新型植酸酶基因YkAPPAS在畢赤酵母中的表達及其酶學特性分析
2018-03-07 10:24:50朱恒文田永生李大偉于成野彭日荷姚泉洪
上海農業學報 2018年1期
朱恒文,田永生,李大偉,于成野,陳 磊,彭日荷,姚泉洪*
(1上海海洋大學食品學院,上海201306;2上海市農業科學院生物技術研究所,上海201106;3未名生物醫藥有限公司,福建361009)
磷是動物生長、繁殖及代謝所必須的礦物質元素之一,它構成機體的組成部分,還分布于皮膚、肌肉和韌帶等組織和組織液中。機體中的磷參與維持機體環境穩定、脂類化合物及脂溶性維他命的利用和轉運等很多新陳代謝及生理過程,缺乏無機磷會造成動物機體生長遲緩、食欲不振等問題。作物中的磷有60%—80%是以植酸的形式存在的[1],植酸是一種肌醇六磷酸酯,屬于維生素B族,它的水解終產物是肌醇和無機磷酸。植酸廣泛存在于農副產品中,尤其是在禾谷類和油料作物的種子中含量豐富[2],而人和單胃動物缺乏分解植酸的酶類,所以無法利用植酸中的磷。此外,植酸能與礦物質元素和蛋白質形成不溶性的穩定復合物,從而降低了礦質元素和蛋白質的利用率[3-4]。然而,研究發現將植酸酶添加到動物飼料中釋放出植酸中的磷分子,不但可以提高食物以及飼料對磷的吸收利用率,而且還能降解植酸和蛋白質的絡合物,從而降低動物排泄物中有機磷的含量,減少對自然環境的污染[5],因而將微生物植酸酶添加到動物飼料中正在被推廣和應用[6]。
植酸酶(Phytase)是催化植酸以及植酸鹽水解成肌醇和磷酸(或鹽)的一類酶的總稱[7],屬磷酸單酯水解酶。根據國際純化學與應用化學聯合會和國際生物化學聯合會定義,依據植酸酶水解植酸的位點的不同,將植酸酶分為3種類型:肌醇六磷酸3-磷酸水解酶(EC.3.1.3.8)、肌醇六磷酸6-磷酸水解酶(EC.3.1.3.26)和非特異性的正磷酸鹽單脂磷酸水解酶[8]。目前,植酸酶作為一種飼料添加劑,有著廣泛的應用前景,但植酸酶在單胃動物中的推廣和應用往往受到其酶性質的局限[9]。天然來源的植酸酶普遍存在提取困難、分泌量太低、成本高的問題,這些問題都導致其難以滿足現實生產的需要。所以構建基因工程菌作為微型生物反應器來實現異源高水平分泌表達[10],成為解決這一問題的關鍵。
本研究根據密碼子偏好性設計并合成了一個來源于Yersinia kristensenii的新型植酸酶基因YkAPPAS,并且成功地在畢赤酵母中分泌和表達。
1 材料與方法
1.1 材料
1.1.1 菌株和質粒
由本實驗室保存的大腸桿菌(Escherichia coli)DH5α菌株;巴斯德畢赤酵母GS115(His-,Mut+)購自Invitrogen公司;載體pYPX88(基因登錄號AY178045)由本實驗室構建。
1.1.2 酶和試劑
限制性內切酶、Taq DNA聚合酶、T4 DNA連接酶、蛋白marker均購自Takara公司;胃蛋白酶和胰蛋白酶以及其他試劑均購自生工生物工程(上海)股份有限公司;植酸鈉購自Sigma公司。
1.1.3 培養基
LB、YPD、BMGY和BMMY培養基。
1.1.4 主要儀器
多功能酶標儀購自TECAN Infinite M200瑞士帝肯公司;電擊儀購自Bio-Rad公司。
1.2 方法
1.2.1 表達載體構建
在不改變植酸酶氨基酸序列的前提下,根據巴斯德畢赤酵母密碼子的偏愛性對植酸酶氨基酸序列進行優化改造。采用PTDS(PCR-based two-step DNA synthesis)方法[11]將來源于Yersinia kristensenii的新型植酸酶基因進行化學合成,并且在其3’末端加上6個組氨酸標簽(便于后續表達的重組蛋白純化)。將優化后的基因連接到克隆載體pUC18上,轉化大腸桿菌DH5α進行序列測定。測序正確后,將酶切得到的基因連接到畢赤酵母表達載體pYPX88上,獲得重組質粒pYPX88-YkAPPAS。
1.2.2 表達載體的電擊轉化
將畢赤酵母GS115于50 mL YPD液體培養基中30℃恒溫過夜培養,取1 mL菌液接種于100 mL新鮮的YPD液體培養基(酵母提取物10 g/L,胰化蛋白胨20 g/L,pH 5.5,用前加入2%的葡萄糖),30℃培養至OD600=1.6—1.8,離心收集菌體,用預冷的無菌水洗菌體,4℃下8 000 r/min離心除去上清液,用20 mL預冷的山梨醇懸浮菌體,再離心收集菌體并用0.5 mL預冷的山梨醇懸浮,根據Invitrogen手冊的方法制備畢赤酵母感受態細胞。取80μL菌液,加入2μL重組質粒pYPX88-YkAPPAS,冰浴5 min后電擊,結束后立即加入800μL預冷的0.6 mol/L山梨醇溶液,混合后取0.2 mL菌液涂布于SD-山梨醇平板上,30℃培養72 h。
1.2.3 重組酵母的誘導表達
挑取陽性菌落接種在100 mL BMGY(含1%甘油)培養基中,28℃、220 r/min搖床培養至OD600值約5.0,離心收集菌體,加入適量無菌水清洗沉淀,清除殘留的甘油。將菌體加入20 mL BMMY誘導培養基中,28℃繼續誘導培養,每隔24 h添加250μL 100%甲醇至終濃度為1%,每隔6 h進行酶活測定,直到72 h。將菌液加入50 mL離心管中,于4℃下8 000 r/min離心5 min,收集上清,即為粗酶液,放置于4℃下保存。
1.2.4 重組酵母植酸酶酶蛋白的純化及分析
取誘導后的粗酶液上清1.5mL,加入已經用平衡緩沖液(50mmol/L Tris-HCl,pH 7.5,10mmol/L咪唑和300 mmol/L NaCl)平衡好的鎳柱(Ni2+-NTA agarose affinity column)中,待粗酶液完全流過鎳柱后,用洗滌液B(50mmol/L Tris-HCl,pH 7.5,20mmol/L咪唑和300mmol/L NaCl)洗滌,將雜蛋白清洗掉,最后用洗脫液 C(50 mmol/L Tris-HCl,pH 7.5,250 mmol/L咪唑和300 mmol/L NaCl)洗脫,將柱子上的重組植酸酶YkAPPAS蛋白洗脫下來。純化后的重組蛋白用SDS-PAGE進行電泳分析,并且通過考馬斯亮藍法測定蛋白含量。
1.3 重組酶酶學性質測定
植酸酶酶活單位定義為:在37℃和pH 5.5的條件下,1 min內從5.0mmol/L的植酸鈉溶液中水解釋放出1μmol無機磷所需的酶量為1個酶活單位,以U表示[12]。
1.3.1 最適pH和pH穩定性的測定
用不同pH緩沖液調節底物pH,再將不同pH的底物與酶液在37℃反應30min后,根據釩-鉬黃分光光度法測OD415計算酶的相對酶活,最適pH值就是其相對活性最高時所對應的pH。pH穩定性是在不同pH的緩沖液條件下,37℃處理24 h,然后在最適條件下(最適pH,37℃,反應30 min)測定酶活,以上每個反應設置3次重復。
本試驗所用不同 pH緩沖液為:0.2 mol/L Gly-HCl(pH 1.0—3.5),0.2 mol/L NaAc-HAc(pH 4.0—6.5),0.2 mol/L Tris-HCl(pH 7.0—8.5)和 Gly-NaOH(pH 9.0—10.0)。
1.3.2 最適溫度和溫度穩定性的測定
最適溫度的測量:先用0.2 mol/L NaAc-HAc(pH 4.0)稀釋植酸酶酶液,再把稀釋好的植酸酶酶液與底物植酸鈉在不同的溫度(10—80℃,每個溫度間隔5℃)下反應30 min后,在OD415時測定相應的酶活性,以最高相對活性所對應的溫度為最適溫度。
溫度穩定性的測定:將稀釋好的酶液分別在90℃和100℃水浴加熱5 min、10 min、15 min、20 min、25 min、30 min后立即冰置冷卻,在45℃,pH 4.5條件下反應30 min測定OD415時的相對酶活,以上每個反應設置3次重復。利用變性失活的粗酶液作為空白對照。
1.3.3 不同金屬離子及化學試劑對植酸酶的影響
試驗分兩組,以不加任何金屬離子及其他化學試劑的緩沖溶液做空白對照;試驗組將底物0.75 mmol/L植酸鈉加入NaAc-HAc(pH 4.5)緩沖溶液中,再分別加入不同的金屬離子(K+、Na+、NH4+、Ca2+、Ba2+、Li+、Ni2+、Mg2+、Pb2+、Zn2+、Fe3+、Fe2+、Cu2+、Mn2+、Al3+、Co2+和 Cr3+)、巰基化合物(DTT)、金屬螯合劑(EDTA)和變性劑(SDS),在45℃、pH 4.5反應30 min后測得重組植酸酶的剩余酶活性。
1.3.4 胃蛋白酶和胰蛋白酶對植酸酶影響的測定
試驗分兩組,以未經過消化酶液處理的初酶活作對照。用0.2 mol/L Gly-HCl(pH 2.0)和0.2 mol/L Tris-HCl(pH 7.5)緩沖液分別稀釋胃蛋白酶和胰蛋白酶,將稀釋液與植酸酶按一定重量比例(蛋白酶/植酸酶 w/w:1/500,1/250,1/100,1/50,1/1,50/1,100/1)混合,然后在 37℃下分別處理 4 h后,在最適條件下(45℃,pH 4.5)反應30 min,測定酶活,每個反應設3次重復。
2 結果與分析
2.1 YkAPPAS基因的合成
本試驗根據畢赤酵母密碼子的偏好性進行了化學合成和結構優化,并且為了便于對表達后蛋白的分離純化,在基因的3’端引入了編碼6×His的核苷酸序列。BLAST對比顯示YkAPPAS基因和野生型基因YkAPPA有69.62%的相似性(圖1)。

圖1 合成基因YkAPPAS和野生型基因YkAPPA序列比對Fig.1 Sequence alignment of YkAPPAS synthetic gene and YkAPPA w ild-type gene
2.2 SDS-PAGE分析及重組植酸酶基因在畢赤酵母中的表達
將粗酶液濃縮純化后,通過SDS-PAGE分析重組植酸酶的表達,結果如圖2所示,純化后的蛋白大小約為46 kD左右,且蛋白條帶單一,與生物信息學預測的理論分子量(46.11 kD)一致,表明重組植酸酶基因經電擊轉化后在畢赤酵母中成功表達。將表達的外源蛋白進行脫糖基化處理后,發現蛋白帶仍為46 kD,證明該外源蛋白并沒有進行糖基化修飾。
2.3 最適pH和pH穩定性
如圖3所示,重組植酸酶YkAPPAS最適pH分別為4.5和6.0,pH 6.0時酶活達到最大值,在pH 3—12時的活性都在30%以上,說明該酶的pH的適宜范圍較廣泛,耐堿性很強。
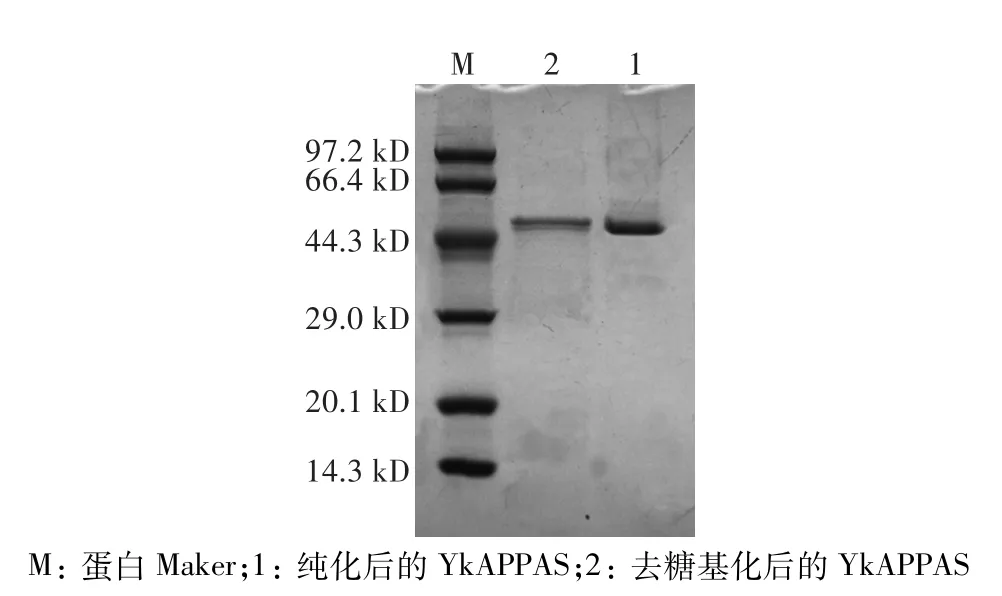
圖2 SDS-PAGE檢測YkAPPASFig.2 Detection of YkAPPAS by SDS-PAGE

圖3 植酸酶的最適pH和pH穩定性Fig.3 The optimum pH and pH stability of phytase
2.4 最適溫度和溫度穩定性
如圖4-a所示,重組植酸酶YkAPPAS最適溫度為45℃,在10—65℃有超過20%的活力。如圖4-b所示,重組植酸酶YkAPPAS在100℃處理5 min后其活性為47.55%,90℃處理5 min后其活性為56.75%,而100℃和90℃處理30 min后該酶的剩余酶活分別為4.17%和8.5%。

圖4 植酸酶的最適溫度(a)和熱穩定性(b)Fig.4 The optimum tem perature(a)and thermal stability(b)of phytase

圖5 金屬離子對植酸酶的影響Fig.5 Effects ofmetal ions on phytase activity
2.5 不同金屬離子及化學試劑對植酸酶的影響
如圖5所示,將不同的金屬離子加入含有0.75 mmol/L植酸鈉的NaAc-HAc(pH 4.5)緩沖溶液中,然后分別測定酶活性,結果表明:金屬離子Li+、Mn2+以及金屬螯合劑EDTA對重組植酸酶有一定的激活作用,其中EDTA和Li+對重組植酸酶YkAPPAS的激活作用比較明顯;K+、Na+、Fe2+、Fe3+、NH4+、Ca2+、Co2+、Ni2+、Cr3+、Al3+、Pb2+、Zn2+、Cu2+及變性劑 SDS對 YkAPPAS的酶促反應均有抑制作用,尤其是 SDS抑制力最強。Ba2+、巰基化合物DTT、Mg2+對YkAPPAS的活性無明顯影響。
2.6 蛋白酶對YkAPPAS植酸酶活性的影響
將胃蛋白酶和胰蛋白酶液與重組植酸酶按不同重量比例混合處理4 h后,植酸酶對胃蛋白酶和胰蛋白酶的耐受性如圖6所示。在胃蛋白酶濃度最低時(w/w為1/500),重組植酸酶有99%的酶活;在最強抑制作用下(w/w為100/1),重組植酸酶仍有56%的活性。隨著胰蛋白酶的比例增加,重組植酸酶的活性逐漸降低,在相對重量比為100/1時,此時抑制作用最強,酶活性最低約為60%。以上說明了重組植酸酶YkAPPAS對于胰蛋白酶和胃蛋白酶都有良好的抗水解的能力。
2.7 酶反應動力學參數
通過Lineweaver-Burk雙倒數作圖法,以反應速度的倒數(1/V)對底物質量濃度的倒數(1/[S])作圖,得到一條直線(圖7)。根據回歸方程得到該重組酶YkAPPAS的米氏常數Km=0.5686μmol/mL,最大反應速度 Vmax=1.2086μmol/(mL·min)。

圖6 胃蛋白酶和胰蛋白酶對植酸酶的影響Fig.6 Effects of pepsin and trypsin on phytase activity

圖7 雙倒數圖Fig.7 The Lineweaver-Burk plot
3 討論
本研究根據密碼子偏好性設計并合成了一個來源于Yersinia kristensenii的新型植酸酶基因YkAPPAS,根據本研究室建立的PTDS法對YkAPPAS基因進行了化學合成,并且成功在畢赤酵母中實現了高水平的異源表達。該重組植酸酶的產量較高,達到106.73μg/mL。由此可見,將該酶應用于大規模的工業生產將會降低生產成本。
與動物、真菌、細菌等表達載體相比,本研究采用畢赤酵母作為表達載體具有相對優勢。首先酵母的遺傳背景較清楚,便于遺傳操作,使用較安全;其次來自細菌和霉菌的異源植酸酶基因已經在酵母表達系統中表達;第三,現今只有少數的酵母可以表達,而主要的表達載體就是畢赤酵母[13];最后,過度的糖基化會阻礙外源蛋白的分泌運輸,而本試驗利用的巴斯德畢赤酵母GS115分泌表達的外源蛋白就沒有被糖基化修飾。
植酸酶分為PhyA和PhyB兩種類型,PhyA具有兩個最適pH,一般在pH 2.5—7.5,而PhyB只有一個最適pH[14]。有研究報道,黑曲霉植酸酶由于具有PhyA基因,其分泌出的植酸酶通常含有兩個pH最適值,例如A.niger NRRL3135和A.niger SK-57均有兩個最適pH,且這兩種黑曲霉植酸酶的最適pH均為pH 2.5和pH 5.5。除此之外,A.niger 113也有兩個最適pH,分別為pH 2.0與pH 5.0[15]。由于本試驗得到的重組植酸酶YkAPPAS也具有兩個最適pH,所以可推測出重組植酸酶屬于PhyA類型的植酸酶。目前,在動物飼養中,飼料酶制劑以其高效、安全、低成本等特點,已成為飼料添加劑研究和應用的熱點[16]。因為植酸酶是一種生物催化活性蛋白,只有當其酶活特性(如最適溫度、最適pH等)與動物腸道生理特點相吻合時,才能充分發揮其催化水解活性,所以在飼料中添加植酸酶作為添加劑主要有三方面的問題制約其大規模的生產應用,即pH最適范圍、熱穩定性以及腸胃中各種消化蛋白酶的作用[17]。本研究所得到的重組植酸酶具有明顯的優勢:首先,畜禽采食飽腹后胃內pH在4.0—5.0,且隨著胃酸的不斷分泌以及食糜的不斷排入后腸道,胃內酸度逐漸增大,進而影響植酸酶的活性[18]。而該酶具有較廣泛的最適pH,pH在1.5—7.0時其相對酶活能保持在30%以上,說明該酶在動物消化道內的適應性較強。有文獻報道,基因工程酵母生產的植酸酶在pH高于6.0時活性急劇下降,且pH在1.5以下或者7.5以上時酶活性接近為零[19]。本試驗重組植酸酶YkAPPAS有兩個最適pH,分別是pH 4.5和6.0,且pH 4.5時的酶活性是pH 6.0時的79.79%,在pH 1.0和pH 7.5時重組植酸酶YkAPPAS的酶活性都在20%以上。與文獻報道相比,本試驗的重組植酸酶耐酸性相對更強,且pH范圍相對更廣,更適宜在動物腸胃道內發揮有效活性。其次,該酶具有較好的耐熱性,90℃處理20 min后其活性為16%,能夠較好地解決飼料在生產過程中因高溫制粒而造成植酸酶活性損失的問題。有文獻選取具有代表性的商品植酸酶昕大洋進行熱穩定性試驗,發現商品酶昕大洋在90℃下處理5 min后幾乎喪失了所有酶活[20]。本試驗獲得的重組植酸酶在90℃下處理5 min后其酶活性為56.75%,與商品植酸酶昕大洋相比,重組植酸酶YkAPPAS在熱穩定性方面具有較大的優越性。
植酸能與金屬離子形成螯合物[21],抑制了植酸酶對植酸的水解作用,降低了植酸酶的活性,所以植酸酶的利用率受多種金屬離子的影響。例如Zn2+、Fe3+等離子易與植酸形成絡合物,所以重組植酸酶的活性受到了抑制作用。有文獻記載DTT對多種植酸酶的活性有強烈的抑制作用,而DTT對重組植酸酶YkAPPAS的活性卻沒有明顯影響。通過重組植酸酶對抗胃蛋白酶和胰蛋白酶能力的研究發現,在胃蛋白酶/植酸酶(w/w)為100/1,作用4 h后,酶活為56%,在胰蛋白酶/植酸酶(w/w)為500/1,作用4 h后,酶活性為60%,說明該重組酶具有良好的抗胃蛋白酶和胰蛋白酶水解的能力。
我國是農業大國,通過基因工程的手段構建植酸酶高產菌株是有重要意義的。植酸酶作為飼料給動物飼喂的效果已得到廣泛確證[22],但是在飼料工業中植酸酶還沒有得到很好的推廣利用,隨著科技的進一步發展,本試驗得到的相關酶活數據將為應用基因工程和蛋白質工程的方法對植酸酶進行改造的研究提供重要的借鑒和參考價值。
[1]朱靖環,楊永紅,毛華明.植酸酶的研究與應用進展[J].微生物學雜志,2002,22(1):43-46.
[2]彭日荷,郭美錦.應用畢氏酵母高效表達耐高溫植酸酶[J].生物化學與生物物理學報(英文版),2002,34(6):725-730.
[3]鄒立扣,王紅寧.植酸酶及其植物基因工程[J].微生物學通報,2005,32(6):128-132.
[4]HOLM PB,KRISTIANSEN K N,PEDERSEN H B.Transgenic approaches in commonly consumed cereals to improve iron and zinc contentand bioavailability[J].Journal of Nutrition,2002,132(3):514S-516S.
[5]汪世華,呂茂洲,孫長春,等.植酸酶的現狀及其研究進展[J].廣州食品工業科技,2002,18(1):54-57.
[6]CROMWELL G L.ASAS centennial paper:Landmark discoveries in swine nutrition in the past century[J].Journal of Animal Science,2009,87(2):778-792.
[7]馬璽,單安山.植酸酶研究進展及其在飼料工業中的應用[J].糧食與飼料工業,2001(4):27-30.
[8]VATSP,BANERJEE U C.Production studies and catalytic properties of phytases(myo-inositol hexakisphosphate phosphohydrolases):an overview[J].Enzyme&Microbial Technology,2004,35(1):3-14.
[9]趙翠燕,許欽坤,柯野.植酸酶的來源及合成研究進展[J].中國飼料,2009(8):11-12.
[10]姚斌,張春義,王建華,等.產植酸酶的黑曲霉菌株篩選及其植酸酶基因克隆[J].農業生物技術學報,1998,6(1):1-6.
[11]XIONG A S,YAO Q H,PENG R H,et al.A simple,rapid,high fidelity and cost-effective PCR based two-step DNA synthesis(PTDS)method for long gene sequences[J].Nucleic Acids Res,2004,32(12):437-442.
[12]VATSP,BANERJEE U C.Biochemical characterization of extracellular phytase(myo-inositolhexakisphosphate phosphohydrolase)from a hyperproducing strain of Aspergillus niger van Teighem[J].Journal of Industrial Microbiology&Biotechnology,2005,32(4):141-147.
[13]王凱,張威,李師翁.植酸酶及其應用[J].中國生物工程雜志,2015,35(9):85-93.
[14]周小玲.粗糙脈孢菌一種新植酸酶基因的克隆、鑒定和重組酶的生化特性[D].無錫:江南大學,2004.
[15]XIONG A S,YAO Q H,PENG R H,et al.Isolation,characterization,and molecular cloning of the cDNA encoding a novel phytase from Aspergillus niger 113 and high expression in Pichia pastoris[J].Journal of Biochemistry&Molecular Biology,2004,37(3):282-291.
[16]李旋亮,吳長德,李建濤.植酸酶的研究進展與應用[J].飼料博覽,2009(8):21-23.
[17]姚斌,范云六.植酸酶的分子生物學與基因工程[J].生物工程學報,2000,16(1):1-5.
[18]李富偉,汪勇,湯海鷗.胃蛋白酶、胰酶及酸度對植酸酶穩定性的影響[J].飼料與畜牧,2008(10):18-20.
[19]慕躍林,李鳳梅,王憶平.基因工程酵母產植酸酶的應用性質研究[J].飼料研究,2000(6):11-12.
[20]WANG X Y,MENG F G,ZHOU H M.The role of disulfide bonds in the conformational stability and catalytic activity of phytase[J].Biochemistry&Cell Biology,2004,82(2):329-334.
[21]賀建華.植酸磷和植酸酶研究進展[J].動物營養學報,2005,17(1):1-6.
[22]SHARMA C B,GOEL M,IRSHAD M.Myoinositol hexaphosphate as a potential inhibitor ofα-amylases[J].Phytochemistry,1978,17(2):201-204.
