 峰, 胡 昕
剛地弓形蟲廣泛寄生于人和動物的有核細胞內,引起弓形蟲病[1]。據統計,全世界約有1/3人群屬于弓形蟲隱性感染人群[2]。當機體免疫力受損或抑制,緩殖子期可轉化為速殖子期,在患者體內播散,造成多器官炎癥反應,嚴重可致死亡[3-4]。然而目前對弓形蟲病的治療主要針對的是顯性感染階段速殖子,無法根除隱性感染人群體內的緩殖子,阻礙了疾病的治療[5]。
沉默信號調節子2(Silent information regulator 2, Sir2),是一類高度保守的、煙酰胺腺嘌呤二核苷酸(NAD+)依賴的組蛋白去乙酰化酶(histone deacetylase, HDAC),屬第III類HDAC家族——sirtuins家族[6-7],其底物包括組蛋白、非組蛋白等多種蛋白[7]。
Sir2最早發現于酵母細胞中,隨后在多種原核、真核細胞中被證實。人體內,Sir2被命名為SIRT2。SIRT2既可定位胞漿,也可入核,對胞漿蛋白、組蛋白或轉錄因子進行去乙酰化修飾,參與微管動力學、氧化應激反應、細胞周期調控等許多生理和細胞活動,與腫瘤、衰老、II型糖尿病等疾病密切相關[7]。
然而,在弓形蟲體內尚未見TgSir2蛋白的相關研究報道。據此,本研究利用生物信息學分析方法,在弓形蟲基因組中挖掘出與酵母細胞TgSir2蛋白的同源蛋白,通過原核表達TgSir2,制備其多克隆抗體,為進一步研究該蛋白功能提供實驗材料。
1 材料與方法
1.1 材 料
1.1.1蟲株、質粒、菌株及實驗動物 弓形蟲RH株速殖子、人包皮成纖維細胞(HFF)、原核表達載體pET28b、大腸埃希菌(E.coli)DH5α和BL21均為本實驗室保存,新西蘭大白兔購自溫州醫科大學實驗動物中心。
1.1.2逆轉錄PCR(RT-PCR)引物的設計與合成 根據ToxoDB中TgSir2基因編碼序列(TGGT1_227020),分別設計去除信號肽片段的上游引物(TgSir2△1-15aa-F: 5′GAATTCTTGCCTCTTTGAGCCTCATGAAC-3′)和下游引物(TgSir2△1-15aa-R: 5′-GCGGCCGCCTATCTCCTTTGCAGCGCC-TT-3′),下劃線部分分別為限制性內切酶EcoRI和NotI的酶切位點。引物由上海祥音生物科技有限公司合成。
1.1.3主要試劑 Trizol Reagent試劑盒、4×蛋白上樣緩沖液購自美國Invitrogen 公司;逆轉錄試劑盒RNA PCR Kit(AMV)Ver3.0購自日本Takara公司;KAPA HiFi HotStart ReadyMix PCR Kit購自美國KAPA Biosystems公司;DNA凝膠回收試劑盒和質粒提取試劑盒購自美國OMEGA Bio-tek公司;限制性內切酶NotI、EcoRI和T4 DNA連接酶購美國Thermo Fisher Scientific公司;GoTaqTMGreen Master Mix購自美國Promega公司;異丙基-β-D-硫代半乳糖苷(IPTG)購自美國Sigma公司;蛋白酶抑制劑購自瑞士Roche 生物公司;蛋白純化鎳柱Ni-NTA Agarose購自QIAGEN公司;鼠源抗His單克隆抗體購自美國CMCTAG公司;Dlight 488山羊抗兔IgG抗體購自美國EarthOx life science公司;辣根過氧化物酶標記的山羊抗小鼠IgG抗體、山羊抗兔 IgG 抗體購自合肥市Biosharp生物科技公司,0.22 μm PVDF轉移膜購自美國Millipore公司。
1.2 方 法
1.2.1生物學信息分析 在弓形蟲數據庫ToxoDB中,采用Blast序列比對軟件進行氨基酸序列比對,尋找與酵母Sir2相似性最高的蛋白序列,并利用ClustalW軟件將TgSir2與其他物種Sir2同源物進行同源比對。
1.2.2目的片段擴增將RH株速殖子在HFF宿主細胞中常規培養(1%胎牛血清的DMEM)。待蟲體完全出胞后,將蟲體經3 μm濾器純化后以Trizol法提取總RNA并逆轉錄合成cDNA,以cDNA為模板,擴增TgSir2△1-15aa編碼序列(去除1-15位氨基酸的編碼序列)反應體系為:cDNA(<1 ng),上、下游引物(10 nmol)各1.5 μL,2×Kapa HiFi hotstart ready mix 25 μL,加去離子水至50 μL。反應條件為:95 ℃預變性1 min,98 ℃變性30 s、60 ℃退火30 s、72 ℃延伸1 min,共30個循環,72 ℃再延伸10 min。
1.2.3重組質粒pET28b-TgSir2△1-15aa的構建與鑒定 用NotI和EcoRI酶分別雙酶切pET-28b載體、PCR目的片段。膠回收酶切產物,連接后將連接產物轉化入DH5α感受態細胞,涂Kan LB固體培養基平板篩選陽性單克隆,提質粒通過PCR擴增和雙酶切鑒定后,陽性質粒送北京六合華大基因科技有限公司測序驗證。
1.2.4TgSir2蛋白誘導表達、純化與鑒定 將測序正確的重組質粒轉化入BL21感受態細胞,挑取單克隆菌落,經IPTG于16 ℃ 220 r/min振蕩誘導22 h。將誘導后產物離心收集菌體,超聲裂解后4 ℃ 12 000 r/min離心10 min,鎳柱純化上清中的TgSir2融合蛋白,隨后以小鼠抗His單克隆抗體作為一抗(1∶1 000),羊抗小鼠HRP-IgG作為二抗(1∶5 000),進行12%十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE)與Western blotting分析。
1.2.5抗體制備 將純化后的TgSir2融合蛋白背部皮下多點注射免疫2只新西蘭大白兔,初次免疫以1 mg,之后每隔15 d,0.5 mg抗原加強免疫1次,共4次。
1.2.6抗體效價和特異性鑒定 免疫結束后,采集兔血清,以His-TgSir2△1-15a蛋白包被酶標板,間接酶聯免疫吸附試驗(ELISA)法檢測免疫后兔血清抗體效價。待血清效價達到1∶64 000后常規心臟取血,收集兩只兔血清,分別命名為1號和2號免疫兔抗TgSir2血清,于-80 ℃保存備用。同時將純化的His-TgSir2△1-15a蛋白經SDS-PAGE分離,轉膜、封閉,后依次用經親和層析純化的1號和2號免疫兔抗TgSir2多克隆抗體作為一抗,羊抗兔HRP-IgG作為二抗,電化學發光試劑(ECL)顯色,觀察結果。
1.2.7多抗識別弓形蟲內源性TgSir2將從弓形蟲中提取的蛋白(按步驟1.2.2收集弓形蟲RH株速殖子,超聲裂解蟲體后,取上清)行SDS-PAGE后,以制備的兔抗TgSir2多克隆抗體(1∶500)作為一抗,羊抗兔HRP-IgG(1∶5000)作為二抗,Western blotting檢測抗體特異性。HFF細胞提取的蛋白為陰性對照。
2 結 果
2.1生物學信息分析 通過Blast軟件發現一個TgSir2基因(ToxoDB:TGGT1_227020),其在ToxoDB數據庫中的基因注釋為組蛋白去乙酰化酶。利用ClustalW進行同源比對,發現雖然TgSir2與酵母、人Sir2相似性較低,但其酶活性中心卻高度保守,尤其是與瘧原蟲Sir2蛋白相似性最高(圖1A)。用Signal 4.1軟件對其表達的蛋白進行信號肽預測,發現TgSir2蛋白存在一段長度為15個氨基酸的信號肽(圖1B)。
2.2TgSir2基因的PCR擴增結果 TgSir2△1-15aa基因片段的理論大小為1 038 bp,PCR產物經瓊脂糖凝膠電泳后,在約1 000 bp處可見一條清晰的特異性擴增條帶,與TgSir2△1-15aa基因片段的理論大小(1 038 bp)一致(圖2)。
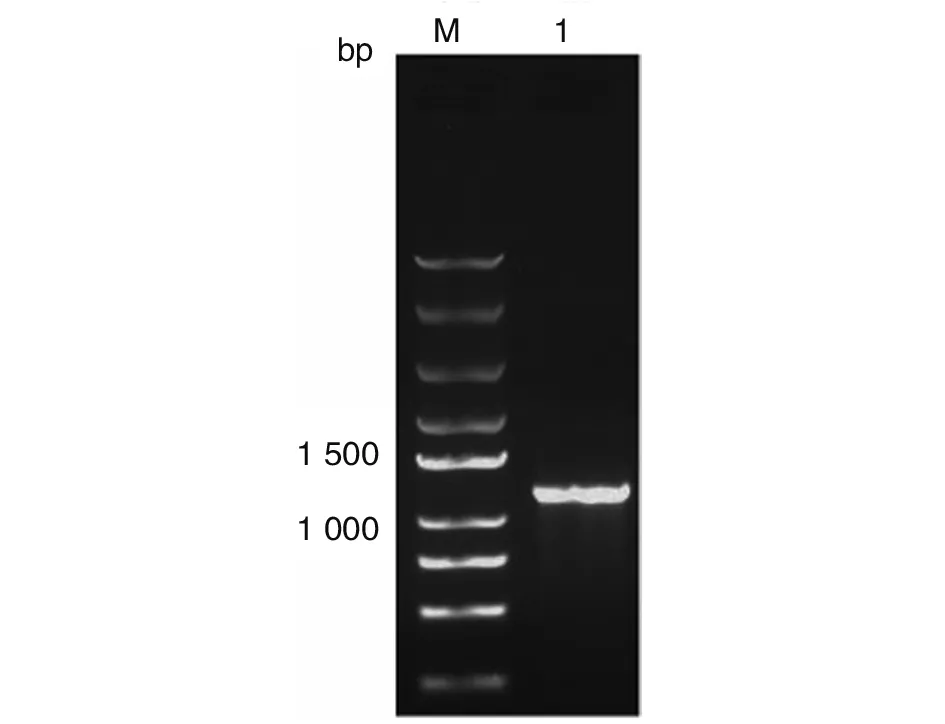
M:DNA標志物;1:TgSir2△1-15aa基因PCR產物
2.3重組質粒pET28b-TgSir2△1-15aa的鑒定 重組質粒pET28b-TgSir2△1-15aa經PCR擴增和雙酶切鑒定,目的基因片段均約為1 000 bp(圖3),測序表明pET28b-TgSir2質粒構建正確。
2.4TgSir2融合蛋白的表達、純化和鑒定 將鑒定正確的重組質粒pET28b-TgSir2轉化至BL21表達菌中,以IPTG誘導表達后,經鎳柱純化,SDS-PAGE檢測可見大量目的蛋白,其蛋白大小符合預

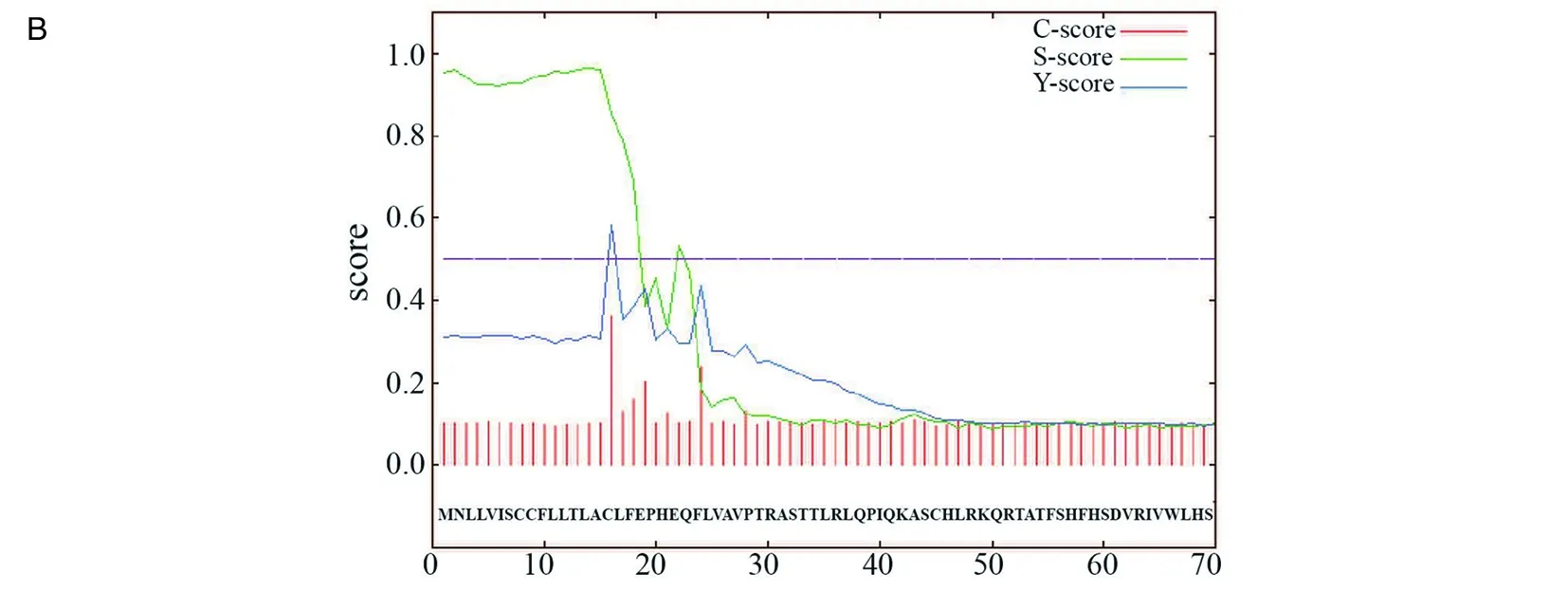
a表示構成酶活性的關鍵氨基酸殘基;b表示NAD+結合位點;c表示既可結合NAD+,同時也是酶活性關鍵氨基酸殘基;d表示構成鋅指結構的關鍵氨基酸殘基。
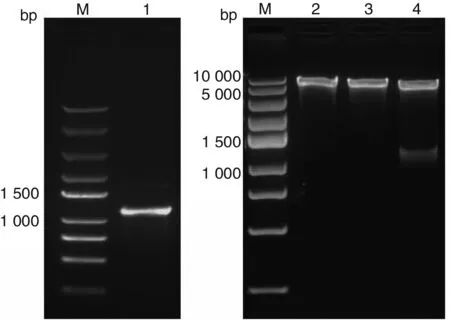
M:DNA標志物;1:陽性克隆PCR產物;2:pET28b;3:pET28b-TgSir2重組質粒;4:pET28b-TgSir2重組質粒雙酶切
期(圖4A)。隨后經Western blotting檢測,TgSir2融合蛋白能被小鼠抗His單克隆抗體識別,在相應分子量大小處有特異條帶(圖4B)。
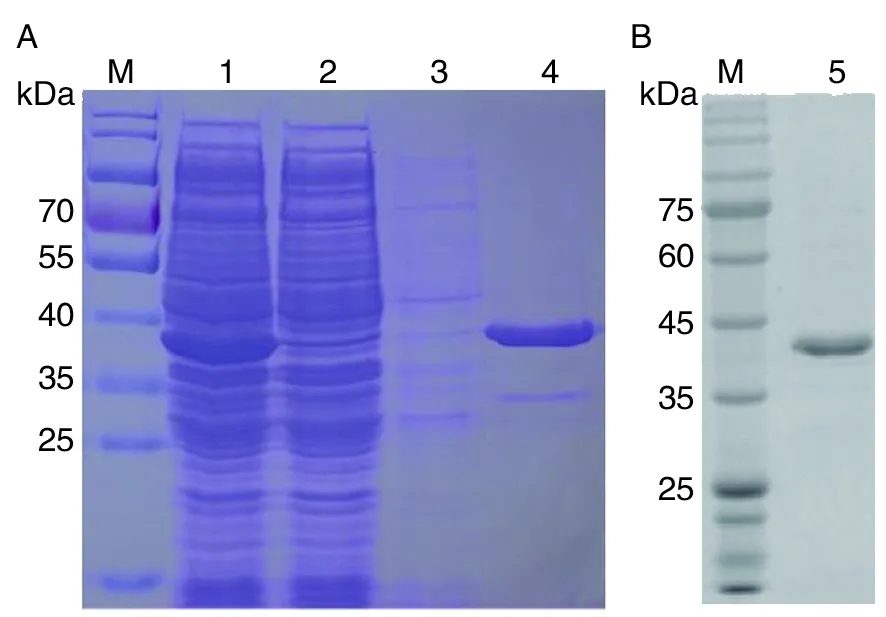
M:蛋白質標志物;1:誘導后樣品;2:誘導前樣品;3:PBS洗脫液;4:純化后融合蛋白;5. 純化后融合蛋白
2.5兔抗TgSir2多克隆抗體鑒定 經4次免疫后,以免疫前兔血清為對照,ELISA法測定2只免疫兔抗His-TgSir2血清效價均達1∶64 000。將2只兔血清收集后經親和層析純化,Western blotting檢測顯示,所制備的抗體能夠特異性識別原核表達
的TgSir2融合蛋白(圖5)。
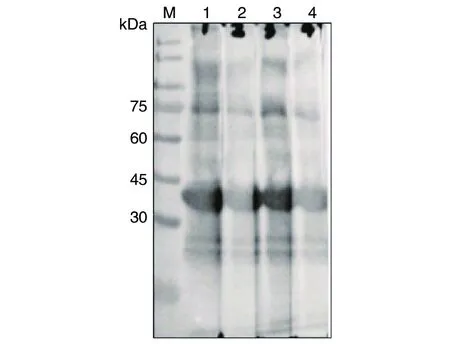
M:蛋白質標志物;1:1號免疫兔抗TgSir2多克隆抗體作為一抗(1∶8 000);2:1號免疫兔抗TgSir2多克隆抗體作為一抗(1∶16 000);3:2號免疫兔抗TgSir2多克隆抗體作為一抗(1∶8 000);4:2號免疫兔抗TgSir2多克隆抗體作為一抗(1∶16 000)
2.6評價兔抗TgSir2多克隆抗體識別弓形蟲內源性TgSirt蛋白的能力 提取弓形蟲全蟲蛋白,經SDS-PAGE與Western blotting檢測,發現兔抗TgSir2多克隆抗體不但可以與預期大小的目的條帶(圖6箭頭所指處)特異性結合,同時在70 kDa左右也有一條反應條帶。而宿主細胞對照組作為陰性對照,與兔抗TgSir2無特性性結合(圖6)。
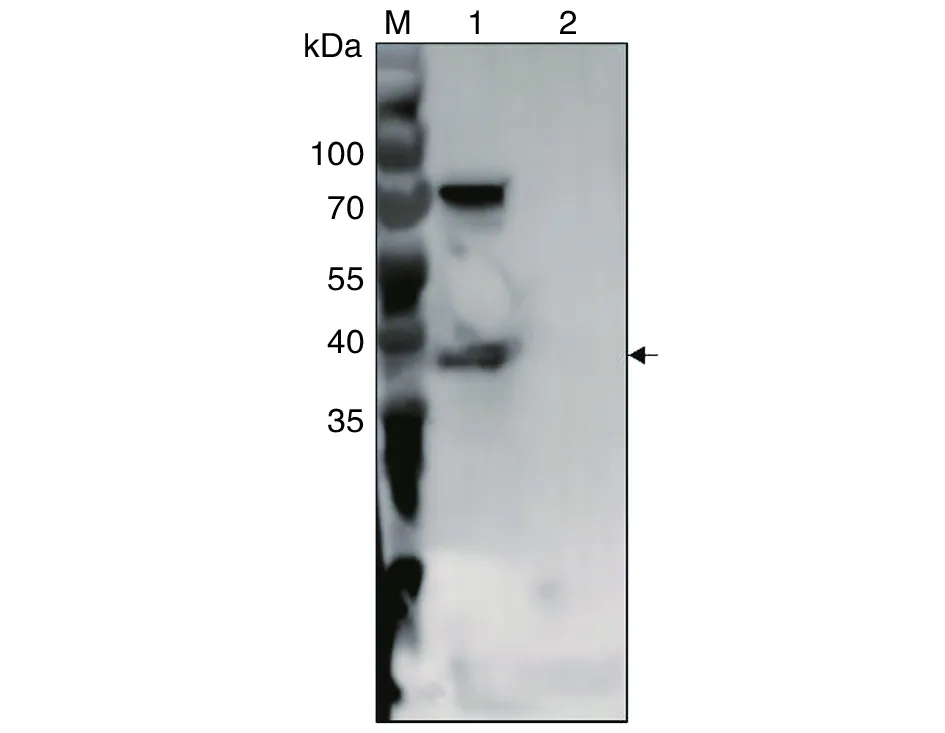
M:蛋白質標志物;1:弓形蟲速殖子全蟲蛋白;2:HFF細胞蛋白
3 討 論
Sir2同源物目前已在溶組織內阿米巴[8]、錐蟲[9-11]、利什曼原蟲[12-14]、瘧原蟲[15-16]等多種寄生性原蟲體內被發現,其功能涉及修復損傷DNA[11]、參與蟲體入侵與增殖[10, 17]、調節宿主免疫應答[11-12]、調控蟲體抗原變異基因從而逃避宿主免疫應答[18]等。因此,在多種寄生性原蟲病防治中,Sir2蛋白是一個具有研發前景的藥物、疫苗靶點[18-22]。在本實驗室前期研究過程中發現,TgSir2蛋白無法利用傳統的double-crossover策略進行基因敲除,提示該蛋白是蟲體生長發育過程中的必需蛋白。
Sir2結構中包含了保守的NAD+結合域和2個酶活性核心域:Rossmann折疊構成大結構域、鋅指結構(CX2CX20CX2C)構成小結構域。在2個結構域之間的縫隙中,乙酰化肽段與NAD+結合形成酶-底物結構而發生催化反應[3]。通過蛋白質同源性分析,我們發現,雖然TgSir2蛋白與其他種屬Sir2蛋白的序列相似度較低,但是核心功能域高度保守。這預示著TgSir2在弓形蟲中可能具有調節蛋白質去乙酰化的功能。隨后,我們通過信號肽預測發現,TgSir2蛋白可能作為一種分泌蛋白發揮作用。而這一信號肽在其他物種Sir2蛋白中尚未發現。
本實驗原核表達并純化了重組TgSir2蛋白,并以此為抗原成功制備了效價達1∶64 000的兔抗TgSir2多克隆抗體,適合進行后續實驗。進一步的Western blotting實驗結果顯示,該多抗既可識別原核表達的TgSir2融合蛋白,也可識別弓形蟲體內的內源性TgSir2蛋白。然而,在多抗識別內源性TgSir2蛋白的Western blotting實驗中,我們發現該抗體與蟲體蛋白反應后,不但出現了預期大小的目的條帶,還出現了一條分子量更大的蛋白條帶。盡管經過多次重復實驗,該條帶仍然存在,而對照組宿主細胞中卻并未檢測到任何條帶,因此,我們認為本研究中所制備的抗體是特異針對弓形蟲蛋白的多克隆抗體。由于該條帶的大小接近于目的條帶的兩倍,我們推測可能由以下3個原因:1)TgSir2在弓形蟲體內很可能以二聚體的形式存在并發揮生物學功能;2)作為一種修飾酶,TgSir2在弓形蟲生長發育過程中與某種蛋白發生了共價結合,從而發揮作用;3)該抗體除了可以識別弓形蟲TgSir2蛋白,還可能與蟲體內某種分子量約70 kDa的蛋白結合。以上推測,我們將在后續研究中進一步通過親和層析與質譜分析進行驗證。
由于本實驗已證實所制備的兔抗TgSir2多克隆抗體可識別弓形蟲內源性TgSir2,這將為進一步研究弓形蟲內的TgSir2分子機制及去乙酰化修飾機制提供了必要的實驗工具。
利益沖突:無
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
