長鏈非編碼RNA FENDRR在子宮頸癌中的作用
2018-05-11 00:02:36蘭霄霄周志陽徐欣欣吳雪清
溫州醫科大學學報 2018年4期
蘭霄霄,周志陽,徐欣欣,吳雪清
(溫州醫科大學附屬第一醫院 婦產科,浙江 溫州 325015)
宮頸癌是女性生殖系統最常見的惡性腫瘤,雖然近年來宮頸癌疫苗和早期篩查的開展使得宮頸癌的發病率有所下降,但是宮頸癌仍然在女性癌癥相關性死亡中占據一定的比例[1]。FENDRR(FOXF1 adjacent non-coding developmental regulatory RNA)是位于第13號染色體的一段長鏈非編碼RNA(long non-coding RNA,lcnRNA),能與多梳蛋白復合物2(polycomb repressive complex 2,PRC2)或TrxG/MLL復合物結合[2],對小鼠心臟和體壁的正常發育具有重要作用[3]。已有研究表明FENDRR能通過影響纖連蛋白1的表達進而調控胃癌細胞的轉移[4],但目前尚未見關于其在宮頸癌中的研究。本研究通過在宮頸癌細胞中應用小干擾RNA(small interfering RNA,siRNA)介導的基因干擾技術特異性干擾FENDRR的表達,結合實時熒光定量PCR(quantitative real-time PCR,qRT-PCR)、細胞增殖CCK-8、Transwell以及裸鼠成瘤實驗,探討FENDRR在宮頸癌發病中的作用。
1 材料和方法
1.1 材料 宮頸癌細胞株(SiHa和HeLa,美國ATCC細胞庫),Advanced MEM培養基(美國Gibco公司),胎牛血清(FBS,美國Gibco公司),siRNA(上海吉瑪生物公司),轉染試劑(美國SignaGen公司),CCK-8試劑盒(日本Dojindo公司),Transwell小室、Matrigel基質膠(美國Corning公司),Trizol(美國Invitrogen公司),氯仿和異丙醇(美國Sigma公司),PCR引物(北京擎科生物技術公司)(見表1),反轉錄試劑盒(PrimeScript? RT reagent Kit)、qRTPCR試劑盒(SYBR?Premix Ex Taq?,日本Takara公司),裸鼠(上海斯萊克實驗動物有限責任公司),實驗動物使用許可證號:SYXK(浙)2015-0009。

表1 qRT-PCR引物的基因序列
1.2 方法
1.2.1 細胞培養:宮頸癌細胞株用含10% FBS、100 ng/mL鏈霉素、100 U/mL青霉素以及2 μmol/mL谷氨酸鈉的Advanced MEM細胞培養基于5% CO2、37 ℃條件下培養,細胞生長至80%~90%融合度時進行傳代。
1.2.2 siRNA轉染實驗:轉染前1 d接種適當數量的細胞至6孔板,使轉染時細胞融合度接近50%。轉染特異性干擾FENDRR的siRNA為實驗組,轉染空白對照siRNA為對照組。具體轉染步驟如下:①轉染前30 min在上述鋪好細胞的6孔板中換上1 mL新鮮細胞培養基;②用100 μL稀釋好的1×轉染緩沖液溶解5 μL siRNA,混勻后再加入4 μL轉染試劑,充分混勻后室溫孵育15 min;③將配好的含有siRNA的試劑加入6孔板的相應孔中,輕輕混勻;④培養6 h后更換新鮮培養基,繼續將6孔板置于5% CO2、37 ℃培養箱中培養;⑤轉染24~48 h后進行siRNA干擾沉默效果及細胞功能檢測。
1.2.3 qRT-PCR:①提取細胞總RNA:運用Trizol一步法提取細胞總RNA。測定RNA濃度及OD260/OD280在1.8~2.0之間,符合要求。RNA保存于-80 ℃冰箱用于后續實驗。②反轉錄:每組各取1000 ng總RNA按照反轉錄試劑盒說明書進行操作,反轉錄成的cDNA置于-20 ℃冰箱保存。③qRT-PCR:利用qRT-PCR試劑盒對反轉錄的cDNA進行PCR擴增,具體擴增體系為:SYBR?Premix Ex Taq II(2×)5 μL,ddH2O 3.4 μL,上下游引物混合液0.4 μL,ROX Reference Dye II 0.2 μL,cDNA 1 μL。
1.2.4 CCK-8實驗:96孔板每孔種植SiHa細胞2000個,HeLa細胞2500個,分別于細胞貼壁后的0、24、48、72、96 h向每孔加入10 μL CCK-8試劑,37 ℃孵育3 h后用酶標儀檢測每孔450 nm波長的吸收值。
1.2.5 Transwell實驗:①細胞遷移實驗:Transwell小室的下室中加入500 μL含10% FBS的完全細胞培養基,上室接種300 μL不含FBS的細胞懸液(內含細胞25000個),37 ℃培養24 h后取出小室,棄上室殘存培養基,待自然涼干后用甲醇固定10 min,0.1%結晶紫染色10 min,用棉簽擦去小室內表面殘留的細胞及染料,顯微鏡觀察穿過小室濾膜的細胞并計數(10倍鏡下隨機選擇5個視野);②細胞侵襲實驗:將存于-20 ℃的Matrigel置于4 ℃解凍,用預冷的無FBS培養基按1∶30稀釋Matrigel,在小室的上室中均勻鋪上100 μL稀釋好的Matrigel,37 ℃放置約2 h使其凝結成膠體。后續實驗步驟同細胞遷移實驗。以上實驗分別至少重復3次。
1.2.6 裸鼠成瘤實驗:2~3周的雌性裸鼠,每組5只。收集指數增長期的細胞用無FBS的培養基和Matrigel按1∶1的比例制備細胞懸液,在每只裸鼠的腋下皮下種植含2.5×106個HeLa細胞的細胞懸液100 μL(左側種植對照組細胞,右側種植實驗組細胞)。種植細胞1周后,每3 d檢測1次腫瘤的大小和裸鼠的體質量,4周后取出腫瘤,測量腫瘤質量并拍照記錄。
1.3 統計學處理方法 應用GraphPad Prism 5和SPSS12.0軟件進行統計學分析。計量資料采用±s表示,組間比較采用獨立樣本t檢驗。P<0.05為差異有統計學意義。
2 結果
2.1 轉染siRNA后FENDRR的表達 qRT-PCR結果顯示,轉染了特異性靶向FENDRR的siRNA后,宮頸癌細胞中FENDRR的表達水平明顯下降,在SiHa細胞中FENDRR相對表達水平由(1.013±0.120)降為(0.226±0.024),在HeLa細胞中FENDRR相對表達水平由(1.001±0.025)降為(0.099±0.010),差異均有統計學意義(P<0.01)。
2.2 FENDRR抑制宮頸癌細胞體外增殖 CCK-8實驗顯示,干擾FENDRR的表達后,宮頸癌細胞株SiHa和HeLa的增殖能力明顯增強,SiHa細胞在48、72和96 h實驗組和對照組的OD分別為: (0.932±0.014)、(1.603±0.012)、 (2.282±0.024)和(0.780±0.005)、 (1.150±0.011)、 (1.693±0.019),差異有統計學意義(P<0.01),見圖1A。HeLa細胞在72和96 h實驗組和對照組的OD分別為: (1.175±0.017)、(1.983±0.021)和(0.889±0.014)、 (1.483±0.017),差異均有統計學意義(P<0.01),見圖1B。
2.3 FENDRR抑制宮頸癌細胞體外運動能力 為了驗證FENDRR對宮頸癌細胞體外遷移以及侵襲能力的影響,應用Transwell小室實驗檢測各組細胞的遷移和侵襲能力。用Leica熒光正置顯微鏡進行拍照,結果顯示,干擾FENDRR后,宮頸癌細胞系的遷移和侵襲能力明顯增強(見圖2A-B)。SiHa遷移細胞數從(95.2±2.9)增至(212.4±7.4),HeLa遷移細胞數從(130.2±6.2)增至(433.8±13.1),差異均有統計學意義(P<0.01)。SiHa侵襲細胞數從(39.6±1.9)增至(97.2±7.2),HeLa侵襲細胞數從(109.2±4.6)增至(252.4±18.2),差異均有統計學意義(P<0.01)。
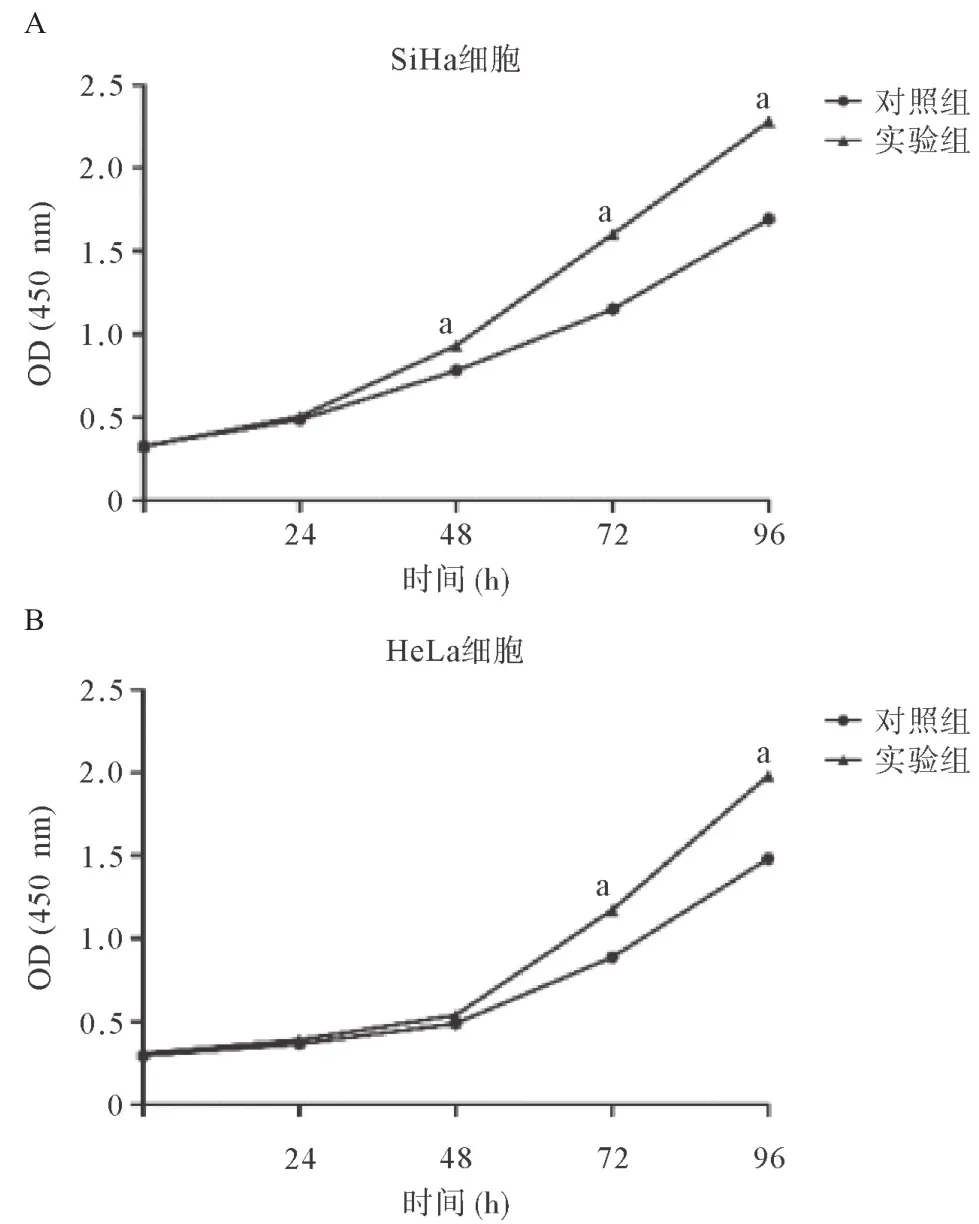
圖1 FENDRR抑制宮頸癌細胞SiHa(A)和HeLa(B)體外增殖
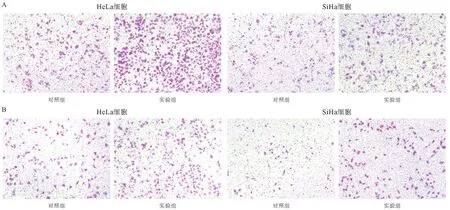
圖2 FENDRR抑制宮頸癌細胞遷移能力(A)和侵襲能力(B)(×10)
2.4 FENDRR抑制宮頸癌細胞體內增殖 為了進一步驗證FENDRR在宮頸癌中的作用,采用HeLa細胞進行了裸鼠體內成瘤實驗。連續監測腫瘤大小和裸鼠體質量,根據體積計算公式(體積=1/2長×寬2),發現干擾FENDRR后HeLa細胞的成瘤能力增強,腫瘤體積與對照組比從(853.2±59.4)mm3增加到(1155.0±51.5)mm3,差異有統計學意義(P<0.01),見圖3A。腫瘤質量從(0.544±0.053)g增加到(0.814±0.061)g,差異有統計學意義(P<0.05),見圖3B。
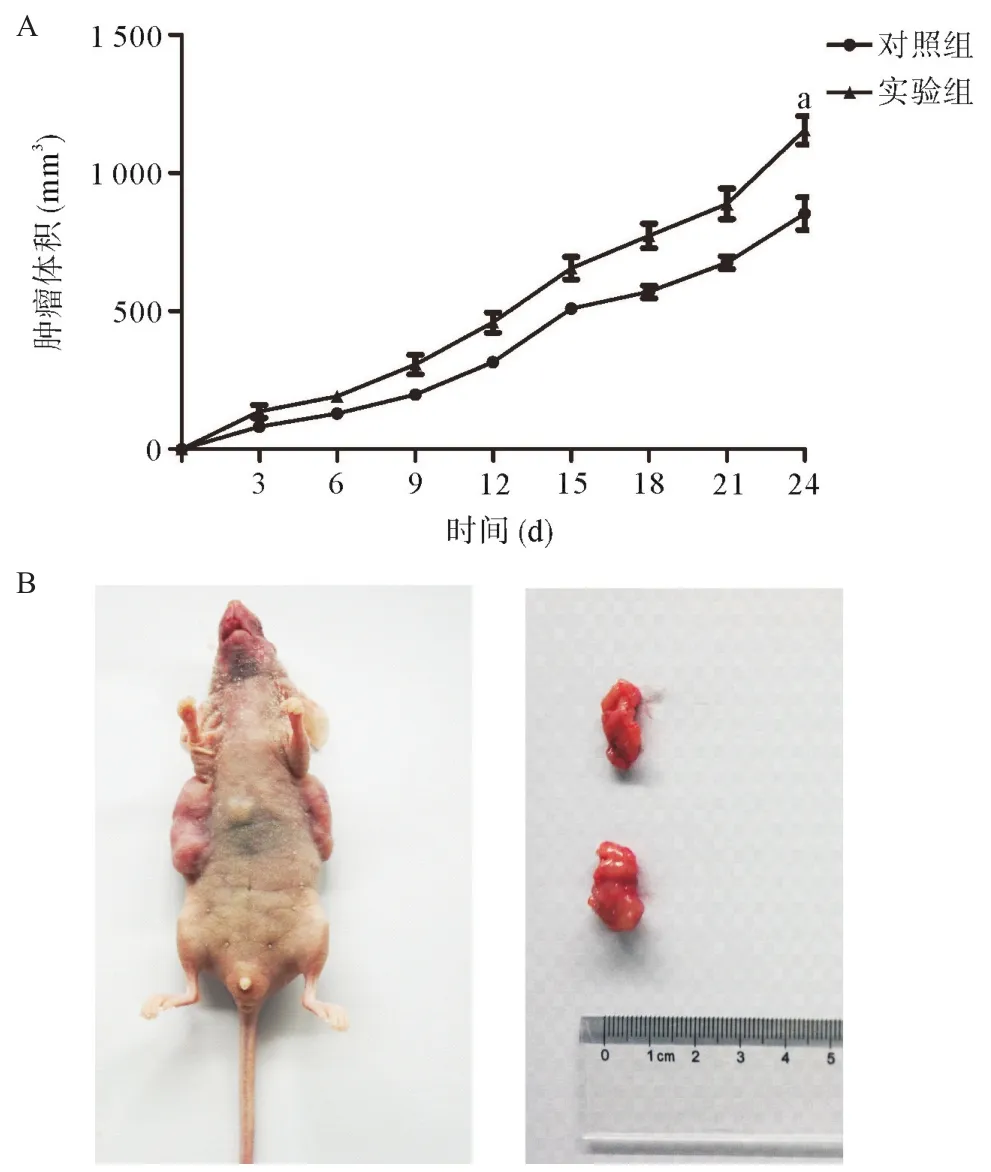
圖3 FENDRR抑制宮頸癌細胞體內增殖
3 討論
隨著基因芯片和高通量測序等生物信息學技術的發展,我們逐漸認識到在人類全基因組以及轉錄組中蛋白編碼基因僅占了不到2%,超過75%的基因不具備蛋白編碼功能,但后者在各種生物學過程中同樣發揮著重要的作用[5]。LncRNA是長度大于200個核苷酸并且不具備蛋白編碼功能的RNA,占據非編碼RNA的主要部分。近幾年,越來越多的研究發現lncRNA參與許多生物學進程[6-7]。在癌癥中,lncRNA的表達具有一定的組織或者腫瘤特異性,并且能與其他生物分子發生相互作用影響細胞周期調節、細胞生存、免疫反應以及細胞干性等過程,進而影響癌癥細胞的表型轉化[8]。部分lncRNA也受一些關鍵抑癌基因或者促癌基因的轉錄調節[9-10]。隨著研究的進一步深入,我們發現lncRNA能作為一種信號分子反映特異性細胞狀態以及作為區分癌癥等細胞病理狀態的標志物[11],為癌癥提供一定的預后價值,甚至還可以作為一種新的治療選擇。BUSSEMAKERS等[12]對前列腺癌組織和正常組織進行比較分析發現了首個與癌癥相關的異常表達的lncRNA-PCA3(prostate cancer associated 3,PCA3),PCA3在前列腺癌組織中明顯高表達并且具有很強的組織特異性,在前列腺癌的進展中具有重要的作用。目前PCA3已經成為臨床上前列腺癌診斷的重要生物學標志物,并且與血清前列腺特異性抗原(PSA)檢測相比具有非侵襲性、高特異性等優點[13]。
FENDRR基因是2009年最先鑒定,定位于染色體3q13.31,毗鄰FOXF1,有4個外顯子,編碼一個長3099個核苷酸的lncRNA[2]。目前對FENDRR的研究較少,其表達和功能異常與腫瘤相關的報道更是罕見,其在子宮頸癌中的作用也尚不清楚。XU等[4]研究發現,與正常組織和細胞相比,FENDRR在胃癌組織和細胞系中的表達均呈現顯著下調,并且其低表達與胃癌的高分期、深侵襲和高轉移相關。進一步研究發現FENDRR通過降低纖維連接蛋白1和MMP2/MMP9的表達進而增強胃癌的遷移和侵襲能力,提示FENDRR在胃癌中具有一定的預后預測價值,并且還可能成為治療的新靶點。MIAO等[14]對非小細胞肺癌(nonsmall cell lung cancer,NSCLC)及其正常對照組織進行基因芯片分析發現FENDRR在NSCLC中明顯低表達,而過表達FENDRR使得肺癌細胞的遷移和侵襲能力下降。此外,FENDRR還能誘導肺癌細胞的凋亡[15]。為了探索FENDRR對宮頸癌細胞生物學特性的影響,本研究通過siRNA介導的基因干擾技術特異性干擾FENDRR在宮頸癌細胞中的表達,結合細胞增殖和Transwell實驗,發現干擾FENDRR后宮頸癌細胞的體外增殖和遷移、侵襲能力均有所增強。此外,本研究還通過裸鼠成瘤實驗進一步證實了干擾FENDRR能增強宮頸癌細胞的體內成瘤能力,說明FENDRR在宮頸癌中確實發揮了一定的抑癌作用,但具體機制目前還不清楚。PRC2是一種甲基轉移酶,能甲基化H3K27以抑制特異性基因的轉錄,在染色質結構修飾和基因活性調節中具有重要作用[16-18]。研究表明,許多lncRNA功能的發揮都在一定程度上依賴于其與染色質修飾復合物PRC2之間的相互聯系,其中研究得比較深入的是lncRNA HOTAIR。GUPTA等[19]通過對正常人類乳腺上皮細胞、原發性乳腺癌以及遠處轉移組織進行生物信息學分析,發現HOTAIR在原發性乳腺癌和轉移性乳腺癌中表達水平增高,并且HOTAIR高表達與乳腺癌患者后期的轉移和死亡密切相關,進一步研究發現HOTAIR主要通過結合PRC2發揮促進腫瘤侵襲和轉移作用。同樣地,近期研究發現FENDRR能夠結合PRC2或TrxG/MLL復合體,后兩者均為染色質修飾復合物,能促進靶基因啟動子的甲基化,沉默靶基因的表達進而發揮相應的作用[3,20],還有研究表明FENDRR可能通過與PRC2復合物中的EZH2發生相互作用進而調節肺癌細胞的上皮-間質轉化、多能性以及轉移功能[14]。因此,我們推測FENDRR可能也是通過與染色質修飾復合物PRC2結合,沉默相應的靶基因,進而改變宮頸癌細胞的表型,但具體的調節機制還有待于進一步研究。
綜上,本研究發現FENDRR對宮頸癌細胞的生物學特性具有抑制作用,干擾FENDRR的表達能增強宮頸癌細胞的體內外增殖能力和體外遷移、侵襲潛能,說明FENDRR可能在宮頸癌的進展中具有重要的抑癌作用。結合目前關于lncRNA的研究,我們推測FENDRR有可能成為一個新的生物標志物,為將來宮頸癌的診斷和治療提供新的思路。
參考文獻:
[1] TORRE L A, BRAY F, SIEGEL R L, et al. Global cancer statistics, 2012[J]. CA Cancer J Clin, 2015, 65(2): 87-108.
[2] KHALIL A M, GUTTMAN M, HUARTE M, et al. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression[J]. Proc Natl Acad Sci U S A, 2009, 106(28): 11667-11672.
[3] GROTE P, WITTLER L, HENDRIX D, et al. The tissuespecific lncRNA Fendrr is an essential regulator of heart and body wall development in the mouse[J]. Dev Cell, 2013,24(2): 206-214.
[4] XU T P, HUANG M D, XIA R, et al. Decreased expression of the long non-coding RNA FENDRR is associated with poor prognosis in gastric cancer and FENDRR regulates gastric cancer cell metastasis by affecting fibronectin1 expression[J]. J Hematol Oncol, 2014, 7: 63.
[5] DJEBALI S, DAVIS C A, MERKEL A, et al. Landscape of transcription in human cells[J]. Nature, 2012, 489(7414):101-108.
[6] PRENSNER J R, CHINNAIYAN A M. The emergence of lncRNAs in cancer biology[J]. Cancer Discov, 2011, 1(5):391-407.
[7] 孫祥威, 胡盈盈, 徐劍峰, 等. 長鏈非編碼RNA AL049452在直腸癌組織中的表達及臨床意義[J]. 溫州醫科大學學報, 2017, 47(8): 566-570.
[8] GUTTMAN M, AMIT I, GARBER M, et al. Chromatin signature reveals over a thousand highly conserved large noncoding RNAs in mammals[J]. Nature, 2009, 458(7235):223-227.
[9] HUARTE M, GUTTMAN M, FELDSER D, et al. A large intergenic noncoding RNA induced by p53 mediates global gene repression in the p53 response[J]. Cell, 2010, 142(3):409-419.
[10] ZHENG G X, DO B T, WEBSTER D E, et al. Dicer-microRNA-Myc circuit promotes transcription of hundreds of long noncoding RNAs[J]. Nat Struct Mol Biol, 2014, 21(7):585-590.
[11] WANG K C, CHANG H Y. Molecular mechanisms of long noncoding RNAs[J]. Mol Cell, 2011, 43(6): 904-914.
[12] BUSSEMAKERS M J, VAN BOKHOVEN A, VERHAEGH G W, et al. DD3: a new prostate-specific gene, highly overexpressed in prostate cancer[J]. Cancer Res, 1999, 59(23):5975-5979.
[13] WEI J T, FENG Z, PARTIN A W, et al. Can urinary PCA3 supplement PSA in the early detection of prostate cancer?[J].J Clin Oncol, 2014, 32(36): 4066-4072.
[14] MIAO L, HUANG Z, ZENGLI Z, et al. Loss of long noncoding RNA FOXF1-AS1 regulates epithelial-mesenchymal transition, stemness and metastasis of non-small cell lung cancer cells[J]. Oncotarget, 2016, 7(42): 68339-68349.
[15] 徐然, 尚超, 石文君. 長鏈非編碼RNA FENDRR對非小細胞肺癌細胞增殖及凋亡的影響[J]. 現代腫瘤醫學, 2016,24(17): 2667-2669.
[16] BRACKEN A P, DIETRICH N, PASINI D, et al. Genomewide mapping of Polycomb target genes unravels their roles in cell fate transitions[J]. Genes Dev, 2006, 20(9): 1123-1136.
[17] KU M, KOCHE R P, RHEINBAY E, et al. Genomewide analysis of PRC1 and PRC2 occupancy identifies two classes of bivalent domains[J]. PLoS Genet, 2008, 4(10):e1000242.
[18] SCHUETTENGRUBER B, CHOURROUT D, VERVOORT M, et al. Genome regulation by polycomb and trithorax proteins[J]. Cell, 2007, 128(4): 735-745.
[19] GUPTA R A, SHAH N, WANG K C, et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis[J]. Nature, 2010, 464(7291): 1071-1076.
[20] GROTE P, HERRMANN B G. The long non-coding RNA FENDRR links epigenetic control mechanisms to gene regulatory networks in mammalian embryogenesis[J]. RNA Biol,2013, 10(10): 1579-1585.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
發明與創新(2016年38期)2016-08-22 03:02:52
