沉默ADAM17基因對人甲狀腺乳頭狀癌細胞增殖、遷移及侵襲的影響
2018-04-11 05:02:47葛志鵬張博王曉濤黃志強張琪田煒張國志
山東醫藥 2018年10期
關鍵詞:意義
葛志鵬,張博,王曉濤,黃志強,張琪,田煒,張國志
(1華北理工大學附屬醫院,河北唐山063000;2華北理工大學醫學實驗中心)
甲狀腺癌最常見的病理類型是乳頭狀癌。目前乳頭狀癌的治療以手術切除為主,但基因靶向治療也逐漸顯露出其優勢性和必要性。解聚素-金屬蛋白酶17(ADAM17)具有促進細胞膜釋放腫瘤壞死因子α的作用[1,2]。研究發現,ADAM17高表達與胰腺癌、乳腺癌、肝癌、宮頸癌等密切相關,能夠促進癌細胞增殖、遷移、侵襲等生物學行為[3~7]。因此,ADAM17可能是甲狀腺乳頭狀癌基因靶向治療的新位點,靶向沉默ADAM17基因可能抑制腫瘤細胞的增殖、遷移及侵襲。李晨浩等[8]發現,在甲狀腺乳頭狀癌中ADAM17 mRNA及蛋白均呈高表達,但ADAM17基因與K1細胞增殖、遷移及侵襲的關系尚不明確。2016年11月~2017年9月,我們通過靶向抑制甲狀腺乳頭狀癌細胞ADAM17的表達,觀察細胞增殖、遷移和侵襲能力的改變,旨在為以ADAM17為靶點的乳頭狀癌基因治療提供實驗支持。
1 材料與方法
1.1 材料
人甲狀腺乳頭狀癌K1細胞(華北理工大學醫學中心實驗室)。攜帶紅色熒光蛋白表達框架的ADAM17-shRNA、無意義序列-shRNA慢病毒(蘇州吉瑪基因公司),RPMI-1640培養基、胎牛血清(Gibco公司),逆轉錄試劑盒、聚合酶鏈反應試劑盒、ADAM17引物(上海Invitrogen公司),兔抗人ADAM17抗體(Abcam公司),BCA蛋白濃度檢測試劑盒(Sigma-Aldrich公司),DAB顯影液(君瑞生物技術公司),MTT細胞增殖試劑盒(上海艾睿生物科技有限公司),12孔Transwell小室(美國Corning公司),Matrigel基質膠(BD公司)。倒置熒光顯微鏡(Olympus公司),Rotor-Gene 3000熒光定量PCR儀器(Gene Company Iimited公司),微孔板檢測器、濕轉系統(上海BIO-RAD公司)。
1.2 細胞培養、分組和轉染
向K1細胞中加入含10%胎牛血清、1%青鏈霉素的RPMI-1640完全培養基,置于5% CO2、37 ℃恒溫箱中培養。將生長狀態良好且融合度達60%的K1細胞重懸,接種到6孔板中,隨機分為空白對照組、無意義序列組、轉染組,各組設6個復孔。待細胞貼壁后,轉染組、無意義序列組分別用整合有ADAM17-shRNA、無意義序列-shRNA的慢病毒轉染細胞,空白對照組加入等量PBS液。培養48 h后更換培養基,72 h后在熒光顯微鏡(400倍鏡)下計算慢病毒攜帶的紅色熒光蛋白框架的表達率。當MOI值取80時,紅色熒光蛋白表達率達到80%,說明慢病毒成功侵入K1細胞,并獲得穩定表達。
1.3 細胞中ADAM17 mRNA檢測
采用RT-PCR方法。細胞轉染72 h后,采用TRIpure法提取各組細胞總RNA,測定純度及濃度后,置于-80 ℃保存。按照逆轉錄盒步驟合成cDNA,進行PCR擴增(內參為β-actin)。引物序列:ADAM17上游5′-ATCAAACCCTTTCCTGCG-3′,下游5′-CAAACCCATCCTCGTCCA-3′;β-actin上游5′-CTGGAACGGTGAAGGTGACA-3′,下游5′-AAGGGACTTCCTGTAACAATGCA-3′。反應條件:95 ℃預變性2 min;95 ℃ 8 s,58 ℃ 35 s,72 ℃ 40 s,40個循環。應用2-ΔΔCT法計算各組ADAM17 mRNA的相對表達量。實驗重復3次,取平均值。
1.4 細胞中ADAM17蛋白檢測
采用Western blotting法。細胞轉染72 h后,提取細胞總蛋白,BCA定量法檢測總蛋白濃度。加入等量2×蛋白上樣緩沖液后煮沸變性5 min,配制10%分離膠,蛋白上樣量為40 μg。隨后電泳(110 V、100 min)、轉膜(80 V、50 min),10%脫脂奶粉封閉1 h。加ADAM17、β-actin一抗(稀釋濃度均為1∶1 000),置于搖床上,4 ℃孵育過夜;洗膜后滴加二抗(1∶1 000),37 ℃孵育30 min,洗膜后用ECL法顯色。應用Image J軟件分析條帶,計算ADAM17蛋白相對表達量。實驗重復3次,取平均值。
1.5 細胞增殖能力觀察
采用MTT法。細胞轉染72 h后,用完全培養基重懸各組細胞,制成密度為30個/μL的單細胞懸液。取100 μL,將細胞接種于3個(標記為A、B、C)96孔板中,各組設8個復孔。A孔板中的細胞培養24 h后,每孔滴加10 μL MTT溶液(濃度為5 mg/mL),細胞繼續培養4 h;棄除培養基,每孔滴加100 μL二甲基亞砜,37 ℃靜置15 min后,用酶標儀(570 nm波長)測定各孔OD值。B孔板細胞培養48 h、C孔板中細胞培養72 h后重復上述步驟。重復實驗3次。
1.6 細胞周期分布觀察
采用流式細胞儀。細胞轉染72 h后,用PBS緩沖液重懸細胞,置于預冷的70%乙醇中固定24 h,用含RNA酶的碘化丙啶(PI)染色,37 ℃避光孵育30 min,上流式細胞儀測定各組細胞的周期時相百分比。
1.7 細胞遷移能力觀察
采用Transwell遷移實驗。轉染72 h后,用胰酶將細胞消化,加入無FBS的培養基重懸,制成細胞密度為3×103/μL的細胞懸液。取100 μL接種于Transwell上層小室,下層小室滴加700 μL含20% FBS的培養基,繼續于細胞恒溫箱中培養24 h。棄除小室內培養基,小心擦去聚碳酸酯膜正面的K1細胞,PBS沖洗2次。4%多聚甲醛溶液固定1 h后,進行HE染色。各組聚碳酸酯膜于倒置顯微鏡(×100)下隨機取8個視野拍照,得各組穿膜細胞的平均數。實驗重復3次,取平均值。
1.8 細胞侵襲能力觀察
采用Transwell侵襲實驗。Matrigel基質膠用4 ℃預冷的無血清培養基稀釋10倍,取10 μL滴加在Transwell上層小室,37 ℃水平靜置30 min后, Matrigel基質膠即可凝固,重復1.7步驟。
1.9 統計學方法
2 結果
2.1 三組ADAM17 mRNA和蛋白表達水平比較
轉染組ADAM17 mRNA和蛋白表達水平較空白對照組和無意義序列組降低(P均<0.01),空白對照組和無意義序列組ADAM17 mRNA和蛋白表達水平比較差異均無統計學意義(P均>0.05)。見表1。
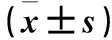
表1 三組ADAM17 mRNA及蛋白表達水平比較
注:與轉染組比較,*P<0.01。
2.2 三組細胞增殖能力比較
轉染組各時間點OD值均較空白對照組和無意義序列組降低(P均<0.05),空白對照組與無意義序列組比較差異無統計學意義(P>0.05)。見表2。
表2 三組細胞不同時間點的OD值比較
注:與轉染組比較,*P<0.01。
2.3 三組細胞周期分布比較
轉染組處于G0/G1期的細胞百分比高于空白對照組和無意義序列組(P均<0.01),出現明顯的G0/G1期阻滯;轉染組進入S期和G2/M期的細胞百分比均低于空白對照組和無意義序列組(P均<0.01);空白對照組與無意義序列組各細胞周期百分比比較無統計學差異(P均>0.05)。見表3。
2.4 三組細胞遷移能力比較
轉染72 h后,空白對照組、無意義序列組、轉染組穿膜細胞數分別為(422.8±24.1)、(432.6±25.5)、(149.2±20.7)個/HP。轉染組穿膜細胞數少于空白對照組和無意義序列組(P均<0.01),空白對照組和無意義序列組比較差異無統計學意義(P>0.05)。

表3 三組細胞周期分布百分比比較
注:與轉染組比較,*P<0.01。
2.5 三組細胞侵襲能力比較
轉染72 h后,空白對照組、無意義序列組、轉染組穿膜細胞數分別為(216.4±13.0)、(211.8±18.3)、(83.8±11.9)個/HP。轉染組穿膜細胞數少于空白對照組和無意義序列組(P均<0.01),空白對照組和無意義序列組比較差異無統計學意義(P>0.05)。
3 討論
ADAM家族自1990年發現至今已有20余個成員,它們與許多細胞的生物學功能密切相關,并具有細胞外功能區剪切酶的作用[4]。ADAM17是ADAM家族成員之一,其主要作用是促進細胞膜釋放腫瘤壞死因子α,從而參與癌癥的發生、發展。研究表明,ADAM17的高表達與甲狀腺乳頭狀癌、胰腺癌、乳腺癌、肝癌、宮頸癌等的發生及進展密切相關。
RNA干擾(RNAi)是利用小干擾RNA(siRNA)激活,具有對靶基因產生特異性抑制作用的一種實驗技術[9],siRNA以“短發夾”(shRNA)形式整合到慢病毒載體中,利用病毒的感染特性將shRNA整合到細胞染色體中,從而發揮對目的基因的沉默作用。本實驗用整合有ADAM17-shRNA的慢病毒載體轉染K1細胞,依據觀察到細胞內紅色熒光蛋白的表達率達80%,再測得轉染組ADAM17 mRNA及蛋白的相對表達量均明顯降低,證實ADAM17-shRNA成功導入K1細胞,并對ADAM17基因產生了靶向沉默作用。
無限增殖是癌細胞的一個重要生物學行為。有學者[10]認為,腫瘤是一種細胞周期性疾病,不受調控的細胞周期與無限的惡性增殖密切相關。孟祥潮等[11]發現,沉默乳腺癌MCF-7細胞中的ADAM17基因后,MCF-7細胞增殖能力顯著下降。路延芹等[12]發現,沉默乳腺癌MCF-7細胞中的ADAM17基因后,MCF-7細胞周期進展被明顯抑制。林鋒等[13]通過靶向抑制前列腺癌PC-3細胞的ADAM17基因表達,發現細胞中cyclin D1表達下降、p21表達增強,認為抑制ADAM17基因表達能夠阻滯細胞周期,降低PC-3細胞的增殖能力。本研究結果顯示,轉染組培養24、48、72 h時的OD值均較空白對照組和無意義序列組降低,細胞周期中轉染組出現了明顯的G0/G1期阻滯,處于S期、G2/M期的細胞減少。說明靶向沉默ADAM17基因能夠阻滯K1細胞進入S期和G2/M期,降低K1細胞的增殖能力,而且細胞周期阻滯現象與增殖能力的下降有關。李舜等[14]報道,沉默人膠質瘤細胞中的ADAM17基因后,細胞的增殖能力降低。本研究結果與文獻報道相符。
遷移和侵襲也是癌細胞重要的生物學行為。吳衡等[6]證實,沉默ADAM17基因能夠降低胰腺癌PANC1細胞的遷移能力。李舜等[14]通過siRNA靶向抑制ADAM17的表達及ADAM17激活劑使ADAM17過表達,雙向證明了ADAM17可通過PI3K/AKT途徑促進人膠質瘤U87MG細胞的侵襲。本研究結果顯示,轉染組Transwell遷移、侵襲實驗中的穿膜細胞數均少于空白對照組和無意義序列組,說明誘導表達ADAM17-shRNA可抑制K1細胞的遷移和侵襲能力。與楊雯靖等[15]在乳腺癌、Zheng等[16]在人膠質瘤中的研究結果相似。
綜上所述,利用整合有ADAM17-shRNA的慢病毒轉染K1細胞后,可成功沉默ADAM17基因,從而顯著抑制K1細胞的增殖、遷移、侵襲。本研究為以ADAM17為靶點的基因靶向治療提供了基礎實驗支持,進而可能為臨床中甲狀腺乳頭狀癌不能完全切除或術后復發不宜再次手術的患者,提供新的靶向治療藥物。
參考文獻:
[1] Rose-John S. ADAM17, shedding, TACE as therapeutic targets[J]. Pharmacol Res, 2013,71:19-22.
[2] Zhang P, Shen M, Fernandez-Patron C, et al. ADAMs family and relatives in cardiovascular physiology and pathology[J]. J Mol Cell Cardiol, 2016,93(4):186-199.
[3] 丁翔,楊連粵,黃耿文,等.ADAM17 mRNA在孤立性大肝癌中的表達及其意義[J].中華肝膽外科雜志,2005,11(8):46-48.
[4] 劉爽,馬榮.ADAM17在惡性腫瘤中的研究及其進展[J].現代生物醫學進展,2013,13(17):3386-3389.
[5] 韓旭,楊星飛,張雪鵬,等.ADAM17在乳腺癌組織中的表達及意義[J].現代中西醫結合雜志,2010,19(20):2486-2488.
[6] 吳衡.ADAM17在胰腺癌細胞中的表達及其對細胞增殖遷移的影響[J].江蘇大學學報(醫學版),2015,25(4):299-303.
[7] 劉爽,馬榮,賀云龍,等.宮頸鱗狀細胞癌中ADAM17的表達及其臨床意義[J].現代生物醫學進展,2016,16(15):2868-2871.
[8] 李晨浩,魏艷杰,田煒,等.ADAM17在甲狀腺乳頭狀癌中表達及意義[J].基礎醫學與臨床,2016,36(3):393-395.
[9] 孫平,趙微,趙毅玲.RNA干擾技術的原理與應用[J].醫學綜述,2011,17(2):164-167.
[10] Kwegyir-Afful AK, Murigi FN, Purushottamachar P, et al. Galeterone and its analogs inhibit Mnk-eIF4E axis, synergize with gemcitabine, impede pancreatic cancer cell migration, invasion and proliferation and inhibit tumor growth in mice[J]. Oncotarget, 2017,8(32):52381-52402.
[11] 孟祥潮,蔡準,陳國福,等.ADAM17-shRNA對人乳腺癌MCF-7細胞增殖的影響及機制[J].山東醫藥,2017,57(4):21-24.
[12] 路延芹,趙光明,張雪鵬.ADAM17-siRNA對MCF-7人乳腺癌細胞增殖的影響[J].現代中西醫結合雜志,2011,20(24):3003-3005.
[13] 林鋒,林平,劉鑫,等.靶向ADAM17基因siRNA對前列腺癌PC-3細胞增殖能力的影響[J].中華男科學雜志,2012,18(8):687-691.
[14] 李舜,唐曉平,馮凌,等.ADAM17對U87MG膠質瘤細胞增殖與侵襲的影響及作用機制[J].實用癌癥雜志,2017,32(6):883-887.
[15] 楊雯靖,張雪鵬,焦桂梅,等.ADAM17-siRNA抑制人MCF-7乳腺癌細胞體外侵襲能力的研究[J].天津醫藥,2011,39(11):1045-1047.
[16] Zheng X, Jiang F, Katakowski M, et al. ADAM17 promotes glioma cell malignant phenotype[J]. Mol Carcinog, 2012,51(2):150-164.
猜你喜歡
新少年(2022年9期)2022-09-17 07:10:54
中學生數理化·中考版(2022年12期)2022-02-16 07:36:36
小天使·一年級語數英綜合(2020年6期)2020-12-16 02:56:41
文苑(2020年12期)2020-04-13 00:54:10
中學生數理化·中考版(2019年12期)2019-09-23 06:23:28
幼兒100(2019年35期)2019-02-11 03:10:36
中學生數理化·七年級數學人教版(2017年5期)2017-11-09 03:06:19
知識經濟·中國直銷(2017年8期)2017-09-05 09:52:58
公民與法治(2016年24期)2016-05-17 04:21:50
北極光(2014年8期)2015-03-30 02:50:51
