沉默Ephrin B2基因表達對結直腸癌細胞侵襲和遷移的影響及其機制
2018-05-15 01:13:39張旭陳旺盛文坤明張夢嬌梁雪梅
山東醫藥 2018年8期
張旭,陳旺盛,文坤明,張夢嬌,梁雪梅
(1 西南醫科大學附屬醫院,四川瀘州646000;2 遵義醫學院附屬醫院)
促紅細胞生成素產生的肝細胞受體(EPH)家族是受體酪氨酸激酶家族中最大的亞族,在腫瘤發生、發展過程中具有重要作用[1]。EPH家族包括受體Eph和配體肝配蛋白(Ephrin)。肝配蛋白B2(Ephrin B2)是配體Ephrin的亞型之一,作為一種跨膜蛋白通過其胞外部分與相應受體Eph B4結合,激活Eph B4介導的正向信號通路,同時受體Eph B4也會使Ephrin B2胞內部分發生磷酸化,從而激活Ephrin B2介導的逆向信號通路,形成特有的雙向信號調節通路,在腫瘤侵襲和轉移過程中發揮雙重調節作用[2,3]。目前關于Ephrin B2介導的逆向信號通路在腫瘤中的作用仍存在爭議,多數研究傾向于經Ephrin B2介導的逆向信號通路與腫瘤侵襲和轉移有關[4~6],但其作用機制尚不清楚。近年研究證實,上皮間質轉化(EMT)在腫瘤侵襲和轉移過程中具有重要作用[7]。而Ephrin B2是否通過調控EMT過程參與腫瘤侵襲和轉移尚不十分清楚。2016年8月~2017年7月,本研究觀察了沉默Ephrin B2基因表達對結直腸癌細胞侵襲和遷移的影響,并探討其可能機制。
1 材料與方法
1.1 材料 人結直腸癌SW480細胞(以下稱SW480細胞),購自中國科學院上海細胞庫。Ephrin B2干擾質粒載體(Ephrin B2-siRNA)及對照質粒載體(NC-siRNA),由天津賽爾生物技術有限公司構建、篩選、包裝。所有引物序列由深圳華大基因科技服務有限公司設計合成。Transwell小室,美國Millipore公司;PCR逆轉錄試劑盒、TRIzol、SYBR?Premix Ex TaqTM,日本TaKaRa公司。兔抗人Ephrin B2、E-cadherin、N-cadherin、Vimentin一抗,美國Epitomics公司;兔抗人GAPDH一抗及HRP標記的山羊抗兔二抗,北京中杉金橋生物技術有限公司;BCA蛋白定量試劑盒、ECL發光試劑盒、SDS-PAGE凝膠配制試劑盒、Loading buffer、蛋白質Marker、Lipofectamine2000、結晶紫染液,上海碧云天生物技術有限公司。
1.2 沉默Ephrin B2基因表達及其效果驗證 將SW480細胞置于含10% FBS的RPMI 1640培養基中,37 ℃、5% CO2恒溫培養箱中常規培養。選擇對數生長期、生長狀態良好的細胞用于后續實驗。收集SW480細胞,0.25%胰酶消化,制成細胞懸液,以2×105個/孔接種于6孔板(內含10% FBS的RPMI 1640培養基),置于37 ℃、5% CO2恒溫培養箱中培養。隨機將細胞分為Ephrin B2組、陰性對照組、空白對照組,當細胞融合至70%~80%時,更換為無血清的RPMI 1640培養基,Ephrin B2組加入Ephrin B2-siRNA 1 μg+Lipofectamine2000 3 μL,陰性對照組加入NC-siRNA 1 μg+Lipofectamine2000 3 μL,空白對照組僅加入Lipofectamine2000 3 μL。各組轉染8 h,更換新的含10% FBS RPMI 1640培養基,繼續培養48 h,收集細胞。采用TRIzol法提取細胞總RNA,經蛋白核酸分析儀檢測,OD260/OD280為1.8~2.0。根據逆轉錄試劑盒說明將總RNA逆轉錄為cDNA。以cDNA為模板,按SYBR?Premix Ex TaqTM說明進行PCR擴增。引物序列:Ephrin B2上游引物5′-CGATTGAGCCTTACGACAC-3′,下游引物5′-TTTTAAGCGCTGAGCATTG-3′;內參GAPDH上游引物5′-TGACTTCAACAGCGACACCCA-3′,下游引物5′-CACCCTGTTGCTGTAGCCAAA-3′。PCR反應體系共25 μL:ddH2O 8.5 μL,5 pmol/μL Primer1 1 μL,5 pmol/μL Primer2 1 μL,cDNA 2 μL,SYBR Green Mix Taq 12.5 μL;反應條件:94 ℃ 30 s,94 ℃ 5 s、56 ℃ 30 s共40個循環,65 ℃ 5 s,72 ℃ 5 min。以GAPDH為內參,采用2-ΔΔCt法計算目的基因相對表達量。實驗重復3次,取平均值。結果顯示,Ephrin B2組、陰性對照組、空白對照組Ephrin B2 mRNA相對表達量分別為0.283±0.060、0.813±0.050、1.000±0.000。Ephrin B2組Ephrin B2 mRNA相對表達量明顯低于陰性對照組和空白對照組(P均<0.05),而陰性對照組與空白對照組比較P>0.05。證實Ephrin B2-siRNA能夠沉默Ephrin B2基因表達。
1.3 沉默Ephrin B2基因表達對SW480細胞侵襲能力影響的觀察 采用Transwell侵襲實驗。將Martrigel膠置于4 ℃冰箱中過夜解凍,以無血清的RPMI 1640培養基∶Martrigel膠=1∶8比例稀釋。取100 μL稀釋后的Martrigel膠置于Transwell小室底部,將小室置于37 ℃下風干。取對數生長期、生長狀態良好的SW480細胞,按1.2方法進行分組、轉染,轉染8 h,更換新的含10% FBS RPMI 1640培養基,繼續培養48 h,收集細胞,0.25%胰酶消化,制成密度為6×105個/mL的細胞懸液。取細胞懸液0.4 mL加入Transwell小室上室,下室加入含10% FBS RPMI 1640培養基0.6 mL,將Transwell小室置于37 ℃、5% CO2恒溫培養箱中培養。培養24 h,取出小室,吸盡上室培養基,PBS清洗,多聚甲醛固定20 min,自然風干后用0.1%結晶紫染色15 min,用棉簽擦去上室未穿透細胞及Martrigel膠。PBS沖洗干凈,自然風干,甘油封片,顯微鏡下計數穿膜細胞數,以此反映細胞侵襲能力。實驗重復3次,取平均值。
1.4 沉默Ephrin B2基因表達對SW480細胞遷移能力影響的觀察 采用劃痕修復實驗。選擇1.3中的細胞懸液1 mL接種于6孔板(內含10% FBS的RPMI 1640培養液),置于37 ℃、5% CO2恒溫培養箱中培養。待細胞貼壁后,用10 μL槍頭劃過細胞,用PBS洗盡漂浮的細胞;加入含1.5% FBS的RPMI 1640培養基,置于37 ℃、5% CO2恒溫培養箱中培養。顯微鏡下拍照觀察培養0、24 h細胞遷移情況,Image-Pro Plus軟件測量劃痕24 h細胞遷移距離。實驗重復3次,取平均值。
1.5 沉默Ephrin B2基因表達對SW480細胞E-cadherin、N-cadherin、Vimentin表達影響的觀察 ①E-cadherin、N-cadherin、Vimentin蛋白表達:采用Western blotting法。按1.2中方法進行分組、轉染。轉染8 h,更換新的含10% FBS的RPMI 1640培養基,繼續培養48 h,收集細胞。按全蛋白提取試劑盒說明提取細胞總蛋白,BCA法蛋白定量后,加入上樣緩沖液,100 ℃水浴10 min,使蛋白充分變性。取變性蛋白50 μg恒壓下行SDS-PAGE,按濕轉法將蛋白電泳產物轉印至PVDF膜。取出PVDF膜,5%脫脂奶粉封閉1 h,分別加入E-cadherin(1∶2 000)、N-cadherin(1∶2 000)、Vimentin(1∶5 000)、GAPDH(1∶5 000)一抗,4 ℃孵育過夜;TBST洗膜10 min×3次,加入HRP標記的山羊抗兔二抗(1∶5 000),室溫避光孵育1 h。ECL發光顯影,Bio-Rad凝膠成像系統掃描蛋白電泳條帶,Quantity One軟件分析各蛋白電泳條帶的灰度值。以目的蛋白電泳條帶灰度值/內參蛋白電泳條帶灰度值作為目的蛋白相對表達量。實驗重復3次,取平均值。②E-cadherin、N-cadherin、Vimentin mRNA表達:采用qRT-PCR法。具體步驟參照1.2。引物序列:E-cadherin上游引物5′-CTTCTCTCACGCTGTGTCATC-3′,下游引物5′-CTCCTGTGTTCCTGTTAATGGT-3′;N-cadherin上游引物5′-ACTCGGCTTAATGGTGAT-3′,下游引物5′-CACTGGCAAACCTTCACG-3′;Vimentin上游引物5′-AGGCAAAGCAGGAGTCCA-3′,下游引物5′-TATCAACCAGAGGGAGTG-3′。以GAPDH為內參,采用2-ΔΔCt法計算目的基因相對表達量。實驗重復3次,取平均值。

2 結果
2.1 各組細胞侵襲和遷移能力比較 見表1。
2.2 各組N-cadherin、Vimentin、E-cadherin蛋白和mRNA表達比較 見表2、3。
3 討論
結直腸癌是嚴重危及人民生命健康的惡性腫瘤,近年來雖然在全球范圍內結直腸癌的發病率及病死率有所下降,但由于我國人口基數大,每年的發病例數及死亡例數均居世界第1位,并呈年輕化趨勢[8]。雖然手術方式不斷改進、抗腫瘤新藥不斷涌現,但結直腸癌患者的總生存期仍不理想,尤其是中晚期結直腸癌患者。侵襲和轉移是導致結直腸癌患者預后不良的最重要原因,也是治療失敗導致患者死亡的重要原因。因此,探索結直腸癌侵襲和轉移的機制具有重要意義。
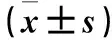
表1 各組細胞侵襲和遷移能力比較
注:與空白對照組比較,*P<0.05;與陰性對照組比較,#P<0.05。
表2 各組N-cadherin、Vimentin、E-cadherin蛋白相對表達量比較
注:與空白對照組比較,*P<0.05;與陰性對照組比較,#P<0.05。
表3 各組N-cadherin、Vimentin、E-cadherin mRNA相對表達量比較
注:與空白對照組比較,*P<0.05;與陰性對照組比較,#P<0.05。
近年來,EPH家族在腫瘤中的作用逐漸被重視。EPH家族包括Eph受體(Eph A1~Eph A10,Eph B1~Eph B6)和配體Ephrin(Ephrin A1~Ephrin A6,Ephrin B1~Ephrin B6)兩部分,其家族成員在腫瘤的發生、發展過程中具有重要作用[9]。Ephrin B2是一種跨膜的內嵌蛋白,由通過GPI與受體連接的胞外區、含PDZ結構域和酪氨酸激酶結合位點的胞內區組成。Ephrin B2主要與受體Eph B4、Eph B3、Eph B2結合。但Ephrin B2與Eph B4的親和力更高,可形成特有的雙向信號傳導通路,即由配體Ephrin B2為媒介作用于受體Eph B4使其發生磷酸化,從而激活下游信號傳導分子形成正向信號傳導通路,同時受體Eph B4介導激活配體Ephrin B2胞外區,使其發生磷酸化,繼而激活其下游信號傳導分子所形成的反向信號傳導通路。Ephrin B2與Eph B4形成的雙向信號通路在血管生成、神經發育、骨骼再生、細胞遷移等多種病理生理過程中發揮重要作用[10,11]。但目前關于Ephrin B2在腫瘤中的作用仍有爭議。有研究發現,Ephrin B2在食管癌、乳腺癌、卵巢癌等多種惡性腫瘤組織中高表達,并與腫瘤的侵襲和轉移有關[12,13]。而Depner等[14]研究發現,下調Ephrin B2表達可抑制某些腫瘤的侵襲和轉移。由此可見,Ephrin B2在腫瘤中的作用即可發揮癌基因作用,又能發揮抑癌基因作用。但其在結直腸癌細胞中的作用及其機制尚不十分清楚,仍需進一步研究。
普遍認為,EMT在腫瘤侵襲和轉移過程中發揮了至關重要的作用[15]。EMT可使具有極性的、相對穩定的上皮細胞通過一系列生化過程轉變成能在細胞基質間自由移動的間質細胞。EMT是一個非常復雜的分子過程,涉及多個信號通路,如TGF-β1通路、Hedgehog通路、Notch通路、Wnt通路等。各信號通路間彼此相互作用,激活一些轉錄因子,如鋅指轉錄因子Snail、堿性螺旋-環-螺旋轉錄因子Twist、鋅指E盒結合蛋白ZEB,共同參與導致上皮細胞標志分子E-cadherin下調及間質細胞標志分子N-cadherin、Vimentin表達上調。EMT可使腫瘤細胞喪失細胞極性和細胞彼此間的連接,使其侵襲和轉移能力增強[16]。但EMT并不是一成不變的,腫瘤細胞通過EMT獲得更強的侵襲和轉移能力,在到達一個新的生長環境后,又可通過一些轉錄因子的調節發生間質上皮轉化(MET),從而獲得具有上皮細胞表型和特性的細胞固定下來,繼而增殖形成新的病灶。眾多轉錄因子在腫瘤細胞發生EMT和MET過程中發揮了重要作用,有可能成為預測腫瘤侵襲和轉移的分子標志物。Ephrin B2是否通過調控腫瘤細胞的EMT過程發揮作用尚不清楚。
本研究通過RNA干擾技術沉默SW480細胞Ephrin B2基因表達,觀察沉默Ephrin B2基因表達對SW480細胞侵襲和遷移的影響。結果發現,Ephrin B2組細胞侵襲和轉移能力均明顯低于陰性對照組和空白對照組,而陰性對照組與空白對照組比較差異無統計學意義。說明沉默Ephrin B2基因表達能抑制SW480細胞侵襲和遷移。本研究結果還發現,Ephrin B2組E-cadherin蛋白和mRNA表達均明顯低于陰性對照組、空白對照組,N-cadherin、Vimentin蛋白和mRNA表達均明顯高于陰性對照組、空白對照組,而陰性對照組與空白對照組比較差異均無統計學意義。表明沉默Ephrin B2基因表達抑制SW480細胞侵襲和遷移的機制可能與調控EMT過程有關。
綜上所述,沉默Ephrin B2基因表達可抑制結直腸癌細胞侵襲和遷移,其機制可能與調控EMT過程有關。但其具體調控機制尚需進一步研究。
參考文獻:
[1] Li RX, Chen ZH, Chen ZK. The role of EPH receptors in cancer-related epithelial-mesenchymal transition[J]. Clin J Cancer, 2014,33(5):231-240.
[2] Djokovic D, Trindade A, Gigante J, et al. Combination of Dll4/Notch and EphrinB2/EphB4 targeted therapy is highly effective in disrupting tumor angiogenesis[J]. BMC Cancer, 2010(10):641-652.
[3] Arvanitis D, Davy A. Eph/ephrin signaling: networks[J]. Genes Dev, 2008,22(4):416-429.
[4] 張旭,陳秀,陳旺盛.結直腸癌組織Ephrin B2表達變化及其對上皮間質轉化的影響[J].山東醫藥,2017,57(36):65-68.
[5] Li X, Song C, Huang G, et al. The coexpression of EphB4 and EphrinB2 is associated with poor prognosis in HER2-positive breast cancer[J]. Onco Targets Ther, 2017(10):1735-1742.
[6] Alam SM, Fujimoto J, Jahan I, et al. Overexpression of ephrinB2 and EphB4 in tumor advancement of uterine endometrial cancers[J]. Ann Oncol, 2007,18(3):485-490.
[7] Chen S, Zhu J, Zuo S, et al. 1,25(OH)2D3attenuates TGF-β1/β2-induced increased migration and invasion via inhibiting epithelial-mesenchymaltransition in colon cancer cells[J]. Biochem Biophys Res Commun, 2015,468(1-2):130-135.
[8] Chen W, Zheng R, Baade PD, et al. Cancer statistics in China, 2015[J]. CA Cancer J Clin, 2016,66(2):115-132.
[9] Kandouz M. The Eph/Ephrin family in cancer metastasis: commu-nication at the service of invasion[J]. Cancer Metastasis Rev, 2012,31(1-2):353-373.
[10] Sturz A, Bader B, Thierauch KH, et al. EphB4 signaling is capable of mediating Ephrin B2-induced inhibition of cell migration[J]. Biochem Biophys Res Commun, 2004,313(1):80-88.
[11] Davies MH, Zamora DO, Smith JR, et al. Soluble Ephrin B2 mediates apoptosis in retinal neovascularization and in endothelial cells[J]. Microvasc Res, 2009,77(3):382-386.
[12] Adams RH, Wilkinson GA, Weiss C, et al. Roles of ephrinB ligands and EphB receptors in cardiovascular development: demarcation of arterial/venous domains, vascular morphogenesis, and sprouting angiogenesis[J]. Gene Dev, 1999,13(3):295-306.
[13] Zhang R, Zhao Y, Zhang S, et al. The expressions of EphrinB2 and VEGF in nasopharyngeal carcinoma and their clinical significance[J].Lin Chung Er Bi Yan Hou Tou Jing Wai Ke Za Zhi, 2013,27(4):178-180.
[14] Depner C, Zum Buttel H, Bogurcu N, et al. EphrinB2 repression through ZEB2 mediates tumour invasion and anti-angiogenic resistance[J]. Nat Commun, 2016(7):1-15.
[15] Yeung KT, Yang J. Epithelial-mesenchymal transition in tumor metastasis[J]. Mol Oncol, 2017,11(1):28-39.
[16] Smith BN, Bhowmick NA. Role of EMT in metastasis and therapy resistance[J]. J Clin Med, 2016,5(2):1-17.
