慢病毒介導的miR194-5p過表達精原細胞株的建立*
2018-05-17 01:41:24夏蒙蒙牛長敏申雪沂王建軍
中國男科學雜志 2018年2期
夏 靜 夏蒙蒙 牛長敏 申雪沂 王建軍 鄭 英
揚州大學醫學院組織學與胚胎學教研室(江蘇揚州 225001)
miRNA是一類長約18-22nt的小分子非編碼RNA,它們能結合在多個下游靶基因的3′UTR(3’端非編碼區),通常導致靶基因的表達沉默,進而在一系列生物學過程中發揮重要功能[1]。研究表明miRNA也參與精子發生過程的調控,且有表達細胞特異性及階段性[2]。一些特異性的miRNA能作為診斷男性不育的潛在標記[3,4]。Stra8(stimulated by retinoic acid 8)對精子發生具有重要意義,其對減數分裂的啟動是必須的,同時它對精原細胞的分化也發揮著重要的作用,但是其調控精子發生的機制尚不清楚。miRNA是否能通過Stra8調控精子發生過程尚不清楚。利用生物信息學分析方法,我們發現Stra8是miR-194-5p下游靶基因之一,因此我們試圖探討miR-194-5p與Stra8之間是否具有調控關系。本研究旨在構建攜帶miR-194-5p的慢病毒表達載體,并包裝成慢病毒,以期達到高效穩定感染目的細胞Stra8-GC1-spg,建立穩定表達miR-194-5p的精原細胞株,為深入研究miR-194-5p與Stra8之間的調控關系及闡明Stra8在精子發生中的作用機制提供參考價值。
材料和方法
一、材料
慢病毒表達質粒pMSCV-PIG及Gag pol、VSV.G輔助質粒由江蘇省非編碼RNA基礎與臨床轉化重點實驗室郁多男教授惠贈。LA tag酶、EcoRⅠ BglⅡ、miRNA逆轉錄試劑盒及熒光定量試劑盒購自TaKaRa公司。pGEM-T-easy 載體、T4 DNA連接酶、切膠回收試劑盒購自Promega公司。293T細胞、Stra8-GC1-spg、大腸桿菌JM109由本實驗室前期保存。DMEM、FBS購自Hyclone公司;X-gal和IPTG、嘌呤霉素、PEI購自北京索萊寶生物有限公司;核酸提取試劑TRIzol購自Invitrogen公司;引物合成及DNA測序由深圳華大基因生物工程公司完成。
二、方法
(一)目的片段的擴增
利用小鼠基因組DNA為模板,進行PCR擴增反應,PCR擴增引物:F:5′-GGAGATCTCCACCA CACACCAGGAAGAG-3′,R: 5′-GGGAATTCT AACCAAGAGCCCACCCAG-3′;PCR反應體系:ddH2O 31.5μL,LA buffer 5μL,2.5mMdNTPs 8μL,F(10μM)2μL,R(10μM)2μL,LA tag酶0.5μL,DNA 1μL;PCR反應條件:95℃ 5min,95℃ 30s,58℃ 30 s,72℃ 60 s,共35個循環,72℃ 5min。PCR產物經瓊脂糖凝膠電泳鑒定。
(二)pGEM-T-easy-miR194-5p重組質粒的構建
PCR產物經AxyPrep DNA凝膠回收試劑盒進行純化,后與pGEM-T-easy 載體連接,連接產物轉化到JM109感受態中,涂于LB瓊脂糖平板(含50μg /μL X-gal和0.1moL IPTG、50μg /mL氨芐青霉素)上。37℃孵育過夜,經藍白斑篩選,取白色克隆,搖菌過夜,堿裂解法提取質粒,EcoRⅠ酶切鑒定,最后進行DNA序列測定驗證。
(三)pMSCV-PIG-miR194-5p重組載體構建
將pGEM-T-easy-miR194-5p及慢病毒載體pMSCV-PIG分別進行EcoRⅠ、BglⅡ雙酶切,回收并純化所需片段,以載體插入片段=1:3~1:5摩爾比進行連接,轉化JM109感受態菌后涂于LB瓊脂糖平板(含50μg /mL氨芐青霉素)上,37℃過夜。次日,挑單菌落接種于LB培養液(50μg /mL氨芐青霉素)中,37℃振搖培養過夜,用堿裂解法提取質粒,利用EcoRⅠ、BglⅡ雙酶切鑒定。
(四)慢病毒包裝
將293T細胞鋪于6cm細胞培養皿內,使次日細胞融合率為60%左右。每孔加3mL DMEM完全培養基。慢病毒包裝轉染體系:0.9%NaCl 500μL、Gag pol 3μg 、VSV.G 3μg、pMSCV-PIG-miR194-5p 8μg(實驗組)或pMSCV-PIG 8μg(對照組)、PEI 100μL(μg/μL),混勻后室溫孵育10min,加至培養的293T細胞中,37℃、5%CO2過夜,12h后更換為DMEM完全培養基。分別收集轉染后36h、48h、60h、72h的病毒上清,經0.45μm濾器過濾,-80℃保存。
(五)慢病毒感染效果的驗證
將293T細胞鋪于6cm細胞培養皿內,使次日細胞融合率為60%左右。按照以下體系感染293T細胞:慢病毒上清 2mL、enhance 液1mL、polybrene 1.5μL,37℃、5%CO2培養箱培養。6h后進行重復感染一次。6h后再進行第三次感染。6h后更換為新鮮的DMEM完全培養基。72h后用倒置熒光顯微鏡觀察感染的293T細胞有無綠色熒光。分別提取pMSCV-PIG-miR194-5p感染組及 pMSCV-PIG對照組293T細胞總RNA,利用逆轉錄試劑盒反轉錄為cDNA,qRTPCR驗證感染慢病毒的293T細胞內的miR194-5p轉錄水平。
(六)穩定過表達miR194-5p Stra8-GC1-spg細胞株的建立
將Stra8-GC1-spg細胞鋪于六孔板內,使次日細胞融合率為50%左右。在六孔板每孔內加入enhance液0.5mL、慢病毒上清1.5mL 、polybrene1μL。37℃、5%CO2培養箱培養。6h后進行重復感染一次。6h后再進行第三次感染。6h后更換為新鮮的DMEM完全培養基。72h后用倒置熒光顯微鏡觀察細胞是否帶有綠色熒光。用5μg/mL嘌呤霉素篩選一周,以獲得穩定過表達miR194-5p Stra8-GC1-spg細胞株。
(七)miR194-5p表達水平的驗證
分別取感染慢病毒pMSCV-PIG-miR194-5p及對照組pMSCV-PIG的Stra8GC1細胞,用TRIzol提取總RNA,按照逆轉錄試劑盒說明書步驟進行反轉錄,以此cDNA為模板,應用實時定量PCR法檢測miR194-5p的表達水平,并用2-ΔΔct計算其相對表達量,兩組間數據比較采用t檢驗,P<0.05有統計學意義。
結 果
一、目的基因片段pri-miR194-5p的擴增
以小鼠基因組DNA為模板,進行PCR反應擴增,獲得目的基因片段pri-miR194-5p,其大小為293bp,與預計片段大小相一致(圖1)。
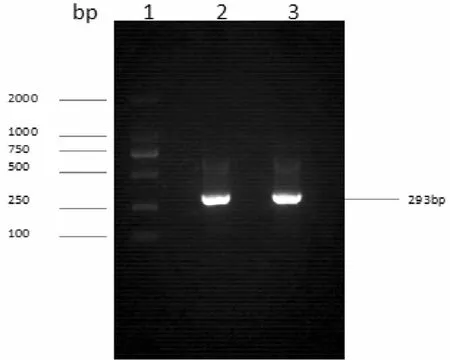
圖1 pri-miR194-5p PCR產物瓊脂糖凝膠電泳圖
二、pri-miR194-5p與pGEM-T-easy 載體的連接
將pri-miR194-5p PCR產物與pGEM-T-easy載體相連接,經酶切鑒定證實PCR片段已成功插入到pGEMT-easy載體(圖2)。DNA序列測定證實此序列與pri-miR194-5p序列完全一致。表明pGEM-T-easy-primiR194-5p重組載體構建成功。
三、pMSCV-PIG-miR194-5p重組載體的構建
將重組質粒pGEM-T-easy-pri-miR194-5p及空載慢病毒載體pMSCV-PIG分別進行雙酶切,然后進行連接,獲得的重組質粒pMSCV-PIG-miR194-5p經限制性內切酶酶切鑒定,發現重組質粒pMSCV-PIG-miR194-5p構建成功(圖3)。
四、慢病毒的包裝及表達的驗證
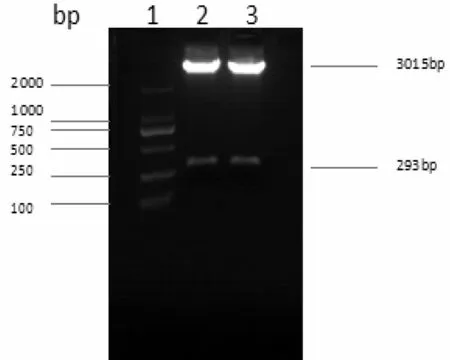
圖2 pGEM-T-easy-pri-miR194-5p酶切鑒定產物瓊脂糖凝膠電泳圖
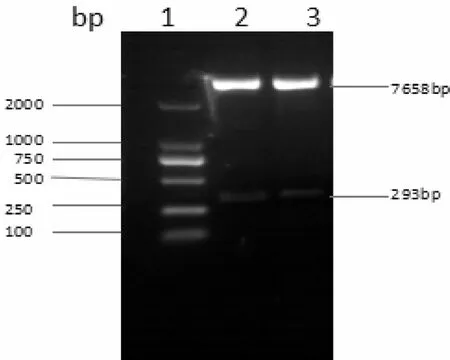
圖3 pMSCV-PIG-miR194-5p酶切鑒定

圖4 pMSCV-PIG及pMSCV-PIG-miR194-5p重組質粒轉染293T細胞72h后熒光圖

圖5 慢病毒pMSCV-PIG及pMSCV-PIG-miR194-5p感染293T細胞72h后熒光圖
應用PEI法轉染293T包裝細胞,72h后熒光顯微鏡觀察發現大多數293T細胞均顯示綠色熒光(圖4)。用慢病毒上清感染293T細胞,72h后熒光顯微鏡觀察,發現大部分293T細胞顯示綠色熒光(圖5)。表明包裝的慢病毒顆粒能成功感染細胞。應用實時定量PCR對感染后的293T細胞進行miR194-5p表達水平檢測,結果發現,與對照組相比較,實驗組細胞表達高水平miR194-5p(圖6)。提示慢病毒能有效感染293T細胞并高表達miR194-5p。
五、過表達miR194-5p Stra8-GC1-spg細胞株的建立
應用慢病毒上清感染Stra8-GC1-spg。72h后熒光顯微鏡觀察,發現細胞有綠色熒光產生(圖7)。經嘌呤霉素篩選1周后,對miR194-5p表達水平進行定量分析。結果發現與對照組相比較,實驗組Stra8-GC1-spg細胞中miR194-5p的表達量明顯增高(圖8),表明成功穩定過表達miR194-5p Stra8-GC1-spg細胞株。
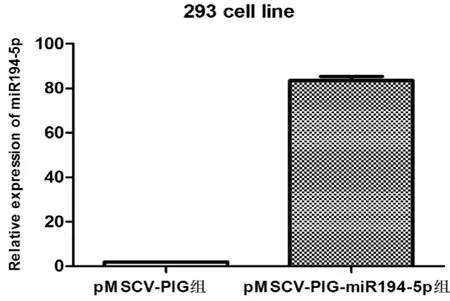
圖6 慢病毒pMSCV-PIG及pMSCV-PIG-miR194-5p感染的293T細胞中miR194-5p相對表達水平分析
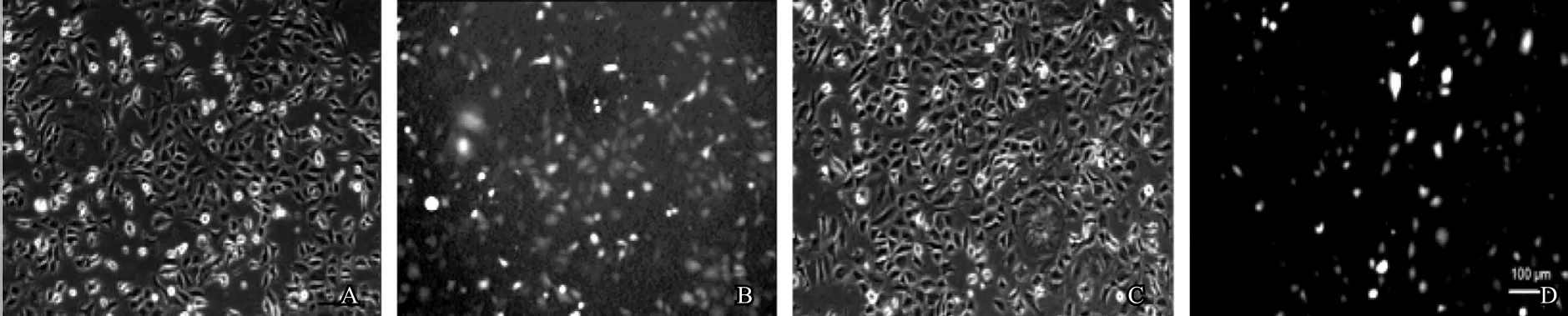
圖7 慢病毒pMSCV-PIG及pMSCV-PIG-miR194-5p感染Stra8-GC1-spg細胞72h后熒光圖
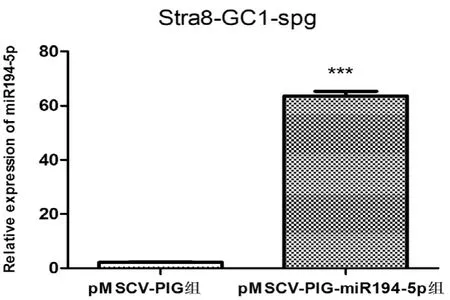
圖8 慢病毒pMSCV-PIG及pMSCV-PIG-miR194-5p感染的Stra8-GC1-spg細胞中miR194-5p的相對表達水平分析
討 論
精子發生是高度精密的過程,此期間需要大量的基因在轉錄、轉錄后及表觀遺傳學水平對其進行調控。大量研究發現在精子發生的不同階段,miRNAs作為轉錄后調控元件,可能通過激活特異性基因的表達參與精子發生[5]。在人類精子發生中,miRNAs也發揮著重要的功能[6,7]。 研究發現在小鼠精子發生中一些miRNAs(miR-221, miR-203, miR-34b-5p)特異性表達于精原干細胞、減數分裂前細胞及減數分裂細胞中[8]。 miR-34c特異性表達在睪丸內,且主要表達在減數分裂晚期的粗線期精母細胞及精子細胞中[9]。 miR-449a 及miR-34b/c在鼠科精子發生中有協同作用[10]。miR-146能作用在靶基因Med1上進而阻礙視磺酸誘導精原細胞分化[11]。 另外,miR-221/222家族通過作用在Kit mRNA 上,進而維持未分化精原細胞數量[12]。
目前已知Stra8作為精子發生中一個基因,對精原細胞的分裂分化及減數分裂的啟動都發揮著重要的作用。研究發現Stra8基因敲除的雄性小鼠不育[13]。但是關于Stra8在精子發生中的作用機制目前仍未能完全闡明。Yu等發現miR-34c可能通過作用在Nanos2的3’UTR上抑制其表達,而這將導致Stra8 、Scp3及 Dazl的表達上調,而促進減數分裂進程[14]。Wang等發現在雞的睪丸內miR-31 可以通過作用在靶基因Stra8的3’UTR(非編碼區)上直接抑制Stra8的表達進而阻滯減數分裂的進程[15]。而有關miRNAs與Stra8之間的關系目前仍未完全闡明。信息學分析發現mmu-miR-194-5p是Stra8上游的靶向miRNA。而目前有關miR-194-5p的研究僅僅是著重在它在各種腫瘤及免疫性疾病上的作用[16-19]。而有關miR-194-5p與Stra8之間的調控關系及它們在精子發生中的作用目前仍未有任何報道。是否有可能miR-194-5p可以在精子發生的特定階段被激活,從而發揮對Stra8的調控作用呢?因此研究miR-194-5p與Stra8之間的關系對揭示Stra8在精子發生中的作用機制具有十分重要的意義。
本實驗室前期工作中已構建了Stra8過表達精原細胞株,將其命名為Stra8-GC1-spg。為了進一步研究miR194-5p與Stra8之間的調控關系,本研究采用了慢病毒重組質粒pMSCV-PIG-miR194-5p來包裝293T細胞,產生慢病毒顆粒。為了檢測該慢病毒是否包裝成功,我們將獲得的病毒上清首先感染了293T細胞,通過熒光實時定量發現與對照組相比,pMSCV-PIG-miR194-5p慢病毒感染組,miR194-5p表達水平明顯增高。說明該慢病毒已經包裝成功。本實驗中所用的pMSCV-PIG空載質粒帶有GFP標簽,能使細胞產生綠色熒光。轉染293T時使用的PEI(Polyethylenimine)是一種聚合物轉染試劑,它是運用最廣泛及最有效的非病毒載體之一[20]。PEI每相隔二個碳原子就有1個氨基,構成了多聚網狀結構,它具有大量的正電荷,能使 DNA 縮合成顆粒狀。PEI/DNA 復合物通過靜電作用吸附到細胞表面,經過細胞內吞作用進入細胞內[21]。PEI 幾乎可在任何 pH 條件下都能夠吸收質子,這使其具有質子海綿的作用[22]。這一特性使 PEI能在內涵體的酸性環境中吸收 H+,進而增高內涵體內的滲透壓,導致膜不穩定甚至破裂,從而使PEI/DNA復合物從內涵體內逃逸出來,避免 DNA 降解。一般認為,PEI 的基因轉染效率隨其相對分子質量的增大而提高。這是由于 PEI 相對分子質量越大,其質子海綿作用越強,PEI/DNA 復合物就更快的被釋放入胞質,從而減少了內涵體對 DNA 的降解,進而提高了轉染效率[23]。重要的是PEI以鹽酸鹽形式提供,大大提高了水溶性。與Lipofectamine 2000相比,PEI轉染效率更高,且細胞凋亡率更低[24]。我們用該慢病毒感染Stra8-GC1-spg細胞72h后,嘌呤霉素篩選獲得穩定表達miR194-5p的Stra8-GC1-spg細胞株。通過熒光實時定量分析發現miR194-5p 在Stra8-GC1-spg細胞中miR194-5p的表達水平與對照組相比顯著性增高。當我們應用瞬時轉染法轉染精原細胞株時轉染效率很低,但用此方法包裝出的慢病毒感染Stra8-GC1-spg細胞使感染效率大大提高,因而更加有利于后續實驗的開展。本研究采用的慢病毒系統優于一般的商品化的慢病毒感染系統,既具有GFP熒光示蹤系統,又能夠通過嘌呤霉素進行陽性細胞的篩選。
總之,本實驗成功構建了過表達miR194-5p 的Stra8-GC1-spg細胞株。這種細胞系可以作為一種細胞模型用于后續研究miR194-5p與Stra8之間的調控關系,為進一步揭示Stra8在精子發生中的作用機制奠定基礎。
參考文獻
1 Pritchard CC, Cheng HH, Tewari M.MicroRNA profiling:approaches and considerations.Nat Rev Genet2012;13(5): 358-369
2 Yang Q, Hua J, Wang L,et al.MicroRNA and piRNA profiles in normal human testis detected by next generation sequencing.PLoS One2013; 8(6): e66809
3 Rahbar S, Novin MG, Alizadeh E,et al.New insights into the expression profile of MicroRNA-34c and P53 in infertile men spermatozoa and testicular tissue.Cell Mol Biol (Noisy-le-grand)2017; 63(8): 77-83
4 Boissière A, Gala A, Ferrières-Hoa A,et al.Cell-free and intracellular nucleic acids: new non-invasive biomarkers to explore male infertility.Basic Clin Androl2017; 27: 7
5 Kotaja N.Micro RNAs and spermatogenesis.Fertil Steril2014; 101(6): 1552-1562
6 Lian J, Zhang X, Tian H,et al.Altered micro RNA expression in patients with non-obstructive azoospermia.Reprod Biol Endocrinol2009; 7: 13
7 Abu-Halima M, Backes C, Leidinger P,et al.MicroRNA expression profiles in human testicular tissues of infertile men with different histopathologic patterns.Fertil Steril2014; 101(1): 78-86.e2
8 Smorag L, Zheng Y, Nolte J,et al.MicroRNA signature in various cell types of mouse spermatogenesis: evidence for stage-specifically expressed miRNA-221, -203and -34b-5p mediated spermatogenesis regulation.Biol Cell2012;104(11): 677-692
9 Bouhallier F, Allioli N, Lavial F,et al.Role of miR-34c microRNA in the late steps of spermatogenesis.RNA2010; 16(4): 720-731
10 Bao J, Li D, Wang L,et al.MicroRNA-449 and microRNA-34b/c function redundantly in murine testes by targeting E2F transcription factor-retinoblastoma protein (E2F-pRb) pathway.J Biol Chem2012; 287(26):21686-21698
11 Huszar JM, Payne CJ.MicroRNA 146 (Mir146)modulates spermatogonial differentiation by retinoic acid in mice.Biol Reprod2013; 88(1): 15
12 Yang QE, Racicot KE, Kaucher AV,et al.MicroRNAs221 and 222 regulate the undifferentiated state in mammalian male germ cells.Development2013; 140(2): 280-290
13 Mark M, Jacobs H, Oulad-Abdelghani M,et al.STRA8-deficient spermatocytes initiate, but fail to complete, meiosis and undergo premature chromosome condensation.J Cell Sci2008; 121(Pt 19): 3233-3242
14 Yu M, Mu H, Niu Z, et al.miR-34c enhances mouse spermatogonial stem cells differentiation by targeting Nanos2.J Cell Biochem 2014; 115(2): 232-242
15 Wang Y, Zuo Q, Bi Y,et al.miR-31 Regulates Spermatogonial Stem Cells Meiosis via Targeting Stra8.J Cell Biochem2017; 118(12): 4844-4853
16 Wang SH, Wu XC, Zhang MD,et al.Long noncoding RNA H19 contributes to gallbladder cancer cell proliferation by modulated miR-194-5p targeting AKT2.Tumour Biol2016; 37(7): 9721-9730
17 Huo D, Clayton WM, Yoshimatsu TF,et al.Identification of a circulating microRNA signature to distinguish recurrence in breast cancer patients.Oncotarget2016;7(34): 55231-55248
18 Lewinska A, Adamczyk-Grochala J, Kwasniewicz E,et al.Reduced levels of methyltransferase DNMT2 sensitize human fibroblasts to oxidative stress and DNA damage that is accompanied by changes in proliferation-related miRNA expression.Redox Biol2018; 14: 20-34
19 van der Ree MH, Jansen L, Kruize Z,et al.Plasma MicroRNA Levels Are Associated With Hepatitis B e Antigen Status and Treatment Response in Chronic Hepatitis B Patients.J Infect Dis2017; 215(9): 1421-1429 20 Deng R, Yue Y, Jin F,et al.Revisit the complexation of PEI and DNA -How to make low cytotoxic and highly efficient PEI gene transfection non - viral vectors with a controllable chain length and structure?J Control Release2009; 140(1): 40-46
21 Cryan SA,O' Driscoll CM.Mechanistic studies on nonviral gene delivery to the intestine using in vitro differentiated cell culture models and an in vivo rat intestinal loop.Pharm Res2003; 20(4): 569-575
22 Kichler A, Leborgne C, Coeytaux E,et al.Polyethylenimine-mediated gene delivery: a mechanistic study.J Gene Med2001; 3(2): 135-144
23 Brunner S, Sauser T, Carotta S,et al.Cell cycle dependence of gene transfer by lipoplex, polyplex and recombinant adenovirus.Gene Ther2000; 7(5): 401-407
24 Lin CW, Jan MS, Kuo JS.The vector-related influences of autophagic microRNA delivery by Lipofectamine 2000 and polyethylenimine 25K on mouse embryonicfibroblast cells.Eur J Pharm Sci2017; 101: 11-21
