阿帕替尼對人肝癌裸鼠皮下移植瘤生長及生物鐘基因表達的影響
2018-05-15 01:13:42何曉曉熊枝繁邱夢君占靜陳仁旺楊盛力
山東醫藥 2018年8期
關鍵詞:肝癌
何曉曉,熊枝繁,邱夢君,占靜,陳仁旺,楊盛力
(1 華中科技大學同濟醫學院附屬梨園醫院,武漢430077;2 華中科技大學同濟醫學院附屬協和醫院)
近年來肝癌的發病率不斷升高,全球每年新發病例超過700 000例[1]。生物鐘是細胞自發產生和維持的,它不依賴于外界因素維持著機體的時間節律性,其作用在于產生和控制晝夜生物節律的運轉。研究證實,生物鐘對腫瘤的發生、發展具有調控作用[2]。目前在哺乳動物體內多個生物鐘基因已被確認,如CLOCK、BMAL1、CKIε、Per(Per1、Per2、Per3)、Cry(Cry1、Cry2)、DEC和NPAS2等[3,4]。阿帕替尼是一種新型的小分子抗血管生成劑,能夠靶向抑制血管內皮生長因子受體2(VEGFR-2)酪氨酸激酶活性,繼而抑制腫瘤血管生成,發揮抗腫瘤作用[5]。有研究證實,阿帕替尼靶向治療晚期胃癌或肝癌的臨床療效較好[6,7],并可延長患者總生存期[8]。Yu等[9]研究認為,生物鐘基因是一種潛在的抑癌基因,可抑制腫瘤細胞增殖,并能促進腫瘤細胞凋亡。Lin等[10,11]研究發現,在肝癌細胞或組織中存在某些生物鐘基因表達紊亂,如Per、Cry等。但目前阿帕替尼靶向治療肝癌能否影響生物鐘基因的表達尚不清楚。2017年3~5月,本研究觀察了阿帕替尼對人肝癌裸鼠皮下移植瘤生長及生物鐘基因表達的影響。現分析結果并報告如下。
1 材料與方法
1.1 材料 BALB/c裸鼠16只,雌雄各半,4~6周齡,體質量16~23 g,SPF條件下分籠喂養。人肝癌細胞株HepG2(以下稱HepG2細胞),購自中國典型物種保藏中心,液氮中保存。甲磺酸阿帕替尼片,江蘇恒瑞醫藥股份有限公司,用10% DMSO溶解,制成終濃度為0.2 g/mL溶液。所有PCR引物由美國Life Technologies公司設計合成。CO2培養箱,美國Harris公司;熒光定量PCR儀,美國Ambion公司;電泳儀,美國Bio-Rad公司;低溫高速離心機,德國Heraeus公司。RPMI 1640、FBS、胰酶,美國Hyclone公司;DMSO,美國MP Biomedicals公司;TRIzol、SYBR?Green PCR Master Mix,美國Life Technologies公司。
1.2 人肝癌裸鼠皮下移植瘤模型建立 HepG2細胞復蘇后,接種于含10% FBS RPMI 1640培養基的培養瓶中,置于37 ℃、5% CO2、飽和濕度的培養箱中。每2~3天換液1次,待細胞鋪滿瓶底80%~90%時,以1∶3比例傳代。收集傳3代對數生長期細胞,胰蛋白酶消化,計數,用PBS緩沖液制成密度為1×107個/mL的細胞懸液。取細胞懸液0.2 mL,接種于裸鼠右腋皮下,放回籠中,繼續在SPF條件下喂養。
1.3 動物分組處理 所有裸鼠接種HepG2細胞后,每日觀察一般狀況、接種部位液體吸收及成瘤情況。接種3天左右,所有裸鼠右腋皮下接種部位出現3~5 mm的腫瘤結節。隨著腫瘤結節體積的增加,所有裸鼠出現精神萎靡、活動遲緩,食欲、體質量較接種前明顯降低。至接種1周,所有裸鼠腫瘤結節體積達100 mm3左右,隨機將裸鼠分為阿帕替尼組、對照組,每組8只。阿帕替尼組給予阿帕替尼50 mg/kg灌胃,對照組給予DMSO 100 μL/20 g灌胃,每天1次,連續灌胃2周。兩組治療期間均無死亡。
1.4 相關指標觀察
1.4.1 腫瘤體積 兩組灌胃治療期間每隔2天用用游標卡尺測量腫瘤的最大徑(a)和最小徑(b)。分別于灌胃治療前,灌胃治療第3、6、9、12天以及灌胃結束24 h,計算腫瘤體積(V)。V(mm3)=ab2/2。
1.4.2 Per、CLOCK、Cry、BMAL1、CKIε mRNA表達 采用Real-time PCR法。末次灌胃結束24 h,頸椎脫臼法處死,剝離瘤體,置于液氮中,-80 ℃冰箱凍存。取兩組凍存腫瘤組織各100 mg,TRIzol法提取組織總RNA,采用紫外分光光度計鑒定總RNA純度,OD260/OD280為1.7~2.0,表明提取的總RNA純度合格,可用于后續實驗。按照SYBR?Green PCR Master Mix試劑盒說明將總RNA逆轉錄為cDNA,以cDNA為模板進行PCR擴增。引物序列:Per1上游引物5′-AGGCAACGGCAAGGACTC-3′,下游引物5′-GGCTGTAGGCAATGGAACTG-3′,溫度60.2 ℃,產物大小101 bp;Per2上游引物5′-CTACAGCAGCACCATCGTC-3′,下游引物5′-CCACTCGCAGCATC-TTCC-3′,溫度58.9 ℃,產物大小78 bp;Per3上游引物5′-TGGTGGTGGTGAATGTAAGAC-3′,下游引物5′-GGCTGTGCTCATCGTTCC-3′,溫度57.2 ℃,產物大小104 bp;Cry1上游引物5′-CAACCTCCATTCATCTTTCC-3′,下游引物5′-CTCATAGCCGACACCTTC-3′,溫度58.9 ℃,產物大小151 bp;Cry2上游引物5′-TGGGCTTCTGGGACTGAG-3′,下游引物5′-GGTAGGTGTGCTGTCTTAGG-3′,溫度57.2 ℃,產物大小136 bp;CLOCK上游引物5′-GCAGCAGCAGCAGCAGAG-3′,下游引物5′-CAGCAGAGAGAATGA-GTTGAGTTG-3′,溫度61.9 ℃,產物大小149 bp;BMAL1上游引物5′-TGCCACCAATCCATACACAGA-AG-3′,下游引物5′-TTCCCTCGGTCACATCCTACG-3′,溫度60.9 ℃,產物大小123 bp;CKIε:上游引物5′-TCAGCGAGAAGAAGATGTC-3′,下游引物5′-GA-AGAGGTTGCGGAAGAG-3′,溫度58.9 ℃,產物大小149 bp;內參β-actin上游引物5′-AGTTGCGTTACACCCTTTCTTGAC-3′,下游引物5′-GCTCGCTCCA-ACCGACTGC-3′,溫度60.0 ℃,產物大小171 bp。每個樣本設3個復孔。PCR反應體系共25 μL:SYBRGreen Mix 12.5 μL,cDNA模板2 μL,上下游引物各1 μL,ddH2O 8.5 μL;反應條件:94 ℃ 5 min,94 ℃ 2 min、59 ℃ 1 min、72 ℃ 1 min共33個循環,最后72 ℃ 10 min。以β-actin為內參,采用2-ΔΔCt法計算目的基因相對表達量。實驗重復3次,取平均值。

2 結果
2.1 兩組灌胃不同時間移植瘤體積變化 見表1。
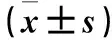
表1 兩組灌胃不同時間移植瘤體積比較
注:與對照組同時間比較,*P<0.05。
2.2 兩組移植瘤組織生物鐘基因Per、CLOCK、Cry、BMAL1、CKIε mRNA表達比較 見表2。
表2 兩組移植瘤組織Per、CLOCK、Cry、BMAL1、CKIε mRNA相對表達量比較
注:與對照組比較,*P<0.05。
3 討論
血管生成是許多腫瘤生長和轉移的重要標志,故抑制血管內皮生長因子信號通路是抗腫瘤治療的有效靶點之一。阿帕替尼為VEGFR-2小分子抑制劑,主要用于標準化療后仍有進展的晚期胃癌患者。研究證實,阿帕替尼具有廣譜抗實體瘤效果,可用于胃癌、非小細胞肺癌、乳腺癌、肝癌等多種腫瘤的靶向治療[12]。本研究結果顯示,阿帕替尼組從灌胃第3天開始腫瘤體積明顯低于對照組,說明阿帕替尼能抑制腫瘤生長,進一步驗證了阿帕替尼的抗腫瘤效果,與文獻[13]報道基本一致。
研究證實,生物鐘基因突變與惡性腫瘤的發生、發展密切相關,尤其是腹部惡性腫瘤患者生物鐘基因普遍存在異常表達[14]。腫瘤細胞中生物鐘基因Per、Cry通常處于低表達狀態[15,16];當這些生物鐘基因過表達時,可誘導腫瘤細胞凋亡,抑制腫瘤生長[17,18]。生物鐘基因CLOCK在腫瘤組織中多呈過表達狀態,其過表達會促進腫瘤細胞侵襲和遷移[19]。目前CKIε表達與惡性腫瘤的關系尚不十分清楚。Kou等[13]研究發現,腫瘤細胞內的一些生物鐘基因,如Per、Cry、CLOCK、BMAL1等,可影響VEGF的轉錄調控。生物鐘基因表達紊亂及其引起的代謝紊亂是肝癌等多種惡性腫瘤發生的重要原因之一[2]。阿帕替尼作為VEGFR-2抑制劑,是否通過調控生物鐘基因表達來抑制腫瘤血管生成,繼而達到抗腫瘤目的尚不清楚。本研究結果顯示,阿帕替尼組移植瘤組織Per3、CLOCK、CKIε、Cry2 mRNA相對表達量明顯低于對照組。表明阿帕替尼的抗腫瘤效果可能與抑制上述生物鐘基因表達有關。
綜上所述,阿帕替尼能抑制人肝癌裸鼠皮下移植瘤生長,其機制可能與下調腫瘤細胞生物鐘基因Per3、CLOCK、Cry2、CKIε mRNA表達有關。
參考文獻:
[1] Jemal A, Bray F, Center MM, et al. Global cancer statistics[J]. CA Cancer J Clin, 2011,61(2):69-90.
[2] 郭金虎,徐瓔,張二荃,等.生物鐘研究進展及重要前沿科學問題[J].中國科學基金,2014,28(3):179-186.
[3] Badiu C. Genetic clock of biologic rhythms[J]. J Cell Mol Med, 2003,71(4):408-416.
[4] Ko CH, Takahashi JS. Molecular components of the mammalian circadian clock[J]. Hum Mol Genet, 2006(15):R271-R277.
[5] 馮久桓,秦叔逵,王琳.甲磺酸阿帕替尼的研究現狀與進展[J].臨床腫瘤學雜志,2017,22(4):345-356.
[6] Li J, Qin S, Xu J, et al. Randomized, double-blind, placebo-controlled phase Ⅲ trial of apatinib in patients with chemotherapy-refractory advanced or metastatic adenocarcinoma of the stomach or gastroesophageal junction[J]. J Clin Oncol, 2016,34(13):1448-1454.
[7] Qin SK. Apatinib in Chinese patients with advanced hepatocellular carcinoma: a phase Ⅱ randomized, open-label trial[J]. J Clin Oncol, 2014,32(15):4019.
[8] Huang L, Wei Y, Shen S, et al. Therapeutic effect of apatinib on overall survival is mediated by prolonged progression-free survival in advanced gastric cancer patients[J]. Oncotarget, 2017,8(17):29346-29354.
[9] Yu EA, Weaver DR. Disrupting the circadian clock: gene-specific effects on aging, cancer, and other phenotypes[J]. Aging (Albany NY), 2011,3(5):479-493.
[10] Lin YM, Chang JH, Yeh KT, et al. Disturbance of circadian gene expression in hepatocellular carcinoma[J]. Mol Carcinog, 2008,47(12):925-933.
[11] Yang SL, Yu C, Jiang JX, et al. Hepatitis B virus X protein disrupts the balance of the expression of circadian rhythm genes in hepatocellular carcinoma[J]. Oncol Lett, 2014,8(6):2715-2720.
[12] Zhang H. Apatinib for molecular targeted therapy in tumor[J]. Drug Des Dev Ther, 2015(9):6075-6081.
[13] Kou P, Zhang Y, Shao W, et al. Significant efficacy and well safety of apatinib in an advanced liver cancer patient: a case report and literature review[J]. Oncotarget, 2017,8(12):20510-20515.
[14] Yang SL, Ren QG, Wen L, et al. Research progress on circadian clock genes in common abdominal malignant tumors[J]. Oncol Lett, 2017,14(5):5091-5098.
[15] Chen ST, Choo KB, Hou MF, et al. Deregulated expression of the PER1, PER2 and PER3 genes in breast cancers[J]. Carcinogenesis, 2005,26(7):1241-1246.
[16] Lee S, Donehower LA, Herron AJ, et al. Disrupting circadian homeostasis of sympathetic signaling promotes tumor development in mice[J]. PLoS One, 2010,5(6):e10995.
[17] Cao Q, Gery S, Dashti A, et al. A role for the clock gene per1 in prostate cancer[J]. Cancer Res, 2009,69(19):7619-7625.
[18] Miyazaki K, Wakabayashi M, Hara Y, et al. Tumor growth suppression in vivo by overexpression of the circadian component, PER2[J]. Genes Cells, 2010,15(4):351-358.
[19] Tian J, Tang ZY, Ye SL, et al. New human hepatocellular carcinoma (HCC) cell line with highly metastatic potential (MHCC97) and its expressions of the factors associated with metastasis[J]. Br J Cancer, 1999,81(5):814-821.
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
