HIV-1感染對人CD4+T淋巴母細胞內ITAMs/ITIMs通路的影響
2018-06-22 07:38:00莫雨曉謝延崢劉京郭曉強何金洋廣州中醫藥大學熱帶醫學研究所廣州510405
山東醫藥 2018年21期
莫雨曉,謝延崢,劉京,郭曉強,何金洋(廣州中醫藥大學熱帶醫學研究所,廣州510405)
艾滋病(AIDS)是由人免疫缺陷病毒(HIV)引起的一種免疫系統缺陷疾病,是人類尚未攻克的致死性疾病之一。近年來,AIDS的發病率和病死率持續攀升,抗病毒治療仍然是主要的防治手段。但抗病毒藥物價格昂貴、不良反應大、患者服藥依從性差,因此對免疫信號通路的研究越來越受到重視[1]。研究表明,免疫激活和炎癥反應是AIDS進展的主要原因[2]。而機體的免疫和炎癥調控機制主要由包含免疫受體酪氨酸激活基序(ITAMs)和免疫受體酪氨酸抑制基序(ITIMs)的受體介導[3],并通過相關受體的磷酸化發揮作用[4]。包含ITIMs的免疫受體(SHP-1、SHP-2、SHIP-1、SHIP-2)起抑制免疫的負性調節作用,包含ITAMs的受體(Zap70、Syk)則主要起激活作用[5]。在信號傳導過程中,一方面,Src酪氨酸激酶使ITIMs的酪氨酸殘基磷酸化,繼而招募包含了SH2結構域的酪氨酸磷酸酶,從而使上游或下游分子去磷酸化,導致細胞激活過程中止[6];另一方面,Src酪氨酸激酶使ITAMs磷酸化,并為其提供一個SH2結構域的結合位點,用于結合Syk家族的非受體蛋白酪氨酸激酶,從而使免疫細胞激活[7]。蛋白酪氨酸磷酸化修飾調節大多數生物學過程,異常的蛋白酪氨酸磷酸化水平與多種人類疾病密切相關。有研究結果顯示,HIV-1感染者ITAMs/ITIMs相關受體的信號調控通路異常[7~9],但目前未見其具體機制的系統研究。因此,2017年9月~2018年3月,本研究以感染HIV-1的人CD4+T淋巴母細胞系(M8166細胞)為體外實驗模型,探討細胞內ITAMs/ITIMs信號通路的變化,為揭示AIDS的發病機制、篩選抗HIV藥物提供依據。
1 材料與方法
1.1 細胞、試劑及儀器 M8166細胞(即HIV-1的敏感細胞)、人類免疫缺陷病毒BK132 HIV-1(病毒載量為5×108copies/mL)均由美國Aaron Diamond AIDS研究中心惠贈。RPMI1640培養基、胎牛血清(Gibco公司),逆轉錄試劑盒(Thermo Scientific公司),定量聚合酶反應PCR試劑盒(ABI公司),TRIzol?Reagent(Ambion公司),各基因PCR引物由上海英駿生物有限公司合成,Western blotting及IP裂解液、SDS-PAGE 5X蛋白上樣緩沖液、BCA蛋白濃度測定試劑盒(碧云天公司),SHP-2抗體、SHIP1抗體、SHIP2抗體、Zap70抗體、Syk抗體、p-SHP-2抗體、p-SHIP1抗體、p-SHIP2抗體、p-Syk抗體、GAPDH抗體、辣根過氧化物酶標記山羊抗兔IgG(CST公司),SHP-1抗體、p-SHP-1抗體、HRP標記抗鼠IgG(santa公司),P-Zap70抗體(Abcam公司),ECL發光液(Milipore公司)。5804R冷凍離心機(德國Eppendorf公司),2720普通PCR擴增儀、ABI7300/7500熒光定量PCR儀(Invitrogen公司),1510多功能酶標儀(Thermo公司),165-8001垂直電泳儀(BioRad公司),C-DiGit化學發光成像儀(Li-Cor公司)。
1.2 M8166細胞培養 按1×107/mL將M8166細胞接種于細胞瓶中,于恒溫37 ℃、5% CO2飽和濕度條件下進行培養,3 d換液1次。培養基組成:RPMI1640培養基補充10%的滅活胎牛血清、丙酮酸鈉(終濃度為1 mmol/L)、L-谷氨酰胺(終濃度為2 mmol/L)、1%非必需氨基酸。
1.3 HIV-1感染 將對數生長期的M8166細胞接種于24孔板中,分別設置空白組和病毒組。空白組常規培養,不作特殊處理;病毒組以10 TCID50/mL的HIV-1進行感染,培養3 d后,收集細胞或提取蛋白。
1.4 M8166細胞內ITAMs/ITIMs通路相關受體基因表達檢測 采用RT-PCR法。提取細胞總RNA:收集細胞,加入TRIzol 500 μL以使其充分裂解;提核酸時加入氯仿0.1 mL,吹勻后靜置10 min,以4 ℃、13 000 r/min離心15 min,將無色上清液吸至新的無酶EP管中。加入異丙醇250 μL,顛倒混勻,13 000 r/min、4 ℃離心10 min,棄上清后加入75%的乙醇500 μL,再以13 000 r/min、4 ℃離心5 min,棄上清,再離心1 min,吸凈液體,管底白色沉淀即為所得細胞總RNA,用DEPC水10 μL溶解沉淀,吹勻。根據Thermo公司的反轉錄試劑盒說明書制作逆轉錄體系,然后置于PCR擴增儀進行逆轉錄。制作PCR反應體系25 μL,反應條件:95 ℃ 1 min;95 ℃ 15 s,60 ℃ 1 min,共40個循環。目的基因與內參基因的比值即為目的基因相對表達量。基因引物通過美國ABI Primer Express 3.0軟件設計。SHP-1產物片段為130 bp,上游引物序列為5′-GACAAGGAGAAGAGCAAGGGT-3′,下游引物序列為5′-GGCACTCCTAGGTTCAGGTTG-3′;SHP-2產物片段為125 bp,上游引物序列為5′-TTGTTCTTTCTGTGCGCACTG-3′,下游引物序列為5′-ATCAAACCGTTCTCCTCCACC-3′;SHIP-1產物片段為134 bp,上游引物序列為5′-GGAAGGAGCAGGAAATCAGCT-3′,下游引物序列為5′-GCTTAACTGTGGCTGGTGATG-3′;SHIP-2產物片段為150 bp,上游引物序列為5′-GTGCCTATTTATTGGGGATCTGC-3′,下游引物序列為5′-AACAGGACAGCACCAGAGTAAC-3′;Zap70產物片段為122 bp,上游引物序列為5′-CACCCGAATGCATCAACTTCC-3′,下游引物序列為5′-CTCCGGCCCTTTCATCTTCTT-3′;Syk產物片段為136 bp,上游引物序列為5′-CAAGGGGCTAGCTGGATTTGT-3′,下游引物序列為5′-GGCCCGATGAGTAACAAATGC-3′。
1.5 M8166細胞內ITAMs/ITIMs通路相關受體蛋白和磷酸化蛋白表達檢測 采用Western blotting法。將細胞收集至EP管中,以800 r/min、4 min離心,棄上清,用4 ℃預冷的PBS清洗3次以洗凈殘余的培養基,然后加入含蛋白酶抑制劑和磷酸酶抑制劑的RIPA裂解液400 μL,再以超聲破碎儀裂解細胞,12 000 r/min,4 ℃離心15 min,取上清,即為所得細胞蛋白。以BCA法進行蛋白定量,調平蛋白上樣濃度,同時以4∶1的比例加入5×上樣緩沖液,煮沸變性。每孔上樣總蛋白量為40 μg,經SDS-PAGE凝膠電泳(恒壓120 V,80 min)后,制備“三明治夾心”,進行轉膜(恒壓100 V,100 min),轉膜結束后,取下PVDF膜,用TBST洗膜5 min,以5%脫脂牛奶在搖床封閉1 h,封閉完成后用TBST洗膜3次,每次5 min,再根據各抗體說明書,摸索條件,最終以SHP-1(1∶200)、p-SHP-1(1∶200)、SHP-2(1∶1 000)、p-SHP-2(1∶1 000)、SHIP1(1∶1 000)、p-SHIP1(1∶1 000)、SHIP2(1∶1 000)、p-SHIP2(1∶1 000)、Syk(1∶1 000)、p-Syk(1∶1 000)、Zap70(1∶2 000)、p-Zap70(1∶500)、GAPDH(1∶1 000)一抗4 ℃孵育過夜。孵育一抗后,洗膜3次,每次5 min,洗去未結合一抗,然后二抗(1∶2 000)室溫孵育1 h,洗膜3次,每次5 min。將ECL發光液A、B底物以1∶1等量混勻后滴加于PVDF膜上,然后在化學發光成像儀中顯影。使用Image J軟件測定條帶灰度值。目的條帶灰度值/內參條帶灰度值即為目的蛋白相對表達量。

2 結果
2.1 HIV-1感染對M8166細胞內ITAMs/ITIMs通路相關基因表達的影響 與空白組比較,病毒組SHP-1 mRNA相對表達量高(P<0.01),SHIP1 mRNA相對表達量低,但差異無統計學意義(P>0.05);兩組SHP-2、SHIP2、Zap70、Syk mRNA相對表達量比較差異無統計學意義(P均>0.05)。見表1。
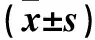
表1 兩組ITAMs/ITIMs通路相關基因相對表達量比較
2.2 HIV-1感染對M8166細胞內ITAMs/ITIMs通路相關蛋白表達的影響 與空白組比較,感染組SHP-1、p-SHP-1蛋白相對表達量高(P均<0.05),SHIP1、p-SHIP1蛋白相對表達量低(P均<0.05),Syk蛋白相對表達量高、p-Syk蛋白相對表達量低,但差異無統計學意義(P均>0.05);兩組SHP-2、SHIP2、p-SHIP2蛋白相對表達量比較差異無統計學意義(P均>0.05);兩組均未見p-SHP2、Zap70、p-Zap70蛋白表達。見表2、3。
表2 兩組ITAMs/ITIMs通路相關蛋白表達比較
表3 兩組ITAMs/ITIMs通路相關磷酸化蛋白表達比較
3 討論
自20世紀80年代AIDS被發現以來,各國都投入了大量資金和科研人才對HIV進行研究。盡管如此,目前AIDS仍是危害全人類健康的一種致死性疾病。CD4+T細胞是HIV-1感染的主要靶細胞。M8166細胞是CD4+T淋巴細胞的一種,為T淋巴母細胞系,被HIV-1感染后能夠產生明顯的細胞病變[10,11]。M8166細胞是對HIV/猴免疫缺陷病毒(SIV)最為敏感的細胞系之一[12],常用于與HIV/SIV相關的分子病毒學研究[13]。本研究選用感染HIV-1的M8166細胞為實驗模型,可更準確地模擬人體感染HIV-1后的免疫環境,具有安全、高效、經濟、便利等特點,特別適用于HIV-1感染后免疫調控通路的相關研究。
由頓挫型感染引發的細胞焦亡是HIV-1感染導致CD4+T細胞死亡的主要方式[14~16]。細胞焦亡是新近發現的一種依賴炎性Caspase(Caspase-1)發生的細胞程序性死亡。這種死亡方式由微生物感染誘導,也可由內源性損傷相關信號引發,在傳染性疾病和免疫性疾病中扮演重要角色[17~19]。HIV-1感染機體后,體內腸道黏膜損傷,大量CD4+T細胞死亡,微生物移位,脂多糖水平升高,從而引發免疫活化和炎癥狀態。這是HIV-1感染者機體內持續過度炎癥反應的重要原因[15]。因此,細胞焦亡是一種促免疫激活、促炎癥的細胞死亡形式,AIDS疾病進展與細胞焦亡密切相關。
在接受高效抗逆轉錄病毒治療(HARRT)的HIV-1感染者的自然殺傷細胞和巨噬細胞中發現,包含ITAMs的受體λ表達降低[8,9],但未對感染者ITAMs/ITIMs的異常有深入闡釋。由包含ITAMs/ITIMs的受體所介導的免疫激活和炎癥調控機制可以調節HIV-1感染導致的劇烈炎癥反應,使其不至于過度反應而傷害自身,而AIDS的發生發展正是由HIV-1感染引發的持續過度免疫激活和炎癥反應導致。目前關于HIV-1感染造成的細胞焦亡的研究報道較少,其免疫調控通路的作用機制更未見報道,而深入研究ITAMs/ITIMs免疫信號調控通路異常的機制與細胞焦亡的內在關系,可進一步闡明HIV-1感染者炎癥反應持續過度的機制,為AIDS治療開辟除HARRT法以外的新途徑。
本研究結果顯示,HIV-1感染M8166細胞后,細胞內SHP-1 mRNA表達升高,SHP-1、P-SHP-1蛋白表達上調,而SHIP1 mRNA表達有下降趨勢,SHIP1、P-SHIP1蛋白表達降低。表明細胞感染HIV-1后,ITAMs/ITIMs信號調控通路被部分激活,并可能通過促進SHP-1的表達及磷酸化或抑制SHIP1的表達及磷酸化發揮作用,使疾病進一步發展。提示HIV-1感染造成的CD4+T細胞焦亡可能通過ITAMs/ITIMs信號通路發揮作用。
參考文獻:
[1] Li Q, Liu Z, Yang J, et al. Comment on the intervention of Traditional Chinese Medicine on survival rates of patients living with human immunodeficiency virus and acquired immune deficiency syndrome[J]. J Tradit Chin Med, 2016,36(3):409-410.
[2] Ipp H, Zemlin AE, Erasmus RT, et al. Role of inflammation in HIV-1 disease progression and prognosis[J]. Crit Rev Clin Lab Sci, 2014,51(2):98-111.
[3] Waterman PM, Cambier JC. The conundrum of inhibitory signaling by ITAM-containing immunoreceptors: potential molecular mechanisms[J]. FEBS Lett, 2010,584(24):4878-4882.
[4] Ivashkiv LB. How ITAMs inhibit signaling[J]. Sci Signal, 2011,4(169):pe20.
[5] Blank U, Launay P, Benhamou M, et al. Inhibitory ITAMs as novel regulators of immunity[J]. Immunol Rev, 2009,232(1):59-71.
[6] Kane BA, Bryant KJ, McNeil HP, et al. Termination of immune activation: an essential component of healthy host immune responses[J]. J Innate Immun, 2014,6(6):727-738.
[7] Lambert AA, Barabé F, Gilbert C, et al. DCIR-mediated enhancement of HIV-1 infection requires the ITIM-associated signal transduction pathway[J]. Blood, 2011,117(24):6589-6599.
[8] Leeansyah E, Zhou J, Paukovics G, et al. Decreased NK Cell FcRgamma in HIV-1 infected individuals receiving combination antiretroviral therapy: a cross sectional study[J]. PLoS One, 2010,5(3):pe9643
[9] Lichtfuss GF, Cheng WJ, Farsakoglu Y, et al. Virologically suppressed HIV patients show activation of NK cells and persistent innate immune activation[J]. J Immunol, 2012,189(3):1491-1499.
[10] Speth C, Schabetsberger T, Mohsenipour I, et al. Mechanism of human immunodeficiency virus-induced complement expression in astrocytes and neurons[J]. J Virol, 2002,76(7):3179-3188.
[11] Speth C, St?ckl G, Mohsenipour I, et al. Human immunodeficiency virus type 1 induces expression of complement factors in human astrocytes[J]. J Viroly, 2001,75(6):2604-2615.
[12] Nagao T, Yoshida A, Sakurai A, et al. Determination of HIV-1 infectivity by lymphocytic cell lines with integrated luciferase gene[J]. Int J Mol Med, 2004,14(6):1073-1076.
[13] Miyake A, Ishida T, Yamagishi M, et al. Inhibition of active HIV-1 replication by NF-kappaB inhibitor DHMEQ[J]. Microbes Infect, 2010,12(5):400-408.
[14] Doitsh G, Galloway NL, Geng X, et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection[J]. Nature, 2014,505(7484):509-514.
[15] Doitsh G, Cavrois M, Lassen KG, et al. Abortive HIV infection mediates CD4 T cell depletion and inflammation in human lymphoid tissue[J]. Cell, 2010,143(5):789-801.
[16] Monroe KM, Yang Z, Johnson JR, et al. IFI16 DNA sensor is required for death of lymphoid CD4 T cells abortively infected with HIV[J]. Science, 2014,343(6169):428-432.
[17] Cookson BT, Brennan MA. Pro-inflammatory programmed cell death[J]. Trends Microbiol, 2001,9(3):113-114.
[18] Lamkanfi M, Dixit VM. Mechanisms and functions of inflamma-somes[J]. Cell, 2014,157(5):1013-1022.
[19] Jorgensen I, Miao EA. Pyroptotic cell death defends against intra-cellular pathogens[J]. Immunol Rev, 2015,265(1):130-142.
