長鏈非編碼RNA lucat1對胃癌細胞侵襲和遷移能力的影響及其機制
2018-06-19 08:20:32于貴林林杰趙國華艾福祿王越
山東醫藥 2018年20期
關鍵詞:胃癌
于貴林,林杰,趙國華,艾福祿,王越
(遼寧省腫瘤醫院,沈陽110042)
長鏈非編碼RNA(lncRNA)是一類長度大于200個核苷酸、不具有編碼能力的核苷酸序列[1,2]。大量研究表明,lncRNA廣泛參與腫瘤細胞的多種生物學過程,如細胞增殖、分化、凋亡等[3];還可通過調節轉錄、調節各種轉錄后過程和招募染色質重組復合物等功能發揮作用[4]。識別癌相關的lncRNA并研究其分子和生物學作用具有重要意義。既往研究發現,胃癌細胞lncRNA lucat1過表達,其過表達與胃癌細胞侵襲和遷移能力呈正相關關系。有報道,甲酰肽受體2(FPR2)表達可影響上皮-間充質轉化(EMT)而參與胃癌細胞的侵襲和轉移[5]。前期我們采用生物信息學方法預測,lncRNA lucat1可能通過調節FPR2發揮作用。2017年3~8月,本研究對這一預測結果進行驗證。
1 材料與方法
1.1 材料 人胃腺癌細胞SGC-7901、BGC-823及正常胃黏膜細胞GES-1取自遼寧省腫瘤醫院中心實驗室,置于含10% FBS的RPMI 1640培養基(Invitrogen公司)中,37 ℃、50 mL/L CO2的培養箱中培養,間隔2 天更換培養基1次。FBS DMEM培養液(美國Gibico公司),CO2無菌培養箱(美國Thermo公司); lucat1 shRNA慢病毒載體及FPR2過表達質粒(ex-FPR2)由上海吉凱生物工程有限公司構建,逆轉錄試劑盒(武漢博士德生物工程有限公司),定量引物(上海吉凱生物工程有限公司),Transwell小室(美國Corning公司),稀釋基質膠(美國BD公司)。
1.2 SGC-7901、BGC-823、GES-1細胞lncRNA lucat1和FPR2 mRNA表達檢測 采用實時熒光定量PCR法。采用TRIzol法提取各細胞總RNA并檢測RNA的純度及濃度。將總RNA逆轉錄為cDNA,用ROCHE實時定量熒光PCR儀行實時熒光定量PCR擴增。lncRNA lucat1上游引物序列5′-ACCAGCTGTCCCTCAGTGTTCT-3′,下游引物序列5′-AGGCCTTTATCCTCGGGTTGCCT-3′;FPR2上游引物序列5′-GAGACCTCAGCTGGTTGTGC-3′,FPR2下游引物序列5′-ACCACCACTTCTGATCCATTCA-3′;內參U6上游引物序列5′-TTATGGGTCCTAGCCTGAC-3′,下游引物序列5′-CACTATTGCGGGCTGC-3′。采用2-ΔΔCt法計算lncRNA lucat1和FPR2 mRNA相對表達量。
1.3 下調lucat1表達對胃癌細胞SGC-7901、BGC-823 FPR2 mRNA表達影響的觀察 將體外培養的胃癌細胞SGC-7901、BGC-823分為觀察組和對照組。觀察組用Lipofectamine2000轉染lncRNA lucat1 shRNA,按操作手冊操作;對照組不轉染。lncRNA lucat1 shRNA正義序列:5′-UUAAGAAGUAGAACACUGAGGGACA-3′,反義序列5′-UGUCCCUCAGUGUUCUACUUCUUAA-3′。轉染48 h,采用1.2中的方法檢測兩組SGC-7901、BGC-823細胞FPR2 mRNA相對表達量。
1.4 轉染lncRNA lucat1 shRNA及FPR2過表達質粒對胃癌細胞BGC-823侵襲和遷移能力影響的觀察 將體外培養的BGC-823細胞分為A、B、C組。A組轉染lncRNA lucat1 shRNA,B組轉染lncRNA lucat1 shRNA和FPR2過表達質粒,方法同1.3。C組不轉染。轉染48 h,觀察各組細胞侵襲和轉移能力。① Transwell小室實驗。在每個8 μm孔徑的Transwell小室內加入50 μg ECM 膠進行包被,37 ℃無菌條件下放置6 h。ECM膠充分凝固后,上室接種細胞2×105/孔后加入無抗生素、無血清培養基,下室加入含10%血清的無抗生素培養基,繼續培養 24 h。將小室取出,用無菌棉簽擦拭去除上室內細胞,以1%結晶紫染液將小室多孔膜下表面的細胞染色,拍照。計數紫色染色的穿膜細胞數。②劃痕實驗。將細胞接種于6孔板,100 μL槍頭垂直于孔板制作細胞劃痕,盡量保證各個劃痕寬度一致。吸去細胞培養液,用PBS沖洗孔板3次,洗去劃痕產生的細胞碎片。加入無血清培養基,拍照記錄。將培養板放入培養箱培養,48 h后再取出拍照,計算遷移距離(熒光顯微鏡拍照,以96孔中心陰影區域為參照,劃痕在圖片的正中)。
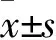
2 結果
2.1 SGC-7901、BGC-823、GES-1細胞lncRNA lucat1和FPR2 mRNA相對表達量比較 SGC-7901、BGC-823、GES-1細胞lncRNA lucat1相對表達量分別為3.651±0.332、3.023±0.331、1.000±0.098,FPR2 mRNA相對表達量分別為3.354±0.433、3.425±0.354、1.000±0.277。SGC-7901、BGC-823細胞lncRNA lucat1、FPR2 mRNA相對表達量均高于GES-1細胞(P均<0.05)。
2.2 下調lncRNA lucat1表達對SGC-7901、BGC-823細胞FPR2 mRNA表達的影響 觀察組、對照組SGC-7901細胞FPR2 mRNA相對表達量分別為1.00±0.05、0.48±0.04,BGC-823細胞FPR2 mRNA相對表達量分別為1.00±0.04 、 0.48±0.04,兩組比較P均<0.05。
2.3 轉染lncRNA lucat1 shRNA及FPR2過表達質粒對胃癌細胞BGC-823侵襲和遷移能力的影響 Transwell小室實驗顯示,A、B、C組穿膜細胞數分別為(143±7)、(194±9)、(233±8)個,三組穿膜細胞數兩兩比較P均<0.05。劃痕實驗顯示,A、B、C組細胞遷移距離分別為(0.342±0.027)、(0.534±0.021)、(0.652±0.029)mm,三組細胞遷移距離兩兩比較P均<0.05。
3 討論
lncRNA表達改變是腫瘤發生的驅動因素之一。lncRNA可在不同水平調節基因表達[6]:部分lncRNA首先作為信號與蛋白質結合(如激素受體和轉錄因子),然后復合物與啟動子位點結合,調節下游基因表達; lncRNA可作為引導調控蛋白到靶向基因啟動子位點的指引;部分lncRNA作為誘餌阻止調控蛋白與啟動子位點結合,或作為支架來招募蛋白復合物。
lucat1又名SCAL1,定位于5號染色體,2013年首次在吸煙者的呼吸道上皮細胞中被發現,在非小細胞肺癌細胞系中高表達[7]。有文獻報道,lucat1可通過降低人非小細胞肺癌組織p21和p57表達而調控腫瘤細胞周期和增殖[8];并能誘導漿液性卵巢癌對順鉑的耐藥[9]。Thai等[10]報道,lucat1可通過DNA甲基化調控甲基轉移酶1(DNMT1)的穩定性,抑制抑癌基因表達,從而促進腫瘤的形成。Sun等[11]報道,miR-200c可通過RNA海綿的作用調節ABCB1和lucat1表達。目前lucat1在胃癌侵襲和遷移中的生物學作用和調控機制仍不清楚。本研究結果顯示,與胃黏膜上皮細胞相比,BGC-823細胞lncRNA lucat1呈過表達;下調lncRNA lucat1表達可抑制細胞的侵襲和遷移能力。證實lncRNA lucat1過表達參與胃癌的侵襲和遷移。
FPR家族成員共包括FPR1、FPR2和FPR3,三者均在胃癌細胞中表達[12]。FPR2是一種典型的G蛋白偶聯受體的化學吸引受體,通過與不同配體和不同細胞結合而被激活,其激活后可觸發不同的信號通路:如磷脂酶C、蛋白激酶C、磷酸肌醇3激酶/蛋白激酶B、絲裂原活化蛋白激酶等[13,14],這些通路均參與腫瘤細胞的侵襲和遷移。為研究lncRNA lucat1調控胃癌細胞侵襲和遷移能力的機制,本研究進一步觀察了下調lncRNA lucat1表達對BGC-823細胞FPR2表達的影響,結果顯示轉染lncRNA lucat1 shRNA的觀察組FPR2 mRNA相對表達量高于對照組。本研究還觀察了轉染lncRNA lucat1 shRNA及FPR2過表達質粒對胃癌細胞BGC-823侵襲和遷移能力的影響,結果顯示共轉染lncRNA lucat1 shRNA及FPR2過表達質粒的B組BGC-823細胞Transwell小室實驗的穿膜細胞數和劃痕實驗的細胞遷移距離均大于轉染lncRNA lucat1 shRNA的A組,表明lncRNA lucat1可能通過FPR2調控胃癌細胞侵襲和遷移。
綜上所述,lncRNA lucat1可促進胃癌細胞的侵襲和遷移,其機制可能與促進FPR2表達有關,但其具體分子生物學機制仍需進一步研究探討。lncRNA lucat1和FPR2有望成為胃癌診斷和靶向治療的新靶點。
參考文獻:
[1] Siegel RL,Miller KD,Jemal A. Cancer Statistics,2017[J]. CA: Cancer J Clin,2017,67(1):7-30.
[2] Chen W,Zheng R,Baade PD,et al. Cancer statistics in China,2015[J]. CA: Cancer J Clin,2016,66(2):115-132.
[3] Wang XN,Liang H. Some problems in the surgical treatment of gastric cancer[J]. Chin J Cancer,2010,29(4):369-373.
[4] Ponting CP,Oliver PL,Reik W. Evolution and functions of long noncoding RNAs[J]. Cell,2009,136(4):629-641.
[5] Schmitz SU,Grote P,Herrmann BG. Mechanisms of long noncoding RNA function in development and disease[J]. Cell Mol Life Sci,2016,73(13):2491-2509.
[6] Batista PJ,Chang HY. Long noncoding RNAs: cellular address codes in development and disease[J]. Cell,2013,152(6):1298-1307.
[7] Khalil AM,Guttman M,Huarte M,et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression[J]. Proc Nat Acad Sci U S A,2009,106(28):11667-11672.
[8] Hou XL,Ji CD,Tang J,et al. FPR2 promotes invasion and metastasis of gastric cancer cells and predicts theprognosis of patients[J]. Sci Rep,2017,7(1):3153.
[9] Kung JT,Colognori D,Lee JT. Long noncoding RNAs: past,present,and future[J]. Genetics,2013,193(3):651-669.
[10] Thai P,Statt S,Chen CH,et al. Characterization of a novel long noncoding RNA,SCAL1,induced by cigarette smoke and elevated in lung cancer cell lines[J]. Am J Respir Cell Mol Biol,2013,49(2):204-211.
[11] Sun Y,Jin SD,Zhu Q,et al. Long non-coding RNA LUCAT1 is associated with poor prognosis in human non-small lung cancer and regulates cell proliferation via epigenetically repressing p21 and p57 expression[J]. Oncotarget,2017,8(17):28297-28311.
[12] Zheng ZG,Xu H,Suo SS,et al. The essential role of H19 contributing to cisplatin resistance by regulating glutathione metabolism in high-grade serous ovarian cancer[J]. Sci Rep,2016(6):26093.
[13] Yoon JH,You BH,Park CH,et al. The long noncoding RNA LUCAT1 promotes tumorigenesis by controlling ubiquitination and stability of DNA methyltransferase 1 in esophageal squamous cell carcinoma[J]. Cancer Letters,2018(417):47-57.
[14] Han Z,Shi L. Long non-coding RNA LUCAT1 modulates methotrexate resistance in osteosarcoma via miR-200c/ABCB1 axis[J]. Biochem Biophysi Res Commun,2018,495(1):947-953.
[15] de Paulis A,Prevete N,Rossi FW,et al. Helicobacter pylori Hp(2-20) promotes migration and proliferation of gastric epithelial cells by interacting with formyl peptide receptors in vitro and accelerates gastric mucosal healing in vivo[J]. J Immun,2009,183(6):3761-3769.
[16] Reddy KB,Nabha SM,Atanaskova N. Role of MAP kinase in tumor progression and invasion[J]. Cancer Metastasis Rev,2003,22(4):395-403.
[17] Coffelt SB,Tomchuck SL,Zwezdaryk KJ,et al. Leucine leucine-37 uses formyl peptide receptor-like 1 to activate signal transduction pathways,stimulate oncogenic gene expression,and enhance the invasiveness of ovarian cancer cells[J]. Mol Cancer Res,2009,7(6):907-915.
猜你喜歡
昆明醫科大學學報(2022年1期)2022-02-28 07:43:36
昆明醫科大學學報(2021年5期)2021-07-22 07:32:22
基層中醫藥(2020年2期)2020-07-27 02:46:06
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
中外醫療(2015年18期)2016-01-04 06:51:55
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:25
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
中國當代醫藥(2015年20期)2015-03-01 02:04:29
