IL-17A對(duì)人永生化角質(zhì)形成細(xì)胞角蛋白17表達(dá)及STAT3信號(hào)通路的影響
2018-04-11 05:02:49盧永申魏明
山東醫(yī)藥 2018年10期
盧永申,魏明
(鄭州大學(xué)第五附屬醫(yī)院,鄭州450052)
銀屑病是一種由T細(xì)胞介導(dǎo)的持續(xù)存在的慢性、炎癥性疾病,其主要表現(xiàn)是角質(zhì)形成細(xì)胞(KC)的異常分化和增殖[1,2]。Th17細(xì)胞是新型的CD4+T細(xì)胞,其標(biāo)志性細(xì)胞因子白細(xì)胞介素-17A(IL-17A)具有強(qiáng)大的致炎性,在銀屑病的發(fā)生發(fā)展過(guò)程中發(fā)揮重要作用[3,4]。角蛋白也是上皮組織增生分化狀態(tài)的分子標(biāo)志,不同類型的上皮組織或同一類型的上皮組織處于增生分化的不同階段,或經(jīng)歷異常的增生分化時(shí),角蛋白的表達(dá)模式有顯著差異。其中,角蛋白17(K17)是表皮細(xì)胞增生分化的重要標(biāo)志分子。大量研究表明,銀屑病的發(fā)病機(jī)制與K17在T淋巴細(xì)胞介導(dǎo)下的免疫反應(yīng)密切相關(guān)[5~7]。信號(hào)轉(zhuǎn)導(dǎo)和轉(zhuǎn)錄激活因子(STAT)信號(hào)通路是細(xì)胞因子發(fā)揮其生物效應(yīng)最普遍且最重要的細(xì)胞內(nèi)信號(hào)傳導(dǎo)通路,其中STAT3廣泛表達(dá)于不同的細(xì)胞和組織。活化的STAT3與啟動(dòng)子結(jié)合,啟動(dòng)下游靶基因的表達(dá),從而影響細(xì)胞周期或抑制細(xì)胞凋亡而參與病變的發(fā)生發(fā)展。已有研究表明,STAT3的活化與銀屑病角質(zhì)形成細(xì)胞的異常增生有關(guān)。2016年5月,我們通過(guò)體外培養(yǎng)人永生化角質(zhì)形成細(xì)胞HaCaT,觀察IL-17A對(duì)細(xì)胞K17及STAT3信號(hào)通路的影響,為探討銀屑病的發(fā)病機(jī)制和靶向治療提供理論依據(jù)。
1 材料與方法
1.1 材料
HaCaT購(gòu)于南京凱基生物有限公司,TRIzol購(gòu)于美國(guó)Invitrogen公司,重組IL-17A購(gòu)于美國(guó)PeproTech公司,改良Eagle培養(yǎng)基(DMEM)、胎牛血清、0.25%胰酶/EDTA、青霉素/鏈霉素雙抗溶液、二甲基亞砜(DMSO)、RPMI1640培養(yǎng)液購(gòu)于美國(guó)Gibco公司;5-溴-4-氯-3-吲哚基-磷酸鹽(BCIP)/四唑硝基藍(lán)(NBT)顯色試劑盒購(gòu)于瑞士Roche公司,四甲基偶氮唑鹽(MTT)試劑盒購(gòu)于美國(guó)Sigma公司,逆轉(zhuǎn)錄PCR試劑盒購(gòu)于大連寶生物工程有限公司,異硫氰酸熒光素(FITC)標(biāo)記的Annexin Ⅴ細(xì)胞凋亡試劑購(gòu)于美國(guó)BD公司,流式細(xì)胞儀檢測(cè)相關(guān)試劑購(gòu)于美國(guó)BD公司,細(xì)胞培養(yǎng)箱為美國(guó)Thermo公司產(chǎn)品。
1.2 細(xì)胞培養(yǎng)
向細(xì)胞中加入含10%胎牛血清及1%青鏈霉素的改良Eagle培養(yǎng)基(DMEM),37 ℃、5% CO2條件下培養(yǎng)。待細(xì)胞單層鋪滿培養(yǎng)瓶后,PBS漂洗1次,加入0.25%胰蛋白酶消化液,待鏡下細(xì)胞回縮變圓時(shí),加入DMEM培養(yǎng)液終止消化。收集細(xì)胞,離心棄上清,將細(xì)胞懸液分別移至冷凍管中,每管1.5 mL。將凍存管先置于4 ℃冰箱2 h,再移至-80 ℃低溫冰箱24 h,然后投入-196 ℃液氮中保存?zhèn)溆谩?/p>
1.3 細(xì)胞分組與處理
將細(xì)胞重懸于細(xì)胞培養(yǎng)液中,按5×105/mL接種于6孔板,加入含10%胎牛血清的RPMI1640培養(yǎng)液培養(yǎng)24 h。待細(xì)胞達(dá)到70%~80%融合度時(shí),將細(xì)胞分為三組。空白對(duì)照組僅加DMEM高糖培養(yǎng)基,誘導(dǎo)組加入含50 μg/L IL-17A的DMEM高糖培養(yǎng)基,抑制劑組加入含50 μg/L IL-17A的DMEM高糖培養(yǎng)基和10 μmol/L STAT3抑制劑Piceatannol。孵育6 h后,置換為含10%胎牛血清的RPMI1640培養(yǎng)液,根據(jù)實(shí)驗(yàn)要求繼續(xù)培養(yǎng)。
1.4 細(xì)胞增殖能力觀察
采用MTT比色法。細(xì)胞分組培養(yǎng)12、24、36、48 h時(shí),分別檢測(cè)細(xì)胞增殖情況。加入胰酶消化細(xì)胞制備單細(xì)胞懸液,按6×104/孔接種于96孔培養(yǎng)板,細(xì)胞融合達(dá)到80%左右時(shí)開始檢測(cè)。棄原培養(yǎng)基,每孔加入5 mg/mL MTT 20 μL,37 ℃孵育4 h。棄上清,每孔加入150 μL DMSO,震蕩10 min充分溶解藍(lán)色結(jié)晶,于490 nm波長(zhǎng)處檢測(cè)吸光度A值。
1.5 細(xì)胞凋亡檢測(cè)
采用流式細(xì)胞術(shù)。細(xì)胞分組培養(yǎng)48 h后,加入不含EDTA的胰蛋白酶消化,PBS洗滌,離心棄上清,加入緩沖液100 μL重懸細(xì)胞,隨后加入Annexin Ⅴ-FITC 5 μL和溴化乙錠(PI)5 μL,室溫避光孵育15 min。加入500 μL緩沖液,流式細(xì)胞儀檢測(cè)細(xì)胞凋亡率。
1.6 細(xì)胞K17 mRNA表達(dá)檢測(cè)
采用RT-PCR方法。提取培養(yǎng)48 h后的各組細(xì)胞總RNA,逆轉(zhuǎn)錄合成cDNA。目的基因K17上游引物5′-ATGGATCCATGACCATGCAGGCCTTGGAGA-3′,下游引物5′-AGGAATTCTCACGTCTTCACATCCAGCAGGA-3′;β-actin上游引物5′-CACGATGGAGGGGCCGGACTCATC-3′,下游引物5′-TAAAGACCTCTATGCCAACACAGT-3′。PCR反應(yīng)體系為25 μL,擴(kuò)增條件為:94 ℃預(yù)變性3 min;94 ℃變性30 s,57 ℃退火30 s,72 ℃延伸50 s,共35個(gè)循環(huán);最后72 ℃延伸5 min。擴(kuò)增產(chǎn)物經(jīng)2%瓊脂糖凝膠電泳,凝膠成像系統(tǒng)掃描拍照,計(jì)算各電泳條帶灰度值與β-actin條帶灰度值的比值,即為目的基因的相對(duì)表達(dá)量。
1.7 細(xì)胞K17、p-STAT3蛋白表達(dá)檢測(cè)
采用Western blotting法。收集培養(yǎng)48 h后的各組細(xì)胞,提取總蛋白,BCA法檢測(cè)蛋白濃度。上樣40 μg,電泳、轉(zhuǎn)膜及封閉;一抗孵育(兔抗人K17、p-STAT3多克隆抗體滴度為1∶1 000,鼠抗人β-actin單克隆抗體為1∶1 000)4 ℃過(guò)夜,二抗孵育(山羊抗兔IgG抗體滴度為1∶2 000,馬抗小鼠IgG抗體為1∶2 000)2 h;BCIP/NBT顯色液顯色,凝膠成像系統(tǒng)觀察結(jié)果。以目的蛋白條帶與相應(yīng)內(nèi)參β-actin條帶的灰度值比值作為其蛋白水平的相對(duì)表達(dá)量。
1.8 統(tǒng)計(jì)學(xué)方法
2 結(jié)果
2.1 各組細(xì)胞增殖能力比較
見(jiàn)表1。隨著培養(yǎng)時(shí)間的延長(zhǎng),各組細(xì)胞增殖能力均增加,作用36、48 h時(shí)高于作用12、24 h時(shí)(P均<0.01)。誘導(dǎo)組各時(shí)間點(diǎn)HaCaT細(xì)胞增殖能力均較空白對(duì)照組、抑制劑組升高(P均<0.01),空白對(duì)照組與抑制劑組比較差異無(wú)統(tǒng)計(jì)學(xué)意義(P均>0.05)。
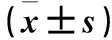
表1 各組細(xì)胞增殖能力比較
注:與空白對(duì)照組比較,*P<0.01;與抑制劑組比較,#P<0.01;與同組作用12、24 h比較,△P<0.01。
2.2 各組細(xì)胞凋亡率比較
空白對(duì)照組、誘導(dǎo)組、抑制劑組的細(xì)胞凋亡率分別為15.34%±1.32%、5.96%±0.68%、15.62%±1.26%,誘導(dǎo)組細(xì)胞凋亡率較空白對(duì)照組和抑制劑組降低(P均<0.05),空白對(duì)照組與抑制劑組比較差異無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05)。
2.3 各組K17 mRNA表達(dá)比較
空白對(duì)照組、誘導(dǎo)組、抑制劑組K17 mRNA的相對(duì)表達(dá)量分別為0.16±0.08、0.31±0.12、0.14±0.09,誘導(dǎo)組K17 mRNA表達(dá)高于空白對(duì)照組和抑制劑組(P均<0.05),空白對(duì)照組與抑制劑組比較差異無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05)。
2.4 各組K17、p-STAT3蛋白表達(dá)比較
空白對(duì)照組、誘導(dǎo)組和抑制劑組K17蛋白相對(duì)表達(dá)量分別為0.49±0.23、0.98±0.31、0.46±0.25,p-STAT3蛋白相對(duì)表達(dá)量分別為0.71±0.28、1.42±0.39、0.68±0.31,誘導(dǎo)組K17、p-STAT3蛋白表達(dá)水平均高于空白對(duì)照組和抑制劑組(P均<0.05),空白對(duì)照組與抑制劑組比較差異無(wú)統(tǒng)計(jì)學(xué)意義(P均>0.05)。
3 討論
角蛋白是角質(zhì)細(xì)胞中的主要骨架蛋白,多于上皮細(xì)胞中表達(dá),是上皮細(xì)胞生長(zhǎng)、分化及成熟的重要標(biāo)志物[8]。研究表明,在銀屑病患者皮損中,KC所表達(dá)的角蛋白譜會(huì)發(fā)生改變,K17水平顯著升高。K17作為銀屑病增殖相關(guān)角蛋白,在正常皮膚中不表達(dá)或表達(dá)量很低,而在銀屑病患者的皮損中呈高表達(dá),同時(shí)其表達(dá)水平與銀屑病發(fā)病過(guò)程和嚴(yán)重程度有一定相關(guān)性[9,10]。所以,K17與銀屑病的關(guān)系非常密切,在銀屑病的發(fā)生發(fā)展中具有重要作用。
文獻(xiàn)[11]報(bào)道,來(lái)源于Th17、Th22細(xì)胞的IL-17A和IL-22在角質(zhì)形成細(xì)胞以劑量相關(guān)性方式上調(diào)K17 mRNA及蛋白表達(dá),這些效應(yīng)被STAT-1和STAT-3特異抑制劑小干擾RNA部分阻斷。該報(bào)道還提出存在K17-T細(xì)胞-細(xì)胞因子免疫環(huán),其中異位表達(dá)K17通過(guò)激活自身反應(yīng)性T細(xì)胞影響銀屑病[12]。應(yīng)用反義寡核苷酸和RNA干擾抑制K17 mRNA和蛋白在銀屑病皮膚組織中的表達(dá),在臨床和組織學(xué)上對(duì)銀屑病均有改善作用[12]。也有研究表明,K17攜帶某些和鏈球菌M6蛋白相似的表位[13],這些表位可以激活T細(xì)胞(如Th1細(xì)胞)增殖。活化的T細(xì)胞產(chǎn)生銀屑病相關(guān)細(xì)胞因子γ干擾素(IFN-γ),細(xì)胞因子IFN-γ又可以依賴STAT1和STAT3信號(hào)通路激活KC表達(dá)K17,繼而形成一個(gè)反應(yīng)環(huán)路,再次激活T細(xì)胞增殖和銀屑病相關(guān)細(xì)胞因子的產(chǎn)生[14]。
Th17細(xì)胞在銀屑病的形成中發(fā)揮關(guān)鍵作用[15],其分泌的IL-17A通過(guò)多種途徑影響角蛋白細(xì)胞[16]。有關(guān)IL-17A激活KC中STAT3信號(hào)通路未見(jiàn)文獻(xiàn)報(bào)道,為此我們對(duì)IL-17A調(diào)控KC中K17表達(dá)的具體機(jī)制進(jìn)行探討。我們預(yù)先用STAT3信號(hào)通路抑制劑Piceatannol處理KC,再用IL-17A進(jìn)行刺激,發(fā)現(xiàn)與誘導(dǎo)組比較,抑制劑組K17 mRNA和蛋白表達(dá)均下降,表明IL-17A調(diào)控K17的表達(dá)可能是通過(guò)激活JAK-STAT3這條信號(hào)轉(zhuǎn)導(dǎo)通路實(shí)現(xiàn)的。這個(gè)結(jié)果與STAT3在銀屑病患者皮損KC中特異性高表達(dá)是一致的[17]。其調(diào)控機(jī)制是IL-17A通過(guò)與KC表面受體結(jié)合后激活STAT3來(lái)上調(diào)K17的表達(dá)。IL-17A可在角蛋白水平上參與促進(jìn)銀屑病的發(fā)生發(fā)展,進(jìn)一步肯定了Th細(xì)胞及其細(xì)胞因子在銀屑病中的作用,為銀屑病的發(fā)病機(jī)制研究和將來(lái)對(duì)細(xì)胞因子、信號(hào)通路的抗銀屑病臨床治療提供了新的靶點(diǎn)和思路。
綜上所述,K17在T細(xì)胞免疫調(diào)節(jié)下,通過(guò)STAT3信號(hào)通路調(diào)節(jié)細(xì)胞增殖和凋亡,活化相關(guān)生長(zhǎng)因子,在促進(jìn)銀屑病發(fā)生發(fā)展過(guò)程中具有重要作用。
參考文獻(xiàn):
[1] 蔣正強(qiáng),滿孝勇,鄭敏.角蛋白17在增殖性皮膚病中的表達(dá)[J].中華皮膚科雜志,2013,46(1):45-46.
[2] 白明輝,欒莉,趙曄.重度尋常型銀屑病患者外周血CD8+T淋巴細(xì)胞CCR6及上清液中IL-22、IL-17的表達(dá)[J].中國(guó)中西醫(yī)結(jié)合皮膚性病學(xué)雜志,2015,14(4):228-230.
[3] Diani M, Altomare G, Reali E. T cell responses in psoriasis and psoriatic arthritis[J]. Autoimmun Rev, 2015,14(4):286-292.
[4] Song X, Gao H, Qian Y. Th17 differentiation and their pro-inflammation function[J]. Adv Exp Med Biol, 2014,841:99-151.
[5] Jin L, Wang G. Keratin17: a critical player in the pathognesis of psoriasis[J]. Med Res Rev, 2014,34(2):438-454.
[6] Guttman-Yassky E, Nograles KE, Krueger JG. Constrasting pathogenesis of atopic dermatitis and psoriasis-Part1: Clinical and pathologic concepts[J]. J Allergy Clin Immunol, 2011,127(5):1110-1118.
[7] Ramot Y, Paus R. Hamessing neuroendocrine controls of keratin expression: a new therapeutic strategy for skin diseases[J]. Bioessays, 2014,36(7):672-686.
[8] 沈柱,王剛,劉玉峰,等.角蛋白17上HLA-DRB1*07限制性T細(xì)胞表位的確定[J].中華微生物學(xué)和免疫學(xué)雜志,2015,25(10):790-793.
[9] 胡小平,張偉,陳辦成,等.過(guò)氧化物酶增殖物激活受體β/δ在銀屑病角質(zhì)形成細(xì)胞中的表達(dá)[J].中華皮膚科雜志,2014,47(10):719-722.
[10] Wolk K, Witte E, Wallance E, et al. IL-22 regulates the expression of gene responsible for antimicrobial defense, cellular differentiation, and mobility in keratinocytes: a potential role in psoriasis[J]. Eur J Immunol, 2006,36(5):1309-1323.
[11] Zhang W, Dang E, Shi X, et al. The pro-inflammatory cytokine IL-22 up-regulates keratin 17 expression in keratinocytes via STAT3 and ERK1/2[J]. PLoS One, 2012,7(6):e40797.
[12] Fu M, Wang G. Keratin 17 as a therapeutic target for the treatment of psoriasis [J]. J Dermatol Sci, 2012,12(67):161-165.
[13] Pattarachotanant N, Rakkhitawatthana V, Tencomnao T. Effect of gloriosa superb and catharan thus roseus extractson IFN-γ-induced keratin 17 expression in HaCaT human keratinocytes[J]. Evid Based Complement Alternat Med, 2014,2014(10):249367.
[14] 王剛,劉玉峰.治療銀屑病的一個(gè)新的藥物作用靶標(biāo)——角蛋白17[J].中國(guó)皮膚性病學(xué)雜志,2005,19(12):753-754.
[15] Rathore JS, Wang Y. Protective role of Th17 cells in pulmonary infection[J]. Vaccine, 2016,34(13):1504-1514.
[16] Loschke F, Seltmann K, Bouameur JE, et al. Regulation of keratin network organization[J]. Curr Opin Cell Biol, 2015,2(32):56-64.
[17] Miyoshi K, Takaishi M, Nakajima K, et al. Stat3 as a therapeutic target for the treatment of psoriasis: a clinical feasibility study with STA-21, a Stat3 inhibitor[J]. J Invest Dermatol, 2011,131(1):108-117.
