共表達抗氧化酶促進脂肪氧合酶在大腸桿菌中的表達
2019-02-06 07:21:40邱芳芳堵國成
食品與生物技術學報 2019年10期
邱芳芳,劉 松,堵國成*,陳 堅
(1.江南大學 工業生物技術教育部重點實驗室,江蘇 無錫 214122;2.江南大學 生物工程學院,江蘇 無錫 214122)
脂肪氧合酶(EC1.13.11.12,LOX)是一類能夠專一催化多元不飽和脂肪酸(含順-1,4-戊二烯結構)形成具有共軛雙鍵的脂肪酸氫過氧化物的酶[1-2]。此類酶分子一般含有非血紅素鐵。LOX 作為一種無毒無害的生物制劑,可替代溴酸鉀和苯甲酰過氧化物等化學增白劑提高面粉白度。此外,LOX 能夠改善面粉筋力,改善面團的水合力,增加面包體積,改善面包質構。LOX 催化不飽和脂肪酸的氧化產物可進一步轉化為己醇、己醛、壬烯醛等風味化合物。
目前商業化的LOX 產品由大豆提取。然而,由于大豆中存在多種同工酶,提取法獲得的LOX 產品批次穩定性差,不利于最終產品質量的控制和大規模應用。目前,大豆、豌豆、人、豬等真核細胞生物來源的LOX 已經在大腸桿菌,畢赤酵母或釀酒酵母中獲得活性表達,但胞內產酶水平最高僅為4.5 U/mL。最近的報道顯示,魚腥藻(Anabaena sp.PCC 7120)LOX 在枯草芽孢桿菌中成功表達,但按標準酶活力定義計算,其胞外LOX 產量僅為0.01 U/mL[2]。本文作者所在團隊將銅綠假單胞菌(Pseudomonas aeruginosa BBE)來源的LOX 基因至大腸桿菌(Escherichia coli),首次實現了LOX 在大腸桿菌中的高效分泌[1]。
一般認為,導致重組LOX 的總體表達水平不高的主要原因可能是活性LOX 產生的氧自由基會對宿主細胞產生毒性,進而抑制宿主細胞的生長和重組蛋白合成及分泌[1]。為降解活性氧中間體,細菌會產生超氧化物歧化酶(SOD)和過氧化氫酶(CAT)來消除活性氧中間體對細胞的毒害[3-6]。本研究通過共表達P.aeruginosa BBE 來源SOD(sodB、sodM)和E.coli 來源的CAT(katE),以期降低LOX 表達對細胞的毒性,進一步提高其在大腸桿菌中的表達水平。
1 材料與方法
1.1 材料
1.1.1 質粒與菌株 E.coli Rosetta(DE3)(pET-22b(+)/pre-LOXlox1)(N6)為分泌表達P.aeruginosa BBE來源LOX的重組E.coli Rosetta(DE3)[1]。pRSFDuet-1 為表達載體。E.coli JM109 分別用于基因克隆。
1.1.2 培養基 LB 培養基:蛋白胨10 g/L,酵母提取物5 g/L,NaCl 10 g/L。
發酵培養基(TB 培養基):蛋白胨12 g/L,酵母提取物24 g/L,甘油4 mL/L,K2HPO472 mmol/L,KH2PO417 mmol/L,誘導時培養基中添加終濃度為1 mmol/L 的IPTG 和1.5%(V/V)的吐溫20。
1.1.3 試劑 Taq DNA 聚合酶,T4 DNA 連接酶,限制性內切酶,DNA 和蛋白Marker 等購自寶生物工程(大連)有限公司。質粒抽提試劑盒購于生工生物工程(上海)股份有限公司。DNA 凝膠回收試劑盒,PCR 產物純化試劑盒購自賽默飛世爾科技公司,亞油酸為Sigma-Aldrich 公司產品。引物由深圳華大基因科技有限公司合成。其余常用試劑購于國藥集團化學試劑有限公司。
1.2 方法
1.2.1 基因的PCR 擴增 設計合成分別帶有NotⅠ和XhoⅠ兩種酶切位點的SodB、SodM、CAT 引物。以P.aeruginosa BBE 基因組為模板,以P1 和P2為上下游引物擴增5’端帶有NotⅠ和3’端帶有和XhoⅠ兩種酶切位點的sodB 基因;以P3 和P4 為上下游引物,擴增5’端和3’端分別帶有NotⅠ和XhoⅠ兩種酶切位點的sodM基因.以大腸桿菌JM109 基因組為模板,以P5 和P6 為引物,擴增5’端和3’端分別帶有NotⅠ和XhoⅠ兩種酶切位點的katE 基因。PCR 引物序列見表1。

表1 PCR 引物序列Table 1 PCR primer sequence
1.2.2 表達載體的構建及轉化 將上述PCR 產物分別克隆至pRSFDuet-1 上的NotⅠ和XhoⅠ之間,分別構建得到表達sodB,sodM 和katE 的重組質粒pRSF-sodB,pRSF-sodM 和pRSF-katE,3 個質粒分別轉化E.coli Rosetta(DE3)(pET-22b(+)/pre-LOXlox1)感受態細胞,冰浴30 min,42 ℃熱激90 s,再加入1 mL LB 培養基37 ℃振蕩培養1 h,涂布LB 平板(含50 μg/mL 氨芐霉素,10 μg/mL 卡那霉素和34 μg/mL 氯霉素),置于37 ℃培養箱中培養12 h。挑取若干LB 平板單菌落分別接種于3 mL LB液體培養基中(含50 μg/mL 氨芐霉素,10 μg/mL 卡那霉素和34 μg/mL 氯霉素),培養12 h,用質粒抽提試劑盒提取質粒,用表1 中的引物進行PCR 擴增,PCR 產物進行瓊脂糖凝膠電泳,以驗證各質粒是否已轉化成功,并進行測序.PCR 產物純化和凝膠回收按試劑盒說明書進行操作,基因的酶切和連接按試劑說明書進行操作。
1.2.3 共表達N6-B,N6-M 和N6-K 菌株的誘導表達 將表達質粒pRSF-sodB,pRSF-sodM 和pRSFkatE 轉化至表達LOX 的重組大腸桿菌N6,分別得到菌株N6-B,N6-M 和N6-K。從固體培養基上挑取原始菌株N6 及N6-B,N6-M 和N6-K 的單菌落,分別接種于裝有20 mL LB 的250 mL 三角瓶中,N6培養基中加入50 μg/mL 氨芐霉素(終濃度)和34 μg/mL氯霉素(終濃度),N6-B,N6-M 和N6-K 的培養基中加入50 μg/mL 氨芐霉素(終濃度),34 μg/mL 氯霉素(終濃度)和10 μg/mL 卡那霉素(終濃度),于37 ℃和200 r/min 培養過夜,按1%接種量接入裝有50 mL LB 的500 mL 三角瓶中,37 ℃,200 r/min 培養至OD600至0.6 左右,加入終濃度為1 mmol/L 的IPTG 和終濃度為1.5%的吐溫20,20 ℃誘導70 h[7]。
1.2.4 誘導條件優化 外源基因在工程菌中的誘導表達受到很多因素例如菌株類型,目的基因密碼子優化,載體類型,培養基,誘導條件(如誘導劑類型和濃度,誘導時間,誘導溫度)等的影響,因此確定還原酶在N6 中表達的優化條件對提高LOX 最終產量具有重要意義[8]。根據第一步的實驗結果,選取LOX 酶活最高的N6-M 重組菌株,由于N6-M 中的兩個質粒均為T7 啟動子,所以選取誘導劑濃度和誘導時菌體濃度兩個因素。采用L12(41×31)正交表,選定誘導時異丙基-β-D-硫代半乳糖苷(IPTG)濃度和菌體濃度(OD600)作為考察因子。其中IPTG的濃度為4 水平,OD600為3 水平,進行正交試驗。實驗以誘導30 h 胞內LOX 酶活為評價指標,實驗因素水平設置如表2 所示,選擇最佳條件和水平。
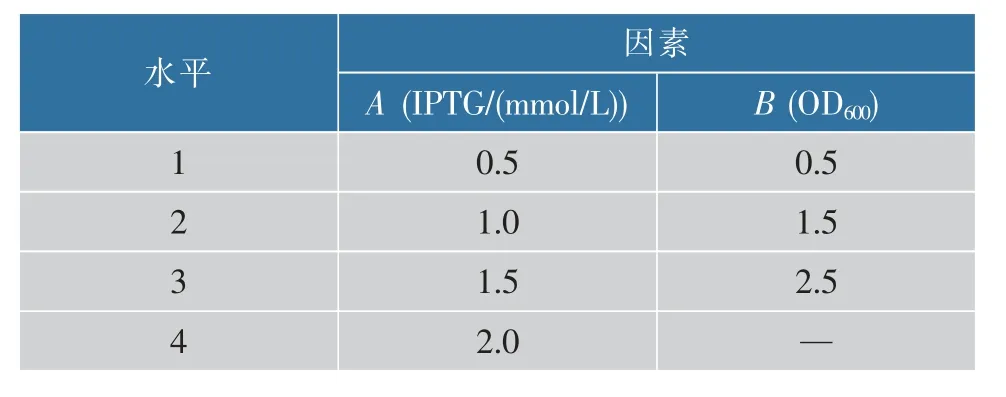
表2 誘導條件正交因素水平表Table 2 Orthogonal factor levels of induction conditions
1.2.5 重組菌株生長測定 將各共表達菌株和對照菌株接種至含有相應抗生素的LB 液體培養基,200 r/min,采用比濁法(OD600)測定菌體生長。
1.2.6 酶活檢測 LOX 酶活測定:在25 ℃下和234 nm 條件下以亞油酸為底物,使用Shimadzu UV-2450 分光光度計在線測定吸光值的變化,以吸光值變化曲線穩定上升部分的斜率計算酶活[7]。LOX 酶活單位定義:25 ℃下每分鐘催化亞油酸形成亞油酸氫過氧化物(HPOD,ε=25 000 L/(mol·cm))1 μmol所需要的酶量定義為1 個酶活單位(U)。
SOD 酶活測定:鄰苯三酚在堿性條件下的自氧化反應生成紅橘酚,同時有O2-產生,鄰苯三酚的自氧化速率與O2-的濃度有關;SOD 能通過催化O2-發生歧化反應得到H2O2和O2,來抑制鄰苯三酚的自氧化;檢測鄰苯三酚自氧化速率的抑制率,可得知樣品中的SOD 的含量;故,使用Shimadzu UV-2450分光光度計測定在25 ℃條件下鄰苯三酚在325 nm處吸光值變化曲線的斜率,計算酶活[6]。酶活單位定義:25 ℃時抑制鄰苯三酚自氧化速率50%時所需要的SOD 酶量為一個酶活力單位(U)。
CAT 酶活測定:在240 nm 波長下H2O2有強烈吸收;過氧化氫能被過氧化氫酶分解,從而反應溶液的吸光值(A240)隨反應時間下降,使用Shimadzu UV-2450 分光光度計測定該反應體系在25 ℃條件下240 nm 處的吸光值變化,以吸光值變化擬合斜率,計算酶活[6]。酶活單位定義:使0.1 mol/L H2O2在1 min 內A240減少0.1 的酶量為一個酶活單位(U)。
為了分析重組菌中SOD 和CAT 的表達情況,將不同時間點取的樣品離心收集菌體,用pH 7.5 的PBS 緩沖液漂洗一遍后重懸,超聲破碎,離心去上清得到胞內可溶物質。破碎后上清液檢測SOD 和CAT的酶活性。
1.2.7 SDS-PAGE 分析 將不同誘導時間的發酵液離心,0.5 mL 的菌體用磷酸緩沖液(PBS,pH 7.5)漂洗一遍,然后用1.5 mL PBS 懸浮,超聲破碎10 min,離心取上清。破碎后的胞內可溶物質和發酵液上清一起進行SDS-PAGE 分析。
2 結果
2.1 重組質粒pRSF-sodB,pRSF-sodM,pRSFkatE 的構建
研究顯示,超氧化物歧化酶和過氧化氫酶可以降解細胞內的活性氧中間體[3-6]。為考察抗氧化酶對LOX 表達的影響,將PCR 擴增得到的P.aeruginosa BBE 超氧化物歧化酶基因sodB 和sodM 以及E.coli 過氧化氫酶基因katE 克隆至pRSFDuet-1,分別得到表達質粒pRSF-sodB,pRSF-sodM 和pRSFkatE(圖1)。重組的pRSF-sodB、pRSF-sodM,pRSFkatE 經雙酶切后可以分別得到約600、600 bp 和2 400 bp 的DNA 片段。測序結果顯示DNA 序列無突變,證明重組表達載體構建成功,抗性平板上長出的即為陽性克隆。

圖1 重組質粒的構建Fig.1 Construction of recombinant plasmids
2.2 抗氧化酶共表達對LOX 表達的影響
將構建得到的抗氧化酶表達質粒pRSF-sodB,pRSF-sodM 和pRSF-katE(圖1)轉化LOX 表達菌N6,分別得到重組菌N6-B,N6-M 和N6-K。將重組菌進行搖瓶發酵,IPTG 誘導70 h。如圖2 所示,誘導70 h 后,N6、N6-B、N6-M 和N6-K 胞外LOX 酶活分別為10.3,2.9,3.1 和7.1 U/mL,胞內LOX 酶活分別為1.5,18.7,25.0 和0 U/mL。因此,N6-B、N6-M 和N6-K 誘導70 h 的LOX 總酶活分別為11.8,21.6,28.1 和7.1 U/mL,其中,N6-B 和N6-M的LOX 酶活分別較對照菌株N6 分別提高的1.8 和2.4 倍,N6-K 較對照菌株N6 下降40%,值得注意的是,當IPTG 誘導32 h,N6-M 中胞內LOX 酶活達到最大值27.2 U/mL,是N6 胞內LOX 酶活的3.1 倍。結果表明:共表達超氧化物歧化酶能有效促進胞內LOX 的高效合成,但降低了其向胞外分泌;共表達過氧化氫酶(katE)抑制了重組菌表達與分泌LOX。

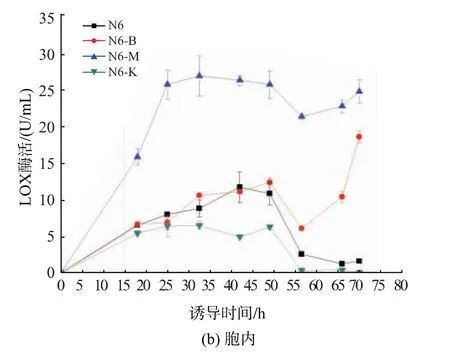
圖2 重組菌發酵產LOXFig.2 Fermentation production of the LOX by the recombinant strains
為進一步分析LOX 在重組菌N6-B、N6-M 和N6-K 中的表達情況,對其胞內可溶物質和發酵液上清液進行SDS-PAGE 分析。如圖3 所示,誘導表達32 h 和70 h 后,N6、N6-B、N6-M 和N6-K 胞內均存在70 kDa 條帶,與LOX 的理論分子量一致[1]。其中,N6-M 的LOX 條帶明顯較對N6 相應條帶增粗,與酶活測定結果一致(圖3)。此外,在胞內樣品中 未 見SodB(21 kDa)、SodM(22 kDa)和CAT(84 kDa)蛋白條帶,可能這幾種酶表達量不高或者表達后被快速降解。

圖3 SDS-PAGE 電泳分析LOX 與抗氧化酶的共表達Fig.3 SDS-PAGE analysis of the co-expression of LOX and antioxidase
2.3 重組菌胞內抗氧化酶活分析
為了分析重組菌中抗氧化酶的表達情況,測定了破碎后上清液檢測SOD 和CAT 活性。如圖4(a)所示,N6-B 和N6-M 胞內SOD 酶活波動較大,但總體上N6-M 中SOD 的酶活明顯高于N6-B 和N6;N6-B 中SOD 酶活總體水平較N6 未見明顯提高,但其酶活較N6 出現更早。研究表明,SOD 的表達受環境中K+和Ca2+影響較大[3],整個發酵周期中伴隨著大腸桿菌的生長和裂解,環境中的離子濃度會隨菌體變化產生一些波動可能引起了N6-B 和N6-M中SOD 酶活波動。因此,SOD 胞內活性的增加可能是LOX 大量累積的重要原因.由圖4(b)可知,在誘導前期N6-K 中CAT 酶活顯著提高,表明CAT 在N6 中也得到了活性表達。但是,N6-K 與N6 中胞內LOX 酶活未見明顯差異,胞外酶活有所下降。這說明CAT 活性未對LOX 的表達產生明顯促進作用,可能表達CAT 加重重組菌株表達外源蛋白質的負擔[9]。

圖4 重組菌胞內抗氧化酶活Fig.4 Antioxidase activity in recombinant strains
2.4 誘導條件的優化
重組蛋白的表達水平在一定范圍內與工程菌的生長具有一定的關聯性,其依賴于培養溫度、培養基組成、IPTG 的濃度和誘導時間等因素[9]。因此,確定重組菌株的誘導條件對LOX 的最終產量和工業化生產具有重要意義。根據第一步的實驗結果,選擇LOX 酶活力最高的N6-M 重組菌株作為優化對象,由于N6-M 中的兩個質粒均為T7 啟動子,同時IPTG 存在著潛在的毒性,所以選取誘導劑濃度和誘導時菌體濃度兩個因素。由于誘導30 h 胞內LOX 已經大量積累,所以檢測誘導30 h 細胞破碎后上清中LOX 酶活作為實驗的判斷標準。正交試驗結果如表3 所示。
分析正交試驗得出以下結論:1)由極差大小可以直觀判斷出各因素對LOX 酶活的影響主次順序為:誘導時菌體濃度(A)>IPTG 終濃度(B)。蛋白的表達依賴于足夠量和生長旺盛的菌體,所以重組菌的生長和蛋白的表達在時間上存在差異[8]。重組菌的數量不足,即菌株處于對數期前期及對數期前,能夠表達重組蛋白的菌體不足,重組蛋白的表達量會受到較大影響。2)N6-M 發酵產LOX 的因素最佳條件組合為:誘導時菌體濃度(OD600)2.5+IPTG 終濃度2.0 mmol/L(即A3B4),胞內LOX 酶活為28.8 U/mL,較優化前(誘導OD600=0.6,IPTG=1 mmol/L)提高5.88%。

表3 正交實驗結果Table 3 Results of orthogonal experiment
3 討論
作為典型的過氧化物酶,LOX 在工業生產中的應用受到人們的日益關注。LOX 可以對面粉進行漂白,改善面粉筋力,改善面團的水合力,提高面團強度和增加食物風味,大豆提取的LOX 各批次之間會有較大差異,因此利用大腸桿菌表達LOX 是工業化生產穩定的LOX 的重要基礎。來源于動物,植物等真核生物的LOX 基因在大腸桿菌[10]、酵母、曲霉等系統中實現了表達。原核LOX 基因研究較少,僅Cyanobacterium、N.punctiforme 及P.aeruginosa 在大腸桿菌系統中得到表達[11]。酵母系統中研究者利用信號肽使LOX 分泌到胞外,簡化了下游操作,但產量十分有限;枯草芽孢桿菌中LOX 胞外產量僅有0.01 U/mL[12-13],Steczko 在E.coli 表達系中實現了大豆LOX 的胞內表達,酶活達到4.5 U/mL[14-15]。但總體而言,LOX 的異源表達的水平偏低。
氧毒性在各種物種中均有發現,是由分子氧的一價還原產物介導的,包括超氧自由基(O2-),過氧化氫(H2O2)和羥基(OH-)[3]。氧毒性對生物和非生物的威脅可能來自于O2-內在和相對選擇性的反應性,而且O2-可以通過質子化得到氧化性更強的HO2-,或者通過鐵催化的Haber-Weiss 反應生成OH·。而LOX 能催化內源性化學物如花生四烯酸的反應并伴有活性氧的產生,LOX 還能通過影響細胞間的信號轉導而激活其他活性氧產生途徑;此外,LOX 還能催化外源化學物代謝為高活性的自由基中間體而誘發活性氧的產生。因此,如何降低LOX活性對宿主菌的毒性可能是其高效表達的關鍵。
SOD 是非常高效的催化劑,25 ℃下速度約為2×109m/s,活化能為26.9 kJ/M[16]。N6 表達的LOX 為銅綠假單胞菌(P.aeruginosa BBE)來源,而P.aeruginosa 中會產生幾種SOD 包括SodB 和SodM,與降解胞內活性氧中間體有關[17]。因此,本研究選擇P.aeruginosa 來源的SodB,SodM 和E.coli 來源的CAT 三種抗氧化酶,構建共表達載體,降低N6 中LOX 大量合成對細胞產生的壓力,從實驗結果來看,共表達SodM 時,N6-M 中LOX 酶活提高最多,是原始N6 的2.4 倍(圖2)。雖然影響外源蛋白在宿主中表達因素還有很多,但本研究為LOX 的異源表達提供了一個方向,降低胞內活性氧中間體可能是一種解決方法。
4 結語
微生物發酵產品想要成功實現產業化,發酵工藝的優化是重要的環節。本研究通過正交實驗對N6-M 發酵的菌體濃度和IPTG 濃度兩個條件進行優化,實驗結果表明OD600為2.5 時,添加終濃度為2 mmol/mL 的IPTG 誘導30 h 后胞 內LOX 酶活達到28.8 U/mL,與OD6000.6,IPTG 終濃度1 mmol/mL的初始條件相比提高了5.88%。
