澤蘭實蠅幼蟲內生細菌的分離鑒定及除草活性研究
2018-01-18 03:04:15蘭明先李建一魯武鋒李召波李麗芳吳國星
江西農業學報 2018年1期
蘭明先,張 某,李建一,魯武鋒,李召波,夏 濤,李麗芳,吳國星,高 熹
(云南農業大學 植物保護學院,云南 昆明 650201)
紫莖澤蘭EupatoriumadenophorumSpreng屬菊科澤蘭屬,為多年生惡性雜草,具較強的入侵性。紫莖澤蘭可分泌許多克生物質抑制其它植物生長,快速侵占草地牧場、田間林地、山谷河流,給入侵地造成嚴重的經濟損失。紫莖澤蘭所含的很多植物次生物質如澤蘭素A和內酯等對很多種類的昆蟲有較強的化感作用[1-3]。因此,大多數昆蟲都拒食或避忌紫莖澤蘭,僅澤蘭實蠅ProcecidocharesutilisStone專性取食紫莖澤蘭。
澤蘭實蠅是雙翅目實蠅科昆蟲,是紫莖澤蘭的天敵昆蟲,其幼蟲專性取食紫莖澤蘭莖稈的幼嫩部分,使紫莖澤蘭莖稈膨大形成蟲癭,對其生長及繁殖均有一定的抑制作用[4-6]。現在已經有很多國家和地區利用澤蘭實蠅對紫莖澤蘭進行防治[7-8]。在20世紀80年代,澤蘭實蠅越過喜馬拉雅山脈,擴散并成功定殖在西藏聶拉木縣,由此傳入我國[9]。為了防治紫莖澤蘭,云南省昆明市從西藏的聶拉木縣引入澤蘭實蠅,后來該蟲逐步擴散到云南省的大部分地區、四川的西南部地區、貴州以及廣西等省份[10-11]。
澤蘭實蠅食物中的營養成分均來自紫莖澤蘭,在澤蘭實蠅幼蟲取食的同時也攝入了大量的有害次生物質,然而幼蟲并未中毒,由此說明澤蘭實蠅對紫莖澤蘭的毒素物質有一定的耐受或者降解作用。有研究表明,共生菌的生長可以對植物的生理進行調控,抑制植物合成一些對昆蟲有不利影響的物質,從而增強昆蟲的抗逆性[12-18];昆蟲內共生菌還可對宿主昆蟲所攝取的植物次生物質(如生物堿、氰苷等)進行解毒作用,例如蘋果繞實蠅(Rhagoletispomonella)中的聚團腸桿菌(Enterobacteragglomerans)可以將植物源有毒物質根皮苷降解脫毒,有利于蘋果繞實蠅的存活[19]。在澤蘭實蠅幼蟲體內同樣有共生菌的生長,共生菌在協助其宿主營養代謝的過程中,可能也有一定的解毒作用。但在目前澤蘭實蠅共生菌的研究中,僅對澤蘭實蠅幼蟲的腸道細菌有初步的研究,還沒有系統地對澤蘭實蠅幼蟲的內生細菌進行研究[20]。
本實驗通過傳統的分離鑒定與16S rDNA序列分析相結合的方法分離鑒定了澤蘭實蠅幼蟲內生細菌的種類,并通過圓葉片法初步研究了內生細菌的除草活性,旨在為明確澤蘭實蠅對紫莖澤蘭的解毒機制提供科學依據,也為更有效地控制紫莖澤蘭奠定基礎。
1 材料與方法
1.1 供試蟲源
澤蘭實蠅老熟幼蟲采自昆明市呈貢區及本實驗室所種植的紫莖澤蘭蟲癭內。
1.2 研究方法
1.2.1 內生細菌的分離 對采回的蟲癭進行整體消毒后在超凈工作臺內解剖出澤蘭實蠅幼蟲,將幼蟲體表滅菌后解剖,獲得體內各組織[21]。分別收集所獲得的各組織于離心管內,研磨后梯度稀釋制備菌懸液。取10-5、10-6、10-7稀釋度的菌懸液各0.1 mL,分別接種在NA培養基上,每個梯度做3個重復[22]。將接菌后的培養皿倒置,于28 ℃培養2 d[23]。待長出單菌落后,進行純化培養。純化3~4次可得純化后的菌株[24]。
1.2.2 菌落特征的觀察 挑取純化后的單菌落,在LB培養基平板上劃線接種,于28 ℃培養2 d,待長出單菌落后觀察其形態[25]。
1.2.3 菌體形態的觀察 挑取純化后的單菌落,在LB培養基平板上劃線接種,于28 ℃培養2 d,獲得單菌落。挑取單菌落涂布于載玻片上,做革蘭氏染色[26]。將做好的玻片放在油鏡下觀察菌體的形態,測定菌體的大小,并記錄革蘭氏染色結果。
1.2.4 生理生化測定 將分離純化出來的菌株做細菌的生理生化實驗[27]。
1.2.5 16S rDNA的擴增和測序 將所純化分離的細菌做16S rDNA基因序列鑒定。對根據形態特征歸類的微生物類群,用試劑盒提取代表性單菌落的DNA,具體方法根據試劑盒說明書。將提取的DNA用于細菌高變動區段V4區的體外擴增,引物為16S V4區通用引物[28]。回收純化PCR產物,與pJET載體連接,進行BJIⅡ酶切鑒定。用基因轉錄間隔區引物對(515F和608R)擴增到400 bp左右的基因轉錄間隔區及核糖體DNA基因片段。由昆明擎科生物公司對16S rDNA擴增產物進行雙向測序,對所得序列在NCBI上進行BLAST比對分析。
1.2.6 共生菌對紫莖澤蘭葉片的影響 共生菌對紫莖澤蘭葉片影響的測定采用葉圓片法。選取紫莖澤蘭植株固定部位大小一致的葉片,在清水里沖洗一段時間;取出葉片晾干,用30%過氧化氫對葉表面滅菌。將滅菌后的濾紙平鋪于無菌培養皿內,實驗組加入5 mL于搖床培養24 h的菌懸液,對照組加入5 mL的無菌水。用直徑為6 mm的打孔器打取紫莖澤蘭葉圓片,將葉背面向上平放于鋪有濾紙、裝有5 mL菌懸液或無菌水的培養皿內,用保鮮膜封口后置于(25±2)℃、L∶D=12 h∶12 h的培養箱內培養,每隔24 h統計一次結果。采用分級統計方法記錄紫莖澤蘭葉圓片的受害情況。評價菌懸液對葉圓片侵害程度的標準如表1所示[29]。侵害指數的計算公式如下:


表1 評價菌懸液對葉圓片侵害程度的標準
2 結果與分析
2.1 菌落特征
通過分離純化澤蘭實蠅幼蟲的內生細菌,共獲得22株細菌,將其標號為ZLSY1~ZLSY22,并進行菌落特征觀察記錄,將結果列于表2。
2.2 菌體的形態特征
對所分離的22株菌在10×100倍油鏡下進行菌體特征及革蘭氏染色結果的觀察,結果如表3。由表3可知:有9株菌產芽孢;僅有4株菌為圓球形或者橢球形,其余18株菌均為桿菌;有13株菌為革蘭氏陽性菌,有9株菌為革蘭氏陰性菌。
2.3 生理生化特性
由表4可知:在22株菌中,有7株菌能分泌淀粉水解酶水解淀粉;有2株菌能水解油脂;有19株菌在石蕊牛奶實驗中產生反應;在明膠液化實驗中產生反應的有16株菌;有8株菌不水解硫化氫,反應結果為陰性。在葡萄糖發酵實驗中,既產酸又產氣的有3株菌,只產酸不產氣的有3株,其余均既不產酸又不產氣;在乳糖發酵實驗中,僅有1株菌既產酸又產氣,有2株既不產酸又不產氣,剩余19株菌只產酸不產氣;在吲哚實驗中,有10株菌呈陽性反應;在甲基紅實驗中有7株菌呈陽性反應;在伏-普實驗中有9株菌呈陽性反應;在檸檬酸鹽實驗中有8株菌可以使其變色而呈陽性反應。

表2 澤蘭實蠅幼蟲內生細菌的菌落特征

表3 澤蘭實蠅幼蟲內生細菌的菌體特征及革蘭氏染色結果
2.4 16S rDNA序列分析
對分離純化后的22株菌進行16S rDNA序列分析,結果如表5。表5所列是所擴增的片段與GenBank中已發表的細菌基因序列的同源性均在98%以上的基因序列。從分子水平鑒定了所分離的細菌種類,在所分離的22株澤蘭實蠅內生細菌中,有1株菌未被鑒定出來,其余21株菌隸屬10個屬,分別為迪茨菌屬(Dietzia)、芽孢桿菌屬(Bacillus)、泛菌屬(Pantoea)、類芽孢桿菌屬(Paenibacillus)、微桿菌屬(Microbacterium)、腸桿菌屬(Enterobacter)、莫拉菌屬(Moraxella)、紅球菌屬(Rhodococcus)、不動桿菌屬(Acinetobacter)、克呂沃爾菌屬(Kluyvera)。其中6株菌屬于泛菌屬,5株菌屬于芽孢桿菌屬,3株菌屬于類芽孢桿菌屬,其余屬均只有1株菌。
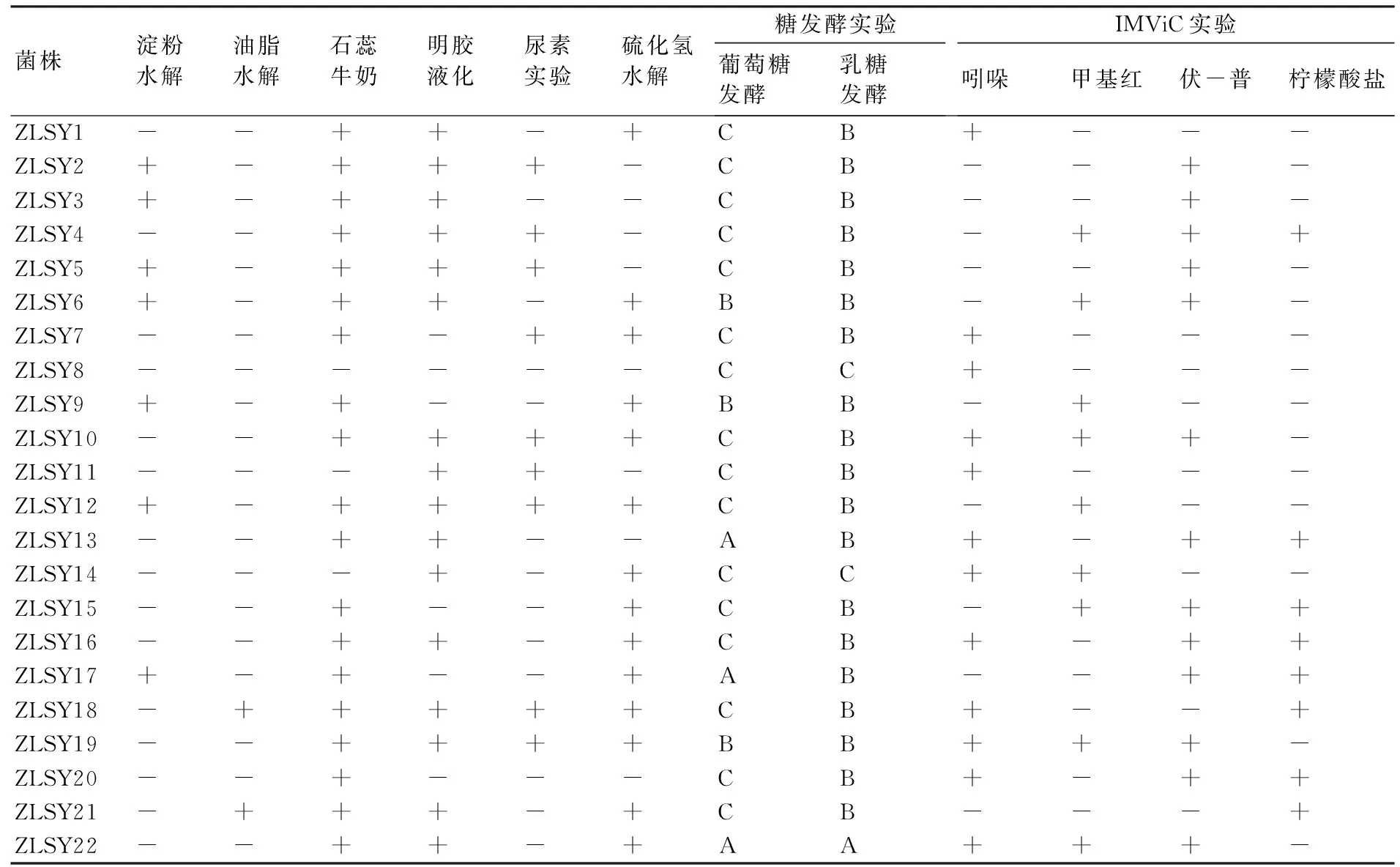
表4 澤蘭實蠅幼蟲內生細菌主要生理生化特性的實驗結果
注:“A”表示既產酸又產氣;“B”表示只產酸不產氣;“C”表示既不產酸也不產氣;“+”表示陽性反應;“-”表示陰性反應。
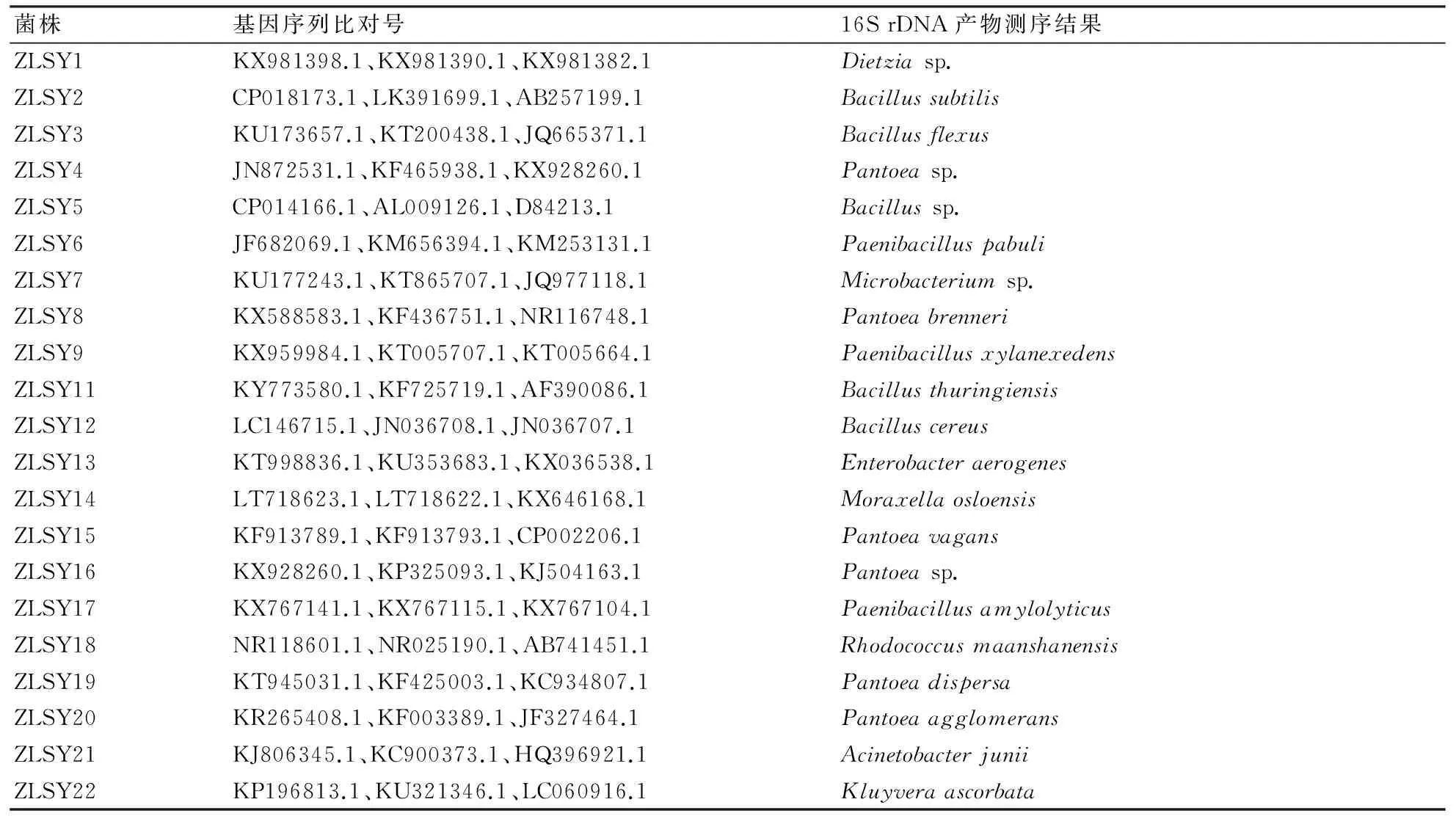
表5 澤蘭實蠅內生細菌16S rDNA序列分析結果
2.5 共生菌對紫莖澤蘭葉片的作用
所分離純化的22株菌經初步篩選有6株菌在實驗中效果很明顯,其余菌株無作用。這6株菌分別為ZLSY2、ZLSY5、ZLSY15、ZLSY16、ZLSY20、ZLSY22。由圖1可以看出,這6株菌與對照組之間的侵害效果均有顯著差異,但各株菌之間差異不顯著。隨著培養時間的增加,6株菌對紫莖澤蘭葉片的侵害程度均呈上升趨勢,在72 h時ZLSY5、ZLSY16、ZLSY22這3株菌對紫莖澤蘭葉片的傷害率達到97%以上。
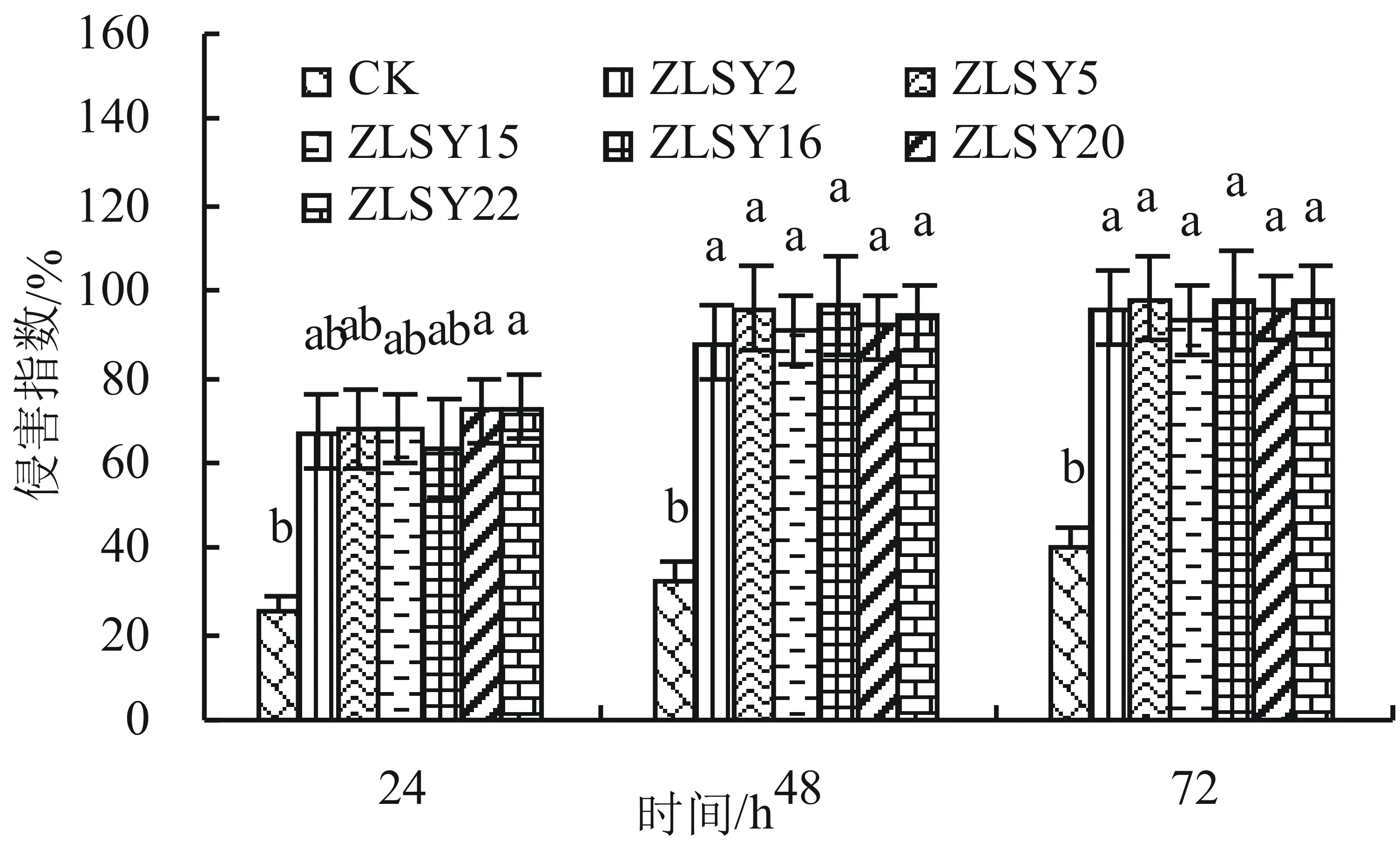
圖1 菌懸液對紫莖澤蘭葉片的侵害程度
3 討論與結論
昆蟲與其共生菌之間有著十分密切的共生關系,共生菌生長于昆蟲體內,昆蟲為共生菌創造了穩定的生活環境并提供必要的營養物質[30-31]。本實驗用形態鑒定和16S rDNA技術相結合的方法第一次分離鑒定了澤蘭實蠅幼蟲內可培養的內生細菌種類,一共分離得到22株菌,編號為ZLSY10的菌株未被鑒定出來,其余21株菌分屬于3個門(厚壁菌門、變形桿菌門、放線菌門)、3個綱、10個屬。其中變形菌門有10株菌,屬于優勢菌;厚壁菌門有8株菌,為次優勢菌。這與張某等的研究結果有所不同,他們利用Illumina HiSeq技術對澤蘭實蠅幼蟲腸道細菌進行多樣性分析時,共注釋了13個門,其中變形菌門占99%,為優勢菌[20]。出現這種差異的原因可能是所采用的方法不同,應用Illumina HiSeq技術所獲得的是腸道內整體的細菌種數,包括可培養菌和不可培養菌,因此所得的物種數量較多。在昆蟲體內生長的共生菌,其特殊的生長環境與昆蟲的內環境息息相關,大多數菌的生長環境是體外培養不能提供的;其特殊的營養物質也受昆蟲的食性以及食物來源的影響,因此可培養菌在數量上就顯得稀少。然而,可培養菌株的獲得在共生菌的研究中是非常有價值的,通過形態或分子鑒定明確其分類地位后,再進行生物學實驗,可以明確這些菌的功能[32],并可有望從中尋找某些有益細菌進行開發或利用,如找尋可降解或耐受紫莖澤蘭有毒物質的菌株。
共生菌在昆蟲體內可為昆蟲生長提供某些食物中缺乏的重要營養成分,補充其食物中的不足,起到調節營養平衡、調節代謝以及解毒作用[33]。在紫莖澤蘭植株中,若沒有某些物質的作用則植株應該均勻生長,不會出現某部分組織膨大的現象,但在有澤蘭實蠅取食的部位卻能形成蟲癭。蟲癭的形成可能是因為澤蘭實蠅幼蟲在取食過程中產生的物質改變了植株的營養代謝途徑[34];據報道,在澤蘭實蠅幼蟲體內分離到的菌株如不動桿菌屬(Acinetobacter)、紅球菌屬(Rhodococcus)的細菌都具有氮素轉化的功能,可以將體內固定的氮源轉移到體外,供寄主植物生長利用[35-36]。因此,我們猜測澤蘭實蠅體內的這兩個屬的細菌在蟲癭的生長上也起了主要作用,通過將氮源物質由體內轉移出,累積于其生活的空間中,從而產生蟲癭并快速膨大,這樣蟲癭可以增加幼蟲的取食面積,并為其提供充足的食物來源[34]。
在本實驗中,初步發現有6株菌可以使紫莖澤蘭的葉片失綠甚至腐爛,對紫莖澤蘭葉片的侵害程度較重。這6株菌分屬于芽孢桿菌屬(Bacillus)、泛菌屬(Pantoea)、克呂沃爾菌屬(Kluyvera)。芽孢桿菌屬(Bacillus)在防治植物病原菌方面是被廣泛應用的一類生防細菌,對多種植物病原真菌有防治作用[37];泛菌屬菌株Pantoeasp. LD1有降解木質素以及水稻秸稈的作用[38],在這6株菌中有3株菌是泛菌屬的;抗壞血酸克呂沃爾菌(Kluyveraascorbata)具有硝酸鹽還原能力,能將硝酸鹽還原為亞硝酸鹽,植物體能吸收低濃度的亞硝酸鹽,但高濃度的亞硝酸鹽對植物體有害[39]。在微生物對紫莖澤蘭葉片致病性的研究中,前人僅研究了鏈格孢菌的作用[29],還沒有細菌致病方面的報道。鑒于以上6株細菌對紫莖澤蘭的降解能力,在后續實驗中,我們將從菌株的安全性入手,篩選出安全的菌株進行后續的實驗。剩余的14株菌雖然對紫莖澤蘭的葉片沒有致病作用,但在抑菌活性、促進澤蘭實蠅對紫莖澤蘭的適合度方面是否存在有益作用,還有待于進一步的研究。
[1] 閆乾勝,楊婕,李華民,等.入侵物種紫莖澤蘭化學成分及生物活性研究進展[J].北京師范大學學報:自然科學版,2006,42(1):70-73.
[2] 姬艷燕,李正躍,肖春,等.紫莖澤蘭對昆蟲的化感作用[J].云南大學學報:自然科學版,2008,30(S1):143-147.
[3] 牛燕芬,莫麗玲,張永福,等.云南本地昆蟲取食紫莖澤蘭的原因研究[J].云南農業大學學報,2015,30(4):489-493.
[4] Bennett P H, Van Staden J. Gall formation in crofton weed,EupatoriumadenophorumSpreng (syn.Ageratinaadenophora), by theEupatoriumgall flyProcecidocharesutilisStone (Diptera: Trypetidae) [J]. Australia Journal of Botany, 1986, 34(4): 473-480.
[5] 馬沙,吳國星,朱家穎,等.補充蜂蜜水及交配行為對澤蘭實蠅成蟲壽命的影響[J].江西農業學報,2012,24(2):83-85.
[6] 高熹,馬沙,汪金蓉,等.澤蘭實蠅內生殖器官的結構及其發育狀況[J].生物安全學報,2013,22(1):61-65.
[7] 李愛芳,高賢明,黨偉光,等.澤蘭實蠅寄生狀況及其對紫莖澤蘭生長與生殖的影響[J].植物生態學報,2006,30(3):496-503.
[8] 馬沙,高熹,朱家穎,等.溫度及補充營養物對澤蘭實蠅壽命的影響[J].生物安全學報,2012,21(3):236-239.
[9] 蘭明先,馬沙,張某,等.澤蘭實蠅研究進展[J].南方農業學報,2017,48(3):459-464.
[10] 王吉秀,高熹,馬沙,等.Cd、Pb和Zn在土壤-紫莖澤蘭-澤蘭實蠅系統中的生物富集效應研究[J].中國生態農業學報,2013,21(7):877-882.
[11] 馬沙,吳國星,譚清,等.澤蘭實蠅雌成蟲的寄主選擇行為[J].生物安全學報,2015,24(1):26-31.
[12] Behar A, Yuval B, Jurkevitch E. Gut bacterial communities in the Mediterranean fruit fly (Ceratitiscapitata) and their impact on host longevity [J]. Journal of Insect Physiology, 2008, 54(9): 1377-1383.
[13] 徐紅星,鄭許松,劉淑平,等.昆蟲內共生菌在昆蟲防御中的作用[J].昆蟲知識,2009,46(3):350-354.
[14] Adams A S, Currie C R, Cardoza Y, et al. Effects of symbiotic bacteria and tree chemistry on the growth and reproduction of bark beetle fungal symbionts [J]. Canadian Journal of Forest Research, 2009, 39(6): 1133-1147.
[15] Tsuchida T, Koga R, Matsumoto S, et al. Interspecific symbiont transfection confers a novel ecological trait to the recipient insect [J]. Biology Letters, 2011, 7(2): 245-248.
[16] Zhang Y, Kong L, Jiang D, et al. Phytotoxic and antifungal metabolites fromCurvulariasp. FH01 isolated from the gut ofAtractomorphasinensis[J]. Bioresource Technology, 2011, 102(3): 3575-3577.
[17] Frago E, Dicke M, Godfray H C J. Insect symbionts as hidden players in insect-plant interactions [J]. Trends in Ecology and Evolution, 2012, 27(12): 705-711.
[18] 楊義婷,郭建洋,龍楚云,等.昆蟲內共生菌及其功能研究進展[J].昆蟲學報,2014,57(1):111-122.
[19] Robacker D C, Lauzon C R. Purine metabolizing capability ofEnterobacteragglomeransaffects volatiles production and attractiveness to Mexican fruit fly [J]. Journal of Chemical Ecology, 2002, 28(8): 1549-1564.
[20] 張某,楊璞,朱家穎,等.基于16S rDNA基因序列的澤蘭實蠅幼蟲腸道細菌多樣性分析[J].昆蟲學報,2016,59(2):200-208.
[21] 邵明偉,靳麗萍,張應烙.黃蜻幼蟲腸道菌QTYCOI除草活性代謝產物的分離和鑒定[J].微生物學報,2016,56(9):1513-1520.
[22] 張浩,薛妍,侯艷飛,等.腸道菌對蘇云金芽胞桿菌殺蟲活性的研究[J].生物技術通報,2012(7):176-180.
[23] 駱米娟,張賀賀,陳家鏵,等.南亞實蠅成蟲腸道細菌的分離與鑒定[J].福建農林大學學報:自然科學版,2016,45(1):8-13.
[24] 楊秀芬,劉崢,張冉,等.七種我國昆蟲病原線蟲共生菌的分離與鑒定[J].微生物學通報,2008,35(2):225-229.
[25] 沈萍,陳向東.微生物學(第8版)[M].北京:高等教育出版社,2016:15-17.
[26] 楊秀芬,劉崢,楊懷文,等.擬雙角斯氏線蟲共生細菌的分離與鑒定[J].微生物學雜志,2007,27(5):14-18.
[27] 沈萍,陳向東.微生物學實驗(第4版)[M].北京:高等教育出版社,2007:207-244.
[28] 朱詩應,戚中田.16S rDNA擴增及測序在細菌鑒定與分類中的應用[J].微生物與感染,2013,8(2):104-109.
[29] 萬佐璽.鏈格孢菌毒素及其對紫莖澤蘭的致病機理[D].南京:南京農業大學,2000.
[30] 譚周進,肖啟明,謝丙炎,等.昆蟲內共生菌研究概況[J].微生物學通報,2005,32(4):140-143.
[31] Schloss P D, Delalibera I, Handelsman J, et al. Bacteria associated with the guts of two wood-boring beetles:AnoplophoraglabripennisandSaperdavestita(Cerambycidae) [J]. Environmental Entomology, 2006, 35(3): 625-629.
[32] 柳麗君,李志紅,戴陽.實蠅共生菌研究進展[J].生物安全學報,2011,20(2):91-99.
[33] Kaltenpoth M, Winter S A, Kleinhammer A. Localization and transmission route ofCoriobacteriumglomerans, the endosymbiont of pyrrhocorid bugs [J]. FEMS Microbiology Ecology, 2009, 69(3): 373-383.
[34] 馬雙敏,虞泓,李晨程,等.植物蟲癭[J].昆蟲知識,2008,45(2):330-335.
[35] Ren Y X, Yang L, Liang X. The characteristics of a novel heterotrophic nitrifying and aerobic denitrifying bacterium,AcinetobacterjuniiYB [J]. Bioresource Technology, 2014, 171: 1-9.
[36] 王澤,徐齊,袁梅,等.菠菜內生固氮菌的分離及其功能特性研究[J].中國土壤與肥料,2015(5):116-121.
[37] 汪靜杰,趙東洋,劉永貴,等.解淀粉芽孢桿菌SWB16菌株脂肪類代謝產物對球孢白僵菌的拮抗作用[J].微生物學報,2014,54(7):778-785.
[38] 蔡紅丹,張衡,劉權.1株能夠分解木質素的泛菌的分離與鑒定[J].安徽農業科學,2014,42(9):2559-2560,2587.
[39] 賴曉芳,蘭全學,楊俊,等.姜料中抗壞血酸克呂沃爾菌特性及其產生亞硝酸鹽能力研究[J].食品工業科技,2012,33(19):63-66,67.
