燈盞乙素對阿爾茨海默病小鼠腦組織β分泌酶途徑相關蛋白表達的影響
2018-07-27 03:35:08王豫君敖俊文郭莉莉于燕妮
山東醫藥 2018年26期
王豫君,敖俊文,郭莉莉,于燕妮
(1貴州醫科大學,貴陽550004;2貴陽市第一人民醫院)
阿爾茨海默病(AD)是一種中樞神經系統變性疾病,主要表現為漸進性記憶障礙、認知功能障礙、人格改變及語言障礙等神經精神癥狀,嚴重影響患者的生活和社交功能。β淀粉樣肽(Aβ)在AD的發病過程中起重要作用,延緩或阻止Aβ沉積所致的神經元機能衰退及神經毒性是治療AD的研究方向之一[1]。β分泌酶途徑是Aβ病理性沉積中β淀粉樣前體蛋白(APP)代謝的主要途徑, 分泌酶作為Aβ產生的重要限速因子,能夠特異靶向膜結合底物,啟動Aβ的生成[2]。燈盞乙素(Scu)是燈盞細辛的藥理活性成分。研究顯示,Scu能夠減輕癡呆大鼠神經型尼古丁受體蛋白表達和膽堿酯酶活性異常所致的膽堿能系統功能損害[3];消除自由基的產生,提高機體抗氧化能力[4];抑制炎性因子釋放、減輕神經炎癥損傷[5];抑制神經細胞凋亡,對抗Aβ的毒性[6]。2015~2017年,我們將類AD動物模型APPswe/PS1dE9雙轉基因小鼠做為研究對象,觀察Scu對β分泌酶的裂解活性物質β淀粉樣前體蛋白裂解酶1(BACE1)、BACE2以及總APP表達的影響,探討藥物干預APP代謝的β分泌酶途徑來抑制Aβ生成的作用機制。
1 材料與方法
1.1 材料 3月齡APPswe/PS1dE9雙轉基因陽性小鼠40只,3月齡背景品系C57BL/6J小鼠10只,雌雄各半,由中國醫學科學院實驗動物所遺傳中心提供,動物實驗經貴陽市第一人民醫院動物倫理委員會批準。Scu由南京澤朗生物醫藥科技公司提供,高效液相色譜(HPLC)法鑒定純度大于98%。多克隆抗BACE1抗體(美國GENETEX公司);抗β-actin抗體(美國CST公司);多克隆抗APP、BACE2抗體(美國Abcam公司);SYBR Green PCR 試劑盒(美國KAPA Biosystems公司);逆轉錄試劑盒(日本TAKARA公司);APP、BACE1、BACE2和內參照β-actin引物由南京金斯瑞生物科技有限公司合成購得。實時熒光定量 PCR 儀(美國BIO-RAD公司);MK30酶標儀(芬蘭雷柏公司);Centrifuge 5424 R離心機(德國Eppendorf公司)。
1.2 動物分組與處理 將APPswe/PS1dE9小鼠隨機分為模型組和Scu低、中、高劑量組各10只,C57BL/6J小鼠作為正常對照組。小鼠進入SPF級實驗室,分籠分性別飼養,每籠5只,普通繁殖飼料飼養。藥物劑量根據患者臨床用量的等效劑量和動物體表面積折算所得,Scu低、中、高劑量組分別給予Scu 1.5、2.5、4.0 mg/(100 g·d)灌胃,正常對照組和模型組給予等量生理鹽水灌胃,每日上午9點給藥,連續90 d。
1.3 學習記憶能力測試 采用Morris水迷宮實驗。治療結束后第1~4天進行定向航行實驗,記錄60 s內小鼠從不同象限入水到爬上平臺(固定于第1象限)所需時間,即逃避潛伏期。每天連續測試4次,取平均值。第5天撤去平臺,進行空間探索實驗,記錄小鼠第1次穿越原站臺時間及1 min內穿過原站臺位置的次數。
1.4 標本采集 Morris水迷宮實驗完成后,將小鼠斷頸處死,剝離腦組織,置于-80 ℃凍存,用于APP、BACE1、BACE2蛋白及mRNA檢測。
1.5 腦組織中APP、BACE1、BACE2蛋白表達檢測 采用Western blotting法。取各組腦組織,加入蛋白裂解液,冰上勻漿,4 ℃ 12 000 r/min離心15 min,取上清進行蛋白質定量。SDS-聚丙烯酰胺凝膠電泳分離蛋白,將蛋白質轉移至PVDF膜上,加入8%脫脂奶粉封閉。分別與一抗(APP、BCAE1、β-actin按1∶1 000稀釋,BACE2按1∶500稀釋)和二抗(按1∶10 000稀釋)孵育后曝光顯影,定影。用Image J軟件對膠片進行灰度掃描得出條帶像素灰度值。以β-actin蛋白條帶作為內參照,計算目的蛋白與β-actin蛋白條帶像素灰度的比值作為目的蛋白的相對表達量。
1.6 腦組織中APP、BACE1、BACE2 mRNA表達檢測 采用Real-time PCR法。用TRIzol試劑提取腦組織總RNA,反轉錄合成cDNA。以cDNA為模板,β-actin為內參進行擴增。引物序列:APP上游5′-ACTGCCAAGAGGTCTA-3′,下游5′-GGTAAGGAATCACGAT-3′;BACE1上游5′-ATGGCTTTTGGCTAG-3′,下游5′-CGGAAGGACTGATTG-3′;BACE2上游5′-CTGGTGATTGGTGCG-3′,下游5′-TTCCGTGGAAAAGGG-3′;β-actin上游5′-CCCATCTACGAGGGCTAT-3′,下游5′-TGTCACGCACGATTTCC-3′。擴增體系:總體積20 μL,其中SYBR Green Mix 10 μL,上、下游引物各0.5 μL,cDNA模板1 μL,ddH2O 8 μL。反應條件:95 ℃ 3 min,95 ℃ 5 s,56 ℃ 10 s,72 ℃ 25 s,共39個循環。應用2-ΔΔCt法計算目的基因的相對表達量。
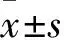
2 結果
2.1 各組學習記憶能力比較 見表1、2。與正常對照組相比,模型組在定向航行中逃避潛伏期時間延長,第1次穿越平臺時間明顯延長,平臺穿越次數減少(P均<0.01);與模型組比較,Scu高劑量組逃避潛伏期時間縮短,第1次穿越平臺時間縮短,平臺穿越次數增多(P均<0.01)。Scu中、低劑量組與模型組比較無統計學差異(P均>0.05)。

表1 各組定向航行實驗中逃避潛伏期時間比較
注:與正常對照組比較,*P<0.05;與模型組比較,#P<0.05。
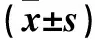
表2 各組第1次穿越平臺時間和平臺穿越次數比較
注:與正常對照組比較,*P<0.05;與模型組比較,#P<0.05。
2.2 各組腦組織中APP、BACE1、BACE2蛋白表達比較 見表3。與正常對照組比較,模型組腦組織中APP、BACE1蛋白表達升高,BACE2蛋白表達下降(P均<0.05);與模型組比較,Scu高劑量組腦組織中APP、BACE1蛋白表達下降,BACE2蛋白表達升高(P均<0.05),Scu中、低劑量組各指標無明顯變化(P均>0.05)。
表3 各組腦組織中APP、BACE1、BACE2蛋白表達比較
注:與正常對照組比較,*P<0.05;與模型組比較,#P<0.05。
2.3 各組腦組織中APP、BACE1、BACE2 mRNA表達比較 見表4。與正常對照組比較,模型組腦組織中APP、BACE1 mRNA表達升高(P均<0.05);與模型組比較,Scu高劑量組腦組織中APP、BACE1 mRNA表達降低(P均<0.05),Scu中、低劑量組無明顯變化(P均>0.05)。各組BACE2 mRNA表達比較差異均無統計學意義(P均>0.05)。
表4 各組腦組織中APP、BACE1、BACE2 mRNA表達比較
注:與正常對照組比較,*P<0.05;與模型組比較,#P<0.05。
3 討論
Aβ沉積在相關腦區是AD的病理學特征之一。在AD發病機制的眾多學說中,“Aβ級聯學說”一直占主導地位[7]。該學說認為,Aβ的異常聚集可產生一系列神經毒性作用,誘發膽堿能功能障礙、神經炎性反應、氧化應激產生、線粒體功能障礙、突觸損害等,這些神經毒性反應相互作用形成環形事件,最終導致神經細胞凋亡,引發癡呆[8]。有研究認為,Aβ是AD的結果而不是起因,Aβ的生成是神經元損傷的非特異性反應,這種損傷導致了APP的表達增多,出現Aβ沉積[9,10]。研究證實,AD的病程與Aβ狀態、分布、性質的改變有關,Aβ寡聚體的毒性和Aβ沉積均可造成大腦結構的破壞;而且Aβ能夠在細胞間傳遞,而細胞內Aβ異常聚集與認知功能障礙直接相關[11,12]。因此尋找對抗Aβ沉積所致的學習記憶能力損害的藥物,在AD治療中具有重要意義。
Aβ由APP在代謝過程中產生。參與APP代謝的主要有α、β、γ分泌酶,α分泌酶主導的是非Aβ生成途徑,此途徑不產生完整的Aβ片段,被廣泛認為是有益的,能夠阻礙Aβ的形成;而β分泌酶主導的是病理條件下Aβ的生成,可形成大小不等的Aβ肽,以Aβ1-42的毒性最強。目前已知的β分泌酶主要包括BACE1及組織蛋白酶B、D、L等,其中BACE1主要分布于神經元細胞,在Aβ級聯學說中占重要地位,被視為AD防治藥物的重要靶點。BACE1所有的底物都是跨膜蛋白,APP為Ⅰ型跨膜蛋白,是BACE1的主要底物。BACE1在β位點處將APP肽段切開后產生一段β可溶性分泌型APP片段(sAPPβ)和一段含有完整Aβ的片段(C99),后者再經過γ分泌酶裂解得到相對分子質量約為4 kD的可溶性Aβ片段(Aβ1-38、Aβ1-40、Aβ1-42等),此途徑也被稱為Aβ生成的β分泌酶途徑。當BACE1過度表達時,細胞內的β分泌酶活性增強,Aβ分泌增多,加速神經元的損傷。目前,BACE1高表達和活性增強已成為診斷AD的一個重要生化指標[13]。研究表明,降低或敲除BACE1基因均可顯著減少Aβ的沉積[14]。BACE2具有與BACE1高度相似的氨基酸序列,同樣在β酶切位點對APP進行切割,不同的是,其主要分布于星型膠質細胞內,切割部位主要在Aβ區域內,不產生沉積的Aβ片段,不僅能降低腦組織中Aβ的沉積,還能對BACE1的切割作用進行競爭性的抑制,阻止Aβ的產生。值得一提的是,BACE2 mRNA在大多組織中都是低水平表達[15],腦組織中也是如此。
研究證實,APPswe/PS1dE9雙轉基因模型小鼠可在短時間內出現類AD的病理改變,腦組織中高表達APP和BACE1,Aβ過量產生,小鼠的學習記憶功能受損,并隨年齡增加,這些改變逐漸加重[16]。本研究結果顯示,與正常對照組比較,模型組腦組織中APP、BACE1表達在蛋白和mRNA水平均明顯升高;模型組BACE2蛋白表達下調,而BACE2 mRNA表達無明顯改變,這可能與其mRNA在腦組織中表達量處于極低水平有關,差異性不明顯可能由于該基因表達不在于轉錄水平的改變而在于翻譯后修飾。考慮到是長期給藥,實驗在藥物劑量設定上有所保留,中、低劑量組雖有改善趨勢,但未能顯示出統計學差異。
研究表明,Scu能夠對抗與Aβ毒性相關的癡呆大鼠學習記憶能力下降[17,18]。本研究著眼于Scu對Aβ生成途徑的干預,結果顯示,Scu高劑量組腦組織中APP、BACE1蛋白和mRNA水平較模型組降低,BACE2蛋白及mRNA水平較模型組升高,提示Scu可能通過降低β分泌酶的活性,減少BACE1對總APP的切割,在一定程度上阻止β分泌酶主導下的APP代謝,減輕Aβ沉積帶來的學習記憶損害。
綜上所述,Scu能夠調節AD模型小鼠Aβ生成途徑中主要因子APP、BACE1蛋白和mRNA以及BACE2蛋白表達,通過干預APP代謝的β分泌酶途徑來抑制Aβ生成。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
課堂內外·初中版(科學少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
國際放射醫學核醫學雜志(2021年10期)2021-02-28 08:41:58
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
