代謝工程改造大腸桿菌合成酪醇
2019-02-06 07:22:14薛宇翔陳獻忠
食品與生物技術學報 2019年10期
薛宇翔,陳獻忠*,楊 翠,沈 微,樊 游
(1.江南大學,工業生物技術教育部重點實驗室,江蘇 無錫 214122;2.江南大學 生物工程學院,江蘇 無錫 214122)
酪醇,即4-(2-羥基乙基)苯酚,作為一種重要的芳香族化合物,是天然的抗氧化劑,也是苯乙醇的衍生物[1]。酪醇天然存在于橄欖油、綠茶及酒中[2-4],它可以保護細胞免受氧化傷害。酪醇及其衍生物具有多方面的生理學活性[5],如抗疲勞、抗缺氧、抗應激、抗寒冷、鎮靜等的藥理作用[2,6],進而運用在相關藥物(主要是治療心血管疾病的藥物)的研發生產上[7]。隨著對酪醇的生理功能的研究進一步加深,酪醇的生產也受到越來越多的關注。
現階段,酪醇制備方法主要是天然材料提取法和化學法合成法。對于天然材料提取來說,酪醇可以從橄欖或橄欖油制備過程中產生的廢水中提取分離純化獲得。天然提取制備酪醇存在很多弊端,對于橄欖油提取酪醇,其回收率和純度極低,成本投入高,這些問題在工業大規模化生產應用時會帶來很大的阻力[8]。化學合成法是主要以對羥基苯乙烯、對羥基苯乙胺等為原料合成酪醇,生產原料價格昂貴,且在生產過程中會產生對環境有害物質,這些問題制約了大規模化、工業化生產[9-10]。
近年來,研究者嘗試通過生物發酵法來合成酪醇,由于生物合成有能夠減少對環境的污染及方便后續酪醇的提取,同時不依賴于植物資源等優點,備受關注。在自然界中,存在兩種生物合成途徑,見圖1,一種是釀酒酵母生物合成途徑:酪氨酸(TYR)經過芳香族氨基酸氨基轉移酶(ARO8)作用脫去氨基,再經丙酮酸脫羧酶(ARO10)的作用脫羧,最后在乙醇脫氫酶的作用下合成酪醇[11-13];一種是酪氨酸經酪氨酸脫羧酶(TDC)的作用生成酪胺,酪胺進一步在酪胺氧化酶(TYO)和乙醇脫氫酶(ADH)的作用下生成酪醇[1]。研究者在釀酒酵母中過表達ARO8及ARO10 基因,以苯丙氨酸為底物,可以提高的苯乙醇合成能力,重組菌的苯乙醇的產量比野生型的提高了36.8%[14]。為了構建產紅景天苷的大腸桿菌,研究者在大腸桿菌異源表達ARO10 和葡萄糖基轉移酶等關鍵基因,并進行代謝途徑改造,重組菌能夠合成紅景天苷和其前體物質酪醇[11]。
本研究在E.coli 中異源表達ARO10 基因,并通過敲除預苯酸脫水酶編碼基因pheA 和苯乙醛脫氫酶編碼基因feaB,阻斷了苯丙氨酸途徑及4-羥基苯乙酸合成途徑,改善了酪醇的合成能力,通過培養條件的優化,提高了酪醇的產量。在外源添加酪氨酸的發酵培養基中,重組菌合成酪醇的能力有所提高。
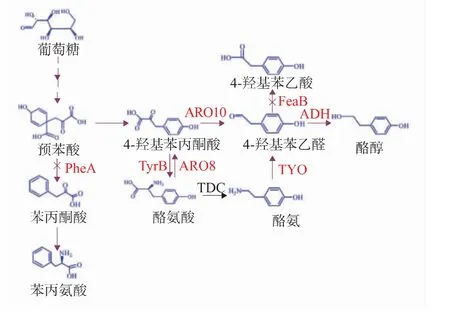
圖1 酪醇的生物合成途徑Fig.1 Biosynthesis pathway of tyrosol
1 材料與方法
1.1 材料
1.1.1 菌株、質粒 菌株:大腸桿菌(Escherichia coli BL21(DE3))、E.coli JM109,釀酒酵母(Saccharomyces cerevisiae EBY100)均為江南大學生物資源與生物能源研究中心保藏。
質粒:pRSFDuet-1、pKD13、pKD46、pCP20 由本實驗室保藏。
1.1.2 主要試劑 限制性內切酶、T4 DNA 連接酶、Taq DNA 聚合酶及蛋白分子量marker 均購自TaKaRa 生物技術公司。B 型質粒少量快速提取試劑盒及DNA 純化與割膠回收試劑盒均購自北京博大泰克生物基因技術有限公司。氨芐青霉素(Amp)、卡那霉素(Kan)與異丙基硫代半乳糖苷(IPTG)購自上海生工生物工程有限公司,酪氨酸和酪醇標準品購自SIGMA(上海)公司,其他的分析純級試劑購自上海生物工程公司。
1.2 方法
1.2.1 培養基和培養條件 LB 培養基:酵母粉0.5%,蛋白胨1%,氯化鈉1%,固體培養基加瓊脂粉2%,121 ℃下滅菌20 min。
M9Y 培養基:葡萄糖1%,酵母粉0.025%,七水磷酸氫二鈉1.28%,磷酸二氫鉀0.3%,氯化鈉0.05%,氯化銨0.1%,115 ℃下滅菌15 min。轉接時需添加分開滅菌的硫酸鎂溶液,其最終濃度應為5 mmol/L。
種子培養:劃單菌落到LB 液體培養基,在37 ℃、200 r/min 的條件下培養約12 h。用紫外分光光度儀測定種子培養液的OD600,轉接1 mL 種子培養液到50 mL 的LB 液體培養基中。
誘導培養:將培養基在37 ℃、200 r/min 的條件下培養,控制培養液OD600在0.6~0.8,添加一定量的IPTG,在30 ℃、200 r/min 的條件下培養10 h。
發酵培養:將誘導表達后的培養液于4 ℃、7 000 r/min 的條件下離心3 min,并用無菌水離心洗滌,收集菌體。轉接入M9Y 培養基在30 ℃、200 r/min的條件下,發酵培養48 h。
1.2.2 目的基因的克隆 本研究中所用引物見表1,引物由上海生工生物工程技術服務部合成。根據NCBI 公布的釀酒酵母(S.cerevisiae EBY100)丙酮酸脫羧酶基因ARO10(GenBank ID:851987)序列設計引物ARO10U 和ARO10D。PCR 擴增參數:95℃變性5 min,然后94 ℃變性30 s,56 ℃退火30 s,72 ℃延伸1.5 min,30 個循環后16 ℃保溫30 min,得到目的基因ARO10。

表1 基因PCR 擴增引物列表Table 1 List of primers in this study
1.2.3 重組表達載體的構建 將ARO10 基因PCR產物與pRSFDUet-1 質粒同時用NcoI 與BamHI 雙酶切后,純化處理,用T4 DNA 連接酶進行連接,16℃反應12 h,得到表達載體pRSFDUet-ARO10,轉化E.coli BL21(DE3)感受態細胞,提取質粒酶切鑒定。
1.2.4 feaB、pheA 和tyrB 基因的敲除 制備含有pKD46 的E.coli BL21(DE3)電轉化感受態細胞[15],將由引物YfeaBU 和YfeaBD(表1)擴增獲得的DNA 片段(feaB 上下游同源序列、FRT 位點和kan基因),電轉化入含有pKD46 的E.coli BL21(DE3)感受態細胞,電轉化條件為2 mm 電轉杯、電壓2 500 V,電轉化后37 ℃下培養2 h,涂布于Amp 和Kan 雙抗性LB 平板上。突變盒片段進入細胞后,在pKD46 輔助質粒所表達的重組蛋白的幫助下,同源臂基因與大腸桿菌基因組發生同源重組,使feaB 基因區域被敲除框替換,即feaB 被kan 替換,因此,重組菌可在Kan 抗性平板上生長。平板培養12 h 后挑取單菌落,用鑒定性引物NfeaBU 和NfeaBD(表1)PCR 擴增,通過確定敲除框上下游是否正確交換來鑒定的敲除。之后將pCP20 質粒轉入E.coli BL21(DE3)ΔfeaB::kan 菌株中,42 ℃培養12 h,取菌液稀釋,涂布于LB 固體平板。pCP20 質粒含編碼FLP 重組酶的基因,可介導2 個FRT 位點間的切除,從而消除kan 基因。pCP20 為溫敏型質粒,培養溫度高于37℃時會自動丟失,因此挑取單菌落分別點到Kan 抗性LB 平板和LB 平板上培養12 h,挑取在LB 固體培養基平板上生長而Kan 抗性平板上不生長的菌落,即為E.coli BL21(DE3)ΔfeaB。
制備含有pKD46 的E.coli BL21(DE3)ΔfeaB的感受態細胞,將由引物YpheAU 和YpheAD(表1)擴增獲得的DNA 片段(含pheA 上下游同源序列、FRT 位點和kan 基因),利用同上的電轉化技術,轉入含有pKD46 的E.coli BL21(DE3)ΔfeaB 的感受態細胞。利用同上的方法,最終得到了E.coli BL21(DE3)ΔfeaBΔpheA。
制備含有pKD46 的E.coli BL21(DE3)的感受態細胞,將引物YtyrBU 和YtyrBD(表1)擴增獲得DNA 片段(含tyrB 上下游同源序列、FRT 位點和kan 基因),利用電轉化技術,轉入含有pKD46 的E.coli BL21(DE3)的感受態細胞,經標記基因彈出后,最終獲得E.coli BL21(DE3)ΔtyrB。
1.2.5 酪醇和酪氨酸的高效液相測定 使用高效液相色譜(HPLC)。色譜條件如下:HPLC 色譜柱為COSMOSIL-5C18-MS-Ⅱ(3.0 ID×150 mm)[16]。流動相A 為0.1%三氟乙酸,流動相B 為純甲醇,柱溫為30 ℃,紫外檢測器,流速設置為0.4 mL/min。流動相需用0.22 μm 孔徑的水系膜進行真空抽濾,使用前超聲10 min 除去氣泡。
樣品處理:取發酵液100 mL,12 000 r/min 離心5 min 收集上清,經0.22 μm 孔徑濾膜過濾后加入HPLC 樣品瓶對發酵液中的酪醇和酪氨酸含量進行檢測[17]。
2 結果與分析
2.1 產酪醇的重組大腸桿菌構建
2.1.1 重組質粒的構建 以釀酒酵母(S.cerevisiae EBY100)染色體DNA為模版,PCR擴增得到ARO10 基因片段,目的基因經NcoI 與BamHI 酶切純化后插入到表達載體pRSFDUet-1 的相應酶切位點上,獲得重組表達載體pRSFDUet-ARO10,酶切驗證(如圖2 所示),ARO10 片段大小為1 908 bp,與預期結果一致,證明表達載體pRSFDUet-ARO10構建成功。
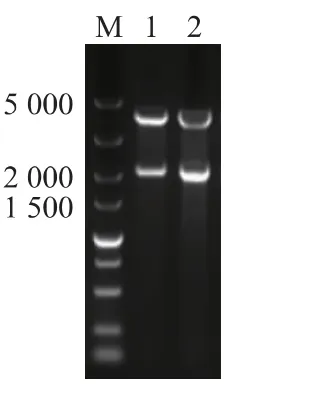
圖2 酶切產物電泳檢測圖Fig.2 Enzyme-digested product of the recombinant plasmid pRSFDUet-ARO10
2.1.2 重組菌發酵生產酪醇 將pRSFDUet-1 質粒和重組表達載體pRSFDUet-ARO10 分別導入E.coli BL21(DE3)中,獲得重組菌BE0 和重組菌BE1,兩株重組菌發酵培養48 h,發酵結果如圖3 所示。由圖3 可知,重組菌BE0 發酵液中未檢測到酪醇,可見大腸桿菌本體內不能合成酪醇。重組菌株BE1的酪醇產量達到0.35 mmol/L,可知通過丙酮酸脫羧酶編碼基因ARO10 異源表達,可以建立E.coli 合成酪醇的途徑。另一方面,雖然重組質粒進行游離表達,而且帶有T7 的強啟動子,但是酪醇的產量還是較低的,這說明重組E.coli 從葡萄糖到酪醇的合成途徑較長,而且芳香族氨基酸合成途徑中存在的反饋調節,也可能阻礙酪醇的合成。
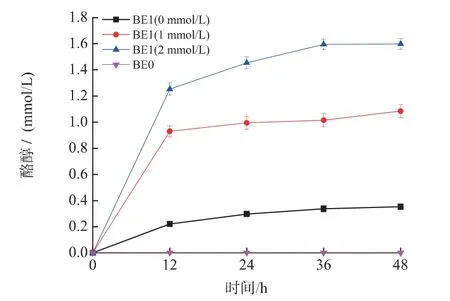
圖3 外源添加酪氨酸對重組菌BE1 酪醇合成的影響及重組菌BE0 發酵生產酪醇過程Fig.3 Effect of supplementing tryrosine on the tryrosol production for the recombinant strain BE1 and the course of producing tyrosol by the recombint strain BE0
2.2 外源添加酪氨酸對重組菌BE1 酪醇合成的影響
由圖1 代謝途徑可知,酪氨酸和酪醇合成過程中需要利用相同的前體—4-羥基苯丙酮酸(4HPP)。因此,推測胞內的酪氨酸濃度可能會影響酪醇的合成。通過添加不同濃度(0、1.0、2.0 mmol/L)的酪氨酸考察了重組菌BE1 對酪醇的產量的影響。發酵結果及酪醇產量提高率如圖3 和表2 所示。
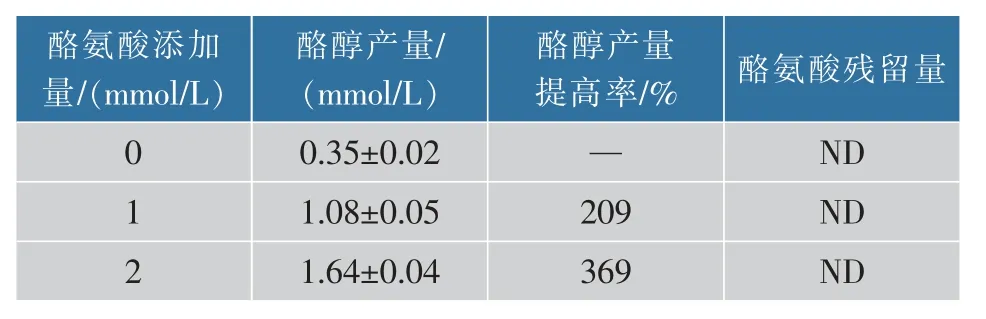
表2 添加不同濃度酪氨酸時重組菌BE1 合成酪醇的產量Table 2 Tyrosol production of different concentration tyrosine sumplemented in M9Y for BE1
從圖3 中可以得出,發酵培養基中添加不同濃度的酪氨酸,酪醇的產量都有了較大的提高。發酵液中未檢測到酪氨酸,說明發酵液中幾乎沒有酪氨酸殘留。從代謝途徑上可以推測重組菌BE1 中大腸桿菌本體內基因tyrB 可能導致酪氨酸生成4-羥基苯丙酮酸,進而產生了酪醇。根據以上結果,決定保留該菌株的酪氨酸合成途徑。
2.3 改造旁路代謝途徑并提高酪醇產量
2.3.1 敲除基因feaB 和pheA 由代謝途徑(圖1)分析可知,敲除feaB 和pheA 基因,可以阻斷4-羥基苯乙酸的合成途徑和苯丙氨酸代謝途徑,推測可能提高酪醇產量。因此,進一步構建含有feaB 基因同源臂和kan 抗性基因的刪除盒并轉化E.coli BL21(DE3)菌株,篩選轉化子并經誘導彈出kan 抗性基因。突變株的PCR 鑒定結果如圖4(a)所示,原始菌株PCR 產物大小為1 780 bp,抗性基因彈出后的突變株PCR 產物大小為398 bp,與預期結果一致,證明成功敲除了feaB 基因。在敲除feaB 基因的基礎上,敲除pheA 基因,阻斷苯丙氨酸代謝途徑。將kan 抗性基因整合E.coli BL21(DE3)ΔfeaB 菌株的pheA 基因位點,轉入pCP20 質粒消除Kan 抗性后轉化子驗證,結果如圖4(b)所示,原始序列為1 658 bp,突變株499 bp。與預期結果一致,證明成功敲除了pheA 基因。
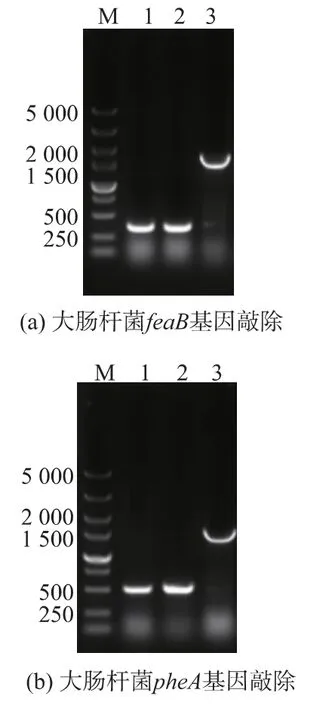
圖4 PCR 驗證Fig.4 PCR identification
2.3.2 基因敲除后對酪醇產量的影響 將構建好的表達載體pRSFDUet-ARO10 導入E.coli BL21(DE3)ΔfeaB 和E.coli BL21(DE3)ΔfeaBΔpheA,獲得重組突變菌株BE3 和BE4。將重組菌BE1、BE3和BE4 發酵48 h,收集發酵液處理后檢測酪醇,結果如圖5 所示。
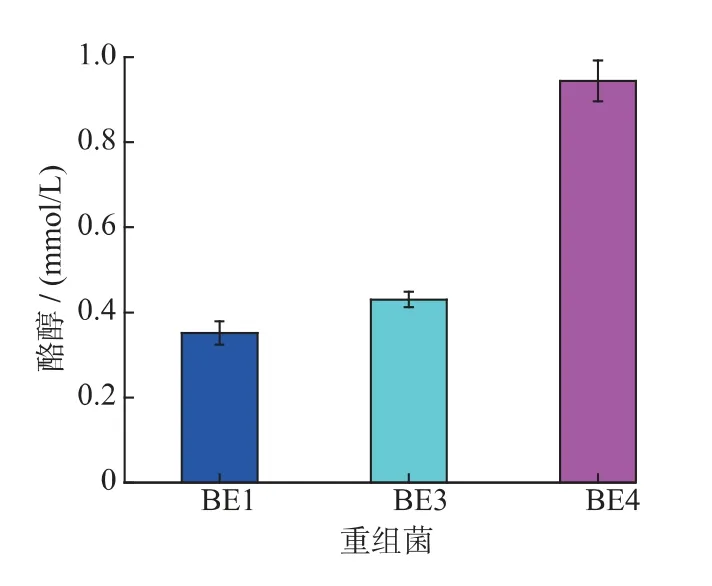
圖5 基因敲除對酪醇產量的影響Fig.5 Effect of gene deletion on tryrosol production
從圖5 中可以看出,對于菌株BE3 來說,酪醇產量達到0.41 mmol/L,可知,敲除feaB 基因,較BE1 提高了17.1%,說明敲除feaB 基因,有助于提高酪醇的合成能力;對于菌株BE4 來說,可產酪醇0.91 mmol/L,較BE1 提高了160%,說明在敲除feaB基因基礎上,敲除pheA 基因,可顯著提高酪醇的合成能力。進而說明敲除feaB 和pheA 基因,阻斷了4-羥基苯乙酸與苯丙氨酸生成途徑,使葡萄糖更多地流向酪醇成途徑,從而提高了酪醇的合成能力。
2.4 重組突變菌株BE4 產酪醇條件的優化
2.4.1 重組蛋白誘導溫度及誘導時機的優化 溫度對酶的活性及其表達有著極大的影響[18],考察了不同誘導溫度(20、25、27、30 ℃對酪醇產量的影響,結果如圖6(a)所示。從圖中6(a)可以看出誘導溫度在25 ℃時,酪醇的產量最高,產量可達3.61 mmol/L。誘導時機對重組蛋白表達有重要影響。考察了在不同OD 值(0.6、0.9、1.2、1.5、1.8、2.1)時誘導劑IPTG 誘導蛋白表達對酪醇產量的影響,結果如圖6(b)所示。圖6(b)表明OD 值高于0.6 時并不利于酪醇的產生,當OD 值在0.6~0.8,大腸桿菌處于對數生長期,生理狀態良好,更利于最終酪醇的產生。最終酪醇產量可達3.48 mmol/L。
2.4.2 誘導劑濃度的優化 誘導劑濃度的大小可以起到促進或抑制的作用,所以適當濃度的誘導劑是十分關鍵的。考察了不同IPTG 濃度(0.2、0.4、0.6、0.8、1.0、1.2 mmol/L)對酪醇產量的影響,結果如圖6(c)所示。由圖中6(c)可以看出當IPTG 濃度在0.8 mmol/L時,酪醇產量最低,當IPTG 濃度在0.2 mmol/L 時,酪醇產量最高,產量可以達到4.15 mmol/L。
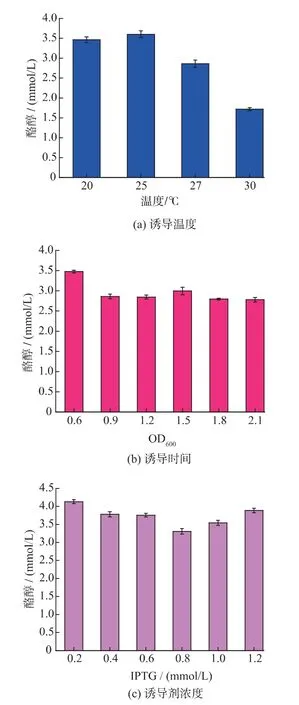
圖6 重組菌誘導條件對酪醇合成的影響Fig.6 Optimization of induction conditions for tyrosol produciton
2.5 基因tyrB 對酪醇合成的影響
2.5.1 基因tyrB 的刪除 為刪除E.coli BL21(DE3)菌株的tyrB 基因,構建含有同源臂和Kan 抗性基因的刪除盒,將其轉化E.coli BL21(DE3)并整合到基因組上,篩選轉化子并經誘導彈出Kan 抗性基因。野生型與突變型轉化子菌落PCR 驗證,原始菌株PCR 產物大小為1 059 bp,抗性基因彈出后的突變株PCR 產物大小為254 bp。結果如圖7 所示,與預期的大小一致,證明基因tyrB 已成功敲除。
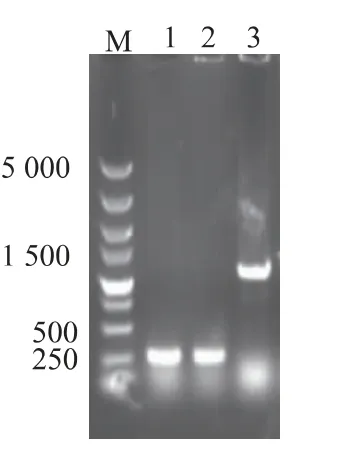
圖7 大腸桿菌tyrB 基因敲除的PCR 驗證Fig.7 PCR identification of the gene tyrB deletion in E.coli strain
2.5.2 刪除基因tyrB 對重組菌合成酪醇的影響將構建好的表達載體pRSFDUet-ARO10 導入E.coli BL21(DE3)ΔtyrB 中,獲得重組大腸桿菌BE2。將兩株重組菌BE1 和BE2 經種子液培養、LB 液體培養、IPTG 誘導表達,分別轉接外源添加不同濃度(0、1、2.8 mmol/L)酪氨酸的M9Y 培養基中,發酵培養48 h,發酵結果如圖8 所示。
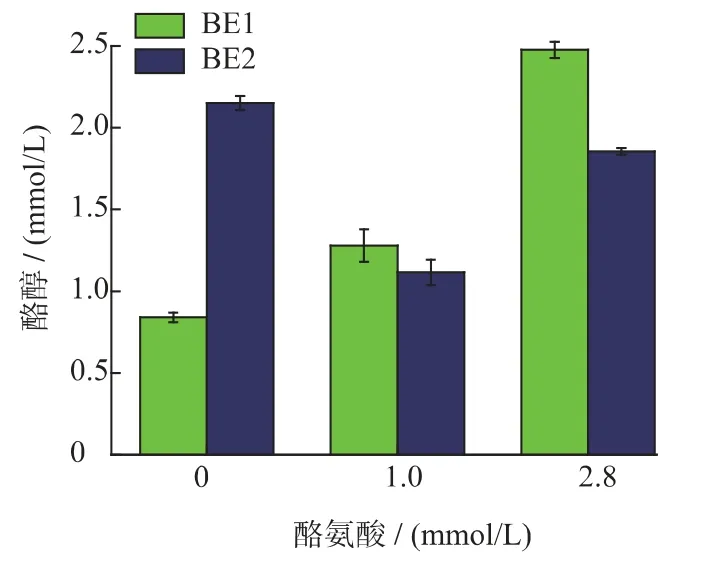
圖8 外源添加酪氨酸對重組菌BE1 和重組菌BE2 酪醇合成的影響Fig.8 Effect of supplementing tryrosine on the tryrosol production for the recombinant strain BE2 and BE1
從圖8 可以看出,發酵培養基不添加酪氨酸時,BE1 可產酪醇0.86 mmol/L,BE2 可產酪醇2.18 mmol/L,BE2 的合成能力高于BE1,可知,刪除基因tyrB 可以提高重組菌酪醇的合成能力。從圖6 可以看出,發酵培養基外源添加2.8 mmol/L 的酪氨酸與1 mmol/L 酪氨酸相比,重組菌BE1 和BE2 的合成能力均有提高。發酵液中未檢測到酪氨酸,說明發酵液中幾乎沒有酪氨酸殘留。可知,大腸桿菌胞內存在多條催化酪氨酸合成4-羥基苯丙酮酸的途徑。
3 討論
本研究通過在E.coli BL21(DE3)中異源表達ARO10 基因,獲得了可以合成酪醇的重組大腸桿菌。通過刪除旁路代謝途徑及對培養條件優化,利用10 g/L 葡萄糖作為碳源,發酵48 h 酪醇產量可達4.15 mmol/L。Yasuharu Satoh 通 過 在E.coli MG1655 中異源表達TYO 及TDC 基因,重組菌利用10 g/L 葡萄糖作為碳源,搖瓶發酵48 h,酪醇產量可達0.5 mmol/L[1]。文獻[11]通過在E.coli MG1655中異源表達ARO10 基因,并對代謝途徑進行了改造,利用20 g/L 葡萄糖作為碳源,酪醇產量可達5.4 mmol/L。與之相比,本研究僅對酪氨酸合成途徑上的個別基因進行改造,重組菌合成酪醇的能力還有進一步的提升空間。本研究中ARO10 基因來源于釀酒酵母,在大腸桿菌的表達效率相對較低。下一步將對ARO10 基因進行密碼子優化,進一步改善酪醇的合成能力。
由于大腸桿菌體內酪醇的前體物質4-羥基苯丙酮酸合成途徑較長[15],對于增強酪醇的合成能力,在未來還有很多研究要做。由酪醇的代謝途徑可知(圖1),4-羥基苯丙酮酸也是合成酪氨酸的前體物質,下一步將采用過量合成酪氨酸的菌株合成酪醇。過去幾十年里,有關于過量合成酪氨酸的菌株的研究成一直是研究熱點[19-21]。理論推測,利用產10 g/L 酪氨酸的工程化的大腸桿菌可以產酪醇7.6 g/L。這個產量比從橄欖油中提取得到的酪醇產量高3 倍。因此,利用合成酪氨酸能力強的宿主菌以葡萄糖為底物來提高重組菌酪醇的合成能力,將成為下一步的研究重點。
4 結語
在外源添加酪氨酸的發酵培養基中培養合成酪醇,可以改善酪醇的合成能力,推測大腸桿菌體內存在催化酪氨酸合成4-羥基苯丙酮酸的酶。下一步,將進一步探索以酪氨酸為底物的全細胞催化合成酪醇的條件。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中國果業信息(2019年10期)2019-11-13 01:21:34
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國衛生(2016年3期)2016-11-12 13:23:26
