逆轉錄病毒介導的穩定表達E4orf1和綠色熒光蛋白的胎肝竇內皮細胞株的建立
2018-08-29 01:01:40李慧琳謝小燕王思涵韓毅范增白云何麗娟南雪裴海云3岳文裴雪濤
生物技術通訊 2018年4期
關鍵詞:血清
李慧琳,謝小燕,王思涵,韓毅,范增,白云,何麗娟,南雪,裴海云3,,岳文,裴雪濤,
1.南方醫科大學 檢驗與生物技術學院,廣東 廣州 510515;2.軍事科學院 軍事醫學研究院 衛生勤務與血液研究所軍事再生醫學研究室,北京 100850;3.軍事科學院 軍事醫學研究院 輻射醫學研究所實驗血液學與生物化學研究室,北京 100850;4.華南生物醫藥研究院 華南干細胞與再生醫學研究中心,廣東 廣州 510005
造血干細胞(hematopoietic stem cells,HSCs)具有高度自我更新能力及多向分化潛能,可以產生所有類型的血液細胞,如紅細胞、白細胞、血小板和淋巴細胞等[1]。它在生命過程中不僅能夠重建整個造血系統,還具備維持長期造血的功能。近年來,HSCs移植越來越多地用于臨床治療血液或非血液系統的惡性腫瘤,顯示出廣闊前景[2]。其中,尤以臍帶血移植(umbilical cord blood transplantation,UCBT)備受青睞,它具有來源廣泛、易于采集、對供體無傷害、HLA配型要求低、移植后復發率及移植物抗宿主病(graft-versushost disease,GVHD)發病率較低等優點。然而單份臍帶血中HSCs絕對數量較少,輸注后易導致中性粒細胞恢復延遲并增加細菌及病毒感染的風險[3],而雙份臍帶血移植則會帶來GVHD發病率增加、血小板恢復時間延長等一系列問題[4]。綜合來看,對HSCs進行體外擴增培養是最直接便捷的解決方法,但迄今構建適宜的HSCs體外擴增微環境仍是亟待解決的瓶頸問題。
造血微環境由臨近HSCs的多種支持細胞構成,包含成骨細胞、內皮細胞、脂肪細胞、基質細胞和免疫細胞等[5],其中內皮細胞與HSCs擁有共祖細胞生血內皮、共表達許多表面標志和轉錄因子如CD34、CD31、Runx1、GATA-2等,而且可以通過分泌多種因子影響HSCs的增殖和自我更新,成為造血微環境的核心組分[6]。在以內皮細胞為支持細胞培養HSCs的研究中,有報道表明血清和促血管生成因子可以抑制內皮細胞的自分泌對造血穩態的維持作用。為保留內皮細胞特性并為與HSCs的共培養提供基本條件,Seandel等將腺病毒E4區開放讀框1(E4orf1)的基因轉入原代臍靜脈內皮細胞,通過活化Akt,使其獲得永生能力且可以維持內皮自身特性,成為研究血管微環境的適宜模型[7]。
在胚胎發育中,肝臟是一個重要的造血器官,也是HSCs自我更新與擴增最旺盛的場所,其中的胎肝竇內皮細胞(human fetal liver sinusoid endothelial cells,HFLSECs)是一類結構獨特的血竇內皮,約占肝非實質細胞總數的70%,具有吞噬、抗原提呈、血流調節等功能[8],可以分泌內皮素1(ET-1)、一氧化氮(NO)、血管內皮生長因子(VEGF)、轉化生長因子β1(TGF-β1)、腫瘤壞死因子(TNF)等多種細胞因子,還參與髓外造血以及HSCs在肝小葉區的選擇性植入[9]。這提示我們,這類肝臟所特有的內皮細胞有可能是影響HSCs維持與自我更新的重要微環境因素。在本研究中,我們利用逆轉錄病毒載體系統建立了穩定表達E4orf1和綠色熒光蛋白(green fluorescent protein,GFP)的 HFLSECs(E4orf1-GFP/HFLSECs),期望以該轉基因的內皮細胞作為飼養層,構建促進HSCs體外擴增的新培養體系,也為多潛能干細胞向HSCs分化或直接重編程獲得HSCs提供維持增殖與自我更新的微環境。
1 材料與方法
1.1 材料
HFLSECs購自Pricells公司;臍帶血由海軍總醫院和北京中西醫結合醫院婦產科提供;感受態大腸桿菌DH5α購自康為世紀公司;質粒MSCVN E4orf1購自Addgene公司;攜帶GFP基因的逆轉錄病毒載體pMX-GFP為美國Hiroyuki Hirai教授惠贈。
LipofectAMINE 2000和Opti-MEM培養基購自Invitrogen公司;多聚-D-賴氨酸氫溴酸鹽(PDL)、明膠、牛血清白蛋白(BSA)購自Sigma公司;高糖DMEM培養基、胎牛血清(FBS)、0.25%胰蛋白酶-EDTA購自Gibco公司;EGM-2 Bullet kit購自Lonza公司;殺稻瘟菌素(blasticidin)、嘌呤霉素(puromycin)購自Invivogen公司;聚凝胺(polybrene)購 自 Millipore公 司 ;CD144-PE、KDR、CD133-PE、CD45-APC抗人單克隆抗體購自BD公司;CD117-APC、CD31-APC抗人單克隆抗體購自Ebioscience公司;兔抗人假性血管血友病因子(von Willebrand Factor,vWF)單克隆抗體購自Sino Biological公司;羊抗兔熒光二抗購自中杉金橋公司;質粒提取試劑盒購自Axygen、QIAGEN公司;紅細胞沉降液、人外周血淋巴細胞分離液購自TBD公司;CD34分選磁珠購自美天旎公司;StemSpan培養基、造血集落培養基MethoCult H4434購自Stemcell公司;干細胞因子(stem cell factor,SCF)、Fms樣酪氨酸激酶3配體(Fms-like tyrosine kinase 3 ligand,Flt-3L)、促血小板生成素(thrombopoietin,TPO)購自Peprotech公司。
1.2 免疫熒光
吸棄培養HFLSECs孔板中的培養上清,用PBS清洗3次后,4%多聚甲醛室溫固定細胞20 min,PBS-T(PBS+0.1%吐溫20)清洗2次,以PBST(含1%BSA+1%吐溫20)室溫孵育10 min進行破膜,再用PBS-T清洗3次,每次5 min,用PBS-T(含1%BSA)室溫孵育30 min封閉后,加入兔抗人vWF抗體(1∶1000)4℃孵育過夜,次日棄上清,用PBS-T清洗3次,每次5 min,再加入羊抗兔FITC IgG抗體(1∶500)避光室溫孵育1 h,棄上清,PBS-T洗3次,每次5 min,最后加入DAPI(1∶500)室溫復染細胞核1 min,棄上清,PBS洗3次,每次5 min,將細胞培養板置于Nikon ECLIPSE Ti型倒置熒光顯微鏡下鏡檢,拍照。
1.3 質粒轉化、提取、酶切鑒定及測序
取50 μL感受態大腸桿菌DH5α加入1~3 μL質粒 MSCV-N E4orf1,冰浴 30 min,42℃熱激 60 s,再冰浴5 min;向細菌-DNA復合物中加入500 μL LB培養基,于37℃搖床上300 r/min培養45 min后8000 r/min離心3 min,取20~80 μL菌液涂布于含氨芐青霉素(100 μg/mL)的LB固體培養基上,37℃倒置培養過夜;次日挑取單個菌落加入5 mL含氨芐青霉素(100 μg/mL)的 LB培養基中,37℃搖床上200 r/min培養16 h,將菌液以1∶200的比例加入500 mL含氨芐青霉素(100 μg/mL)的LB培養基中,37℃搖床上200 r/min培養16 h;用質粒提取試劑盒提取質粒MSCV-N E4orf1,用限制性內切酶XhoⅠ和EcoRⅠ酶切鑒定,送Sangon Biotech公司測序。
1.4 嘌呤霉素條件培養殺滅實驗
用0.25%胰酶消化 HFLSECs,以1×105/孔接入24孔板培養,16~24 h后替換為分別含0.5、1、2、4 μg/mL嘌呤霉素的EGM-2培養基,最后一孔仍然用不含嘌呤霉素的EGM-2培養基培養,作為對照組,在顯微鏡下觀察細胞存活情況。
1.5 逆轉錄病毒包裝、細胞轉染、藥物篩選及轉染后HFLSECs流式細胞分選(fluorescence-activated cell sorting,FACS)
復蘇包裝細胞Plat-A,接種至預先包被0.1%明膠的培養瓶中,用 DMEM(含 10%FBS、1 μg/mL殺稻瘟菌素、0.1 μg/mL嘌呤霉素)培養,當細胞生長至70%~80%融合時,用0.25%胰酶消化Plat-A,以5×105/孔接入預先包被PDL的6孔板中用1.5 mL DMEM(含10%FBS)培養,包裝所用培養基不含抗生素,24 h后,用LipofectAMINE 2000介導質粒MSCV-N E4orf1和pMX-GFP轉染,將5 μg質粒及10 μL脂質體加入500 μL無血清Opti-MEM中,室溫靜置20 min后,將含有DNA-LipofectAMINE 2000復合物的無血清培養基加入Plat-A中,培養20 h后替換EGM-2培養基1.5 mL/孔,同時復蘇HFLSECs,以1×105/孔接入 6 孔板中用EGM-2培養,24 h后收集病毒上清,將用0.45 μm無菌濾膜過濾器過濾后的病毒液(含8~10 μg/mL聚凝胺)直接加入用PBS洗滌1次的HFLSECs中,Plat-A加入EGM-2繼續培養24 h,用同樣方法收取第二次病毒加入HFLSECs中,培養24 h后換液,1~2 d后加壓藥篩,之后隔天換液,當細胞培養至90%匯合時用0.25%胰酶消化,收集細胞進行流式分選。
1.6 流式細胞術
培養E4orf1-GFP/HFLSECs至細胞長至80%~90%匯合,PBS洗滌細胞1次,用0.25%胰酶于37℃消化細胞3~5 min,終止消化后收集細胞于15 mL離心管中,室溫1000 r/min離心5 min,棄上清,PBS重懸細胞后分裝于1.5 mL EP管中,每管50~100 μL,分別標記流式抗體,避光4℃旋轉孵育 40 min~1 h,用 PBS 洗滌 2 次,用 200 μL PBS重懸細胞,篩網過濾后收集細胞于流式管中,1 h內上BD FACSAria型流式細胞儀進行檢測。
1.7 分離人臍帶血(umbilical cord blood,UCB)CD34+細胞
將臍帶血轉入無菌T75細胞培養瓶中,按總體積的三分之一加入紅細胞沉降液,充分混勻,自然沉降20~40 min,用滴管收集上層血漿層至50 mL離心管中,室溫2000 r/min離心5 min,10 mL PBS洗滌1次,用5 mL PBS重懸細胞;用密度梯度離心法獲得單個核細胞(mononuclear cells,MNCs),在15 mL離心管中加入5 mL 外周血淋巴細胞分離液,用滴管將5 mL細胞懸液在分離液液面上方1 cm處沿離心管壁緩慢加至分離液液面之上,將離心管置于水平離心機中,以最低檔升降速室溫2000 r/min離心20 min,離心后管內細胞懸液分為4層,用滴管吸取灰白色云霧狀細胞層置于50 mL離心管中,加入PBS洗滌1次,重懸細胞至1.5 mL EP管中,按照CD34 Microbeads kit說明書加入CD34分選磁珠,4℃避光旋轉孵育40 min,PBS洗滌1次,于磁場中進行CD34+細胞分選。
1.8 E4orf1-GFP/HFLSECs作為飼養層培養人UCB來源的CD34+細胞
從UCB中分離獲得CD34+細胞,以5×104/孔接入E4orf1-GFP/HFLSECs中,以無飼養層的懸浮培養作為對照組,以StemSpan培養基(含SCF、Flt-3L、TPO各50 ng/mL)連續培養15 d,每隔3 d計數有核細胞(nucleated cells,NC)總數,當細胞總數超過1×106時,須將細胞傳代至新的飼養層上繼續培養。
1.9 集落形成實驗
將擴增后的HSPCs以250/孔接種于造血集落培養基MethoCult H4434中連續培養14 d,觀察并拍照記錄不同集落形成單位(colony forming units,CFU),用血球計數板和臺盼藍染色進行細胞計數。
1.10 數據統計分析
數據以x±s表示,用Excel進行統計分析。
2 結果
2.1 免疫熒光鑒定vWF在HFLSECs中的表達
vWF也稱Ⅷ因子相關抗原,是一種由血管內皮細胞合成并分泌的大分子蛋白多聚體,在體內參與血液凝固和血栓形成[10],常被作為特征因子來鑒定體外培養的內皮細胞。免疫熒光結果(圖1)表明,原代培養的HFLSECs的vWF表達陽性。
2.2 質粒MSCV-N E4orf1的酶切鑒定及測序
提取質粒后用XhoⅠ和EcoRⅠ雙酶切,獲得約530 bp的E4orf1編碼序列片段(圖2)。此外,基因測序結果進一步證實與GenBank報道的序列一致(序列略)。
2.3 篩選穩定轉染MSCV-N E4orf1的HFLSECs的最佳藥物濃度
在含0.5 μg/mL嘌呤霉素的培養條件下,原本貼壁生長的原代培養的HFLSECs從第2 d起開始逐漸變圓,第3 d起逐漸漂浮到培養基中(圖3),5 d內基本觀察不到貼壁生長的細胞,而在1、2、4 μg/mL嘌呤霉素培養條件下,原代培養的HFLSECs第2 d便全部漂浮,藥物作用過強,因此,選擇0.5 μg/mL嘌呤霉素為最適藥篩濃度。

圖1 原代培養的HFLSECs高表達vWF

圖2 質粒MSCV-N E4orf1的酶切鑒定

圖3 嘌呤霉素的最佳藥物濃度篩選
2.4 逆轉錄病毒包裝
包裝20 h后即可見Plat-A細胞內有GFP表達,表達率超過90%(圖4)。

圖4 Plat-A細胞的GFP表達
2.5 E4orf1-GFP/HFLSECs可在無血清環境中存活
分別包裝質粒MSCV-N E4orf1和pMX-GFP為逆轉錄病毒,收集毒液按1∶1的比例同時轉染HFLSECs,并采用0.5 μg/mL嘌呤霉素加壓篩選1周,再進行流式細胞分析,其GFP陽性率為90.5%,通過流式分選獲得GFP+細胞(圖5),分選后的E4orf1-GFP/HFLSECs能在含0.5 μg/mL嘌呤霉素的培養基中穩定擴增傳代(圖6),將其轉入無血清的EGM-2中進行培養,仍能維持存活且并不增殖(圖7)。
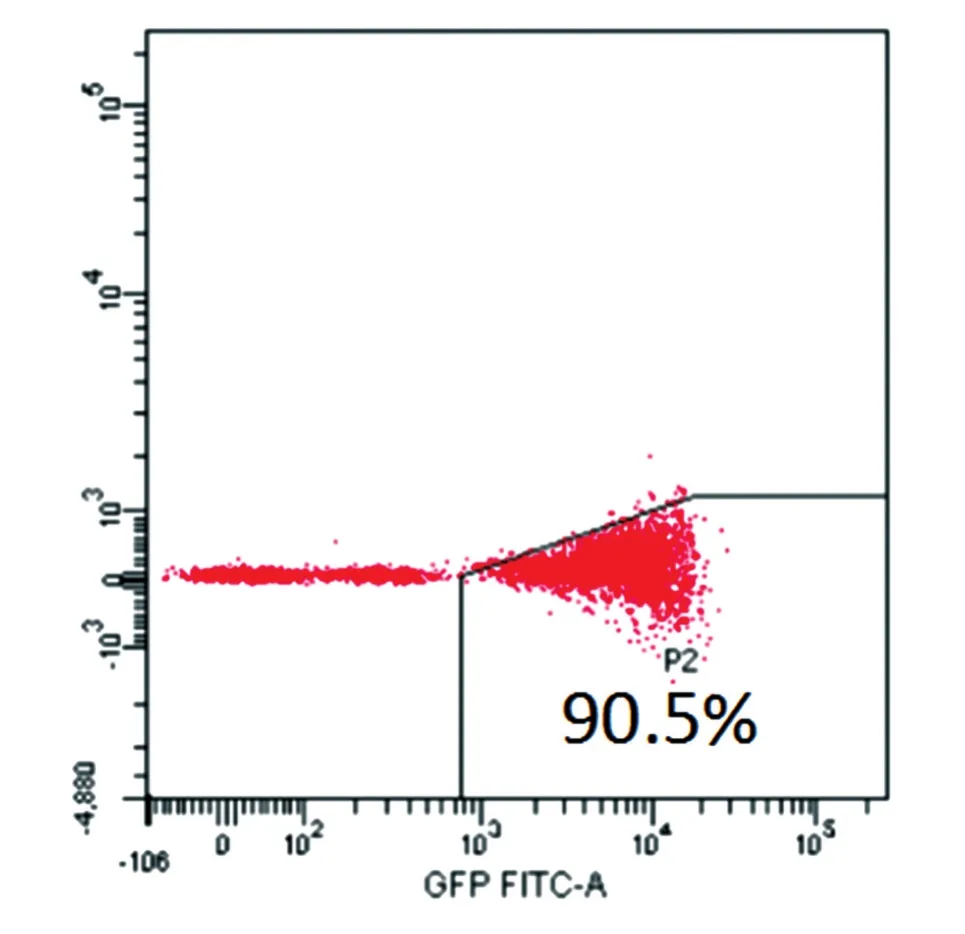
圖5 流式分選中E4orf1-GFP/HFLSECs的GFP表達率
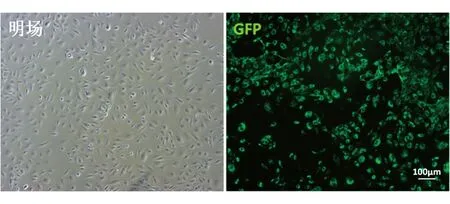
圖6 分選后E4orf1-GFP/HFLSECs的GFP表達
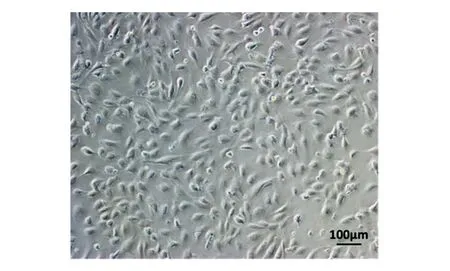
圖7 無血清環境下的E4orf1-GFP/HFLSECs
2.6 流式細胞術檢測E4orf1-GFP/HFLSECs的表面標志
流式細胞檢測結果(圖8)顯示,E4orf1-GFP/HFLSECs的內皮細胞表面標志CD144和CD31的陽性率分別為99%和99.5%,而干細胞表面標志CD117、內皮祖細胞表面標志CD133及血細胞表面標志CD45則幾乎不表達,血管內皮生長因子受體KDR表達率為69%。
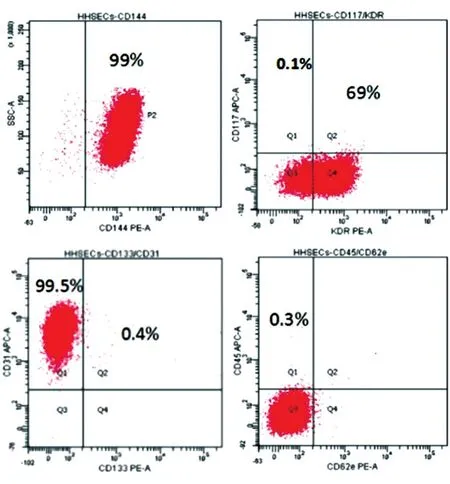
圖8 流式分析E4orf1-GFP/HFLSECs的表面標志
2.7 E4orf1-GFP/HFLSECs作為飼養層支持人臍帶血來源的CD34+細胞的體外擴增
每3 d計數各組中懸浮的有核細胞總數,結果表明,在E4orf1-GFP/HFLSECs作為飼養層的體外擴增體系中,人臍帶血來源的CD34+細胞15 d有核細胞總數擴增了360倍,而作為對照的單純因子懸浮培養組僅擴增了165倍,實驗組的擴增效率是對照組的2.2倍(圖9)。
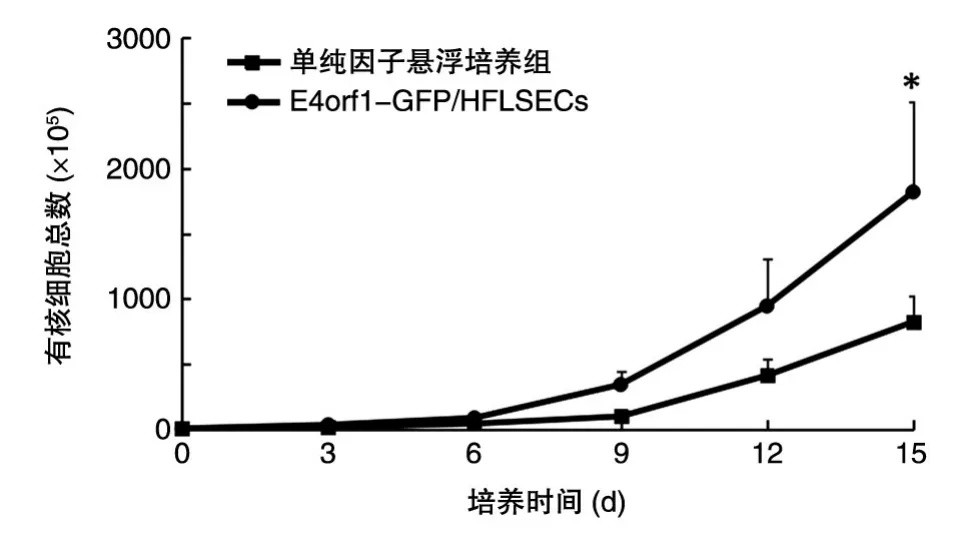
圖9 E4orf1-GFP/HFLSECs促進HSPCs的體外擴增*P<0.05
2.8 人臍帶血來源的CD34+細胞經過體外擴增后仍具有造血集落形成能力
在以E4orf1-GFP/HFLSECs作為飼養層的體外擴增體系中,人臍帶血來源的CD34+細胞經過擴增后在體外仍具有分化為不同CFU的能力(圖10),包括紅系爆式集落形成單位(BFU-E)、紅系集落形成單位(CFU-E)、粒系集落形成單位(CFU-G)、巨核系集落形成單位(CFU-M)、粒系和巨噬系集落形成單位(CFU-GM)及紅系、粒系、巨核系、巨噬系混合集落形成單位(CFUEGMM)。
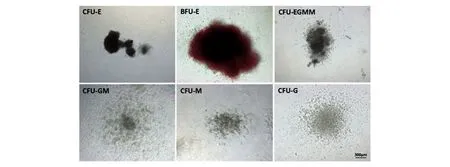
圖10 擴增后細胞在體外仍具有多系分化能力
3 討論
1978年Schofield首次提出了干細胞龕(niche)的概念,指出特異性的微環境會對干細胞功能產生作用,隨后有大量研究證實了多種組織中干細胞龕的存在[11-13]。目前,維持HSCs自我更新的造血微環境至少包括3個解剖區域(骨內膜、血竇壁和固有造血組織)和多種細胞類型(成骨細胞、血竇內皮細胞、脂肪細胞、基質細胞和免疫細胞等)。人們曾嘗試多種方式來模擬體內造血微環境,以期望構建維持HSCs自我更新與擴增的體外培養體系,如造血相關的細胞因子、小分子,基質細胞共培養,Notch配體等[14-18],但效果仍然差強人意。
胎肝是胚胎發育時期HSCs迅速擴增的重要場所,以小鼠為例,當造血中心從背主動脈和胎盤遷移至胎肝后,僅僅4 d,HSCs在其中完成近40倍的擴增。已有研究證實[19-20],肝血竇是肝臟內造血集落的形成場所,而其中的肝竇內皮細胞對于髓外造血以及HSCs在肝小葉區的選擇性植入具有關鍵作用,是重要的肝內造血微環境因素之一。有鑒于此,我們擬建立胎肝竇內皮細胞飼養層作為造血微環境平臺。考慮到血清會影響血管的功能,干擾造血干祖細胞的擴增,所以與HSCs共培養時實現飼養層細胞的無血清存活是至關重要的。而原代內皮細胞在無血清、無血管內皮相關因子(如VEGF、FGF、IGF、EGF等)的條件下會喪失功能并迅速凋亡。已有文獻揭示腺病毒的E4orf1在激活存活通路的同時,并不會促進內皮細胞的擴增和轉化,可以作為生存蛋白,使內皮細胞即便在無血清的條件下仍然存活、維持內皮細胞的特性且不爭奪與之共培養細胞的營養[7],因此我們采用逆轉錄病毒系統并利用載體上的嘌呤霉素抗性基因篩選獲得穩定轉染E4orf1的HFLSECs。此外,為了便于日后應用于多潛能干細胞向HSCs分化或直接重編程獲得HSCs時與起始細胞進行區分,我們共轉染了表達GFP的逆轉錄病毒載體,并通過流式分選進行純化。實驗結果顯示,該細胞的組織特異性在轉染前后并未發生轉變,仍能保留原代HFLSECs的內皮特性如vWF的表達,且能在無血清培養狀態下正常存活,適宜作為HSCs體外擴增培養體系中的飼養層細胞。
與以往報道的以飼養層細胞提供微環境促進HSCs體外擴增方法[21]相比,E4orf1-GFP/HFLSECs無須經過絲裂霉素C或放射線的預處理,而且能在無血清培養狀態下維持內皮細胞的特性,可以更好地為HSCs提供體外擴增微環境。初步實驗結果表明,在E4orf1-GFP/HFLSECs的支持下,人臍帶血來源的CD34+細胞在連續培養15 d后有核細胞總數擴增了360倍,而最近報道的胎肝細胞[22]或內皮祖細胞[23]僅僅實現了短期(7 d)擴增,有核細胞總數擴增效率分別為8倍和130倍。
綜上所述,我們建立了能夠在無血清、無內皮細胞相關因子條件下存活的內皮微環境平臺,以此作為飼養層細胞可以為HSCs體外擴增、多潛能干細胞向HSCs分化或直接重編程獲得HSCs帶來新的研究思路和方法。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
