新型生態浮島對改善水質的效果
2017-11-06 02:53:39王金旺夏海濤陳秋夏
浙江農業科學 2017年10期
王金旺,楊 升,夏海濤,王 新,陳秋夏*
(浙江省農業科學院 a亞熱帶作物研究所,浙江 溫州 325005; b植物保護與微生物研究所,浙江 杭州 310021)
新型生態浮島對改善水質的效果
王金旺a,楊 升a,夏海濤a,王 新b,陳秋夏a*
(浙江省農業科學院 a亞熱帶作物研究所,浙江 溫州 325005; b植物保護與微生物研究所,浙江 杭州 310021)
設計一種微生物與水生植物耦合,同時添加生物膜載體的新型生態浮島,并探索其對水質改善的作用。結果表明,生態浮島試驗區水體氨氮、總氮和總磷去除率分別為22.7%、19.2%和18.0%,但對水體硝態氮去除效果不佳,浮島內不同植物浮島單元硝態氮濃度增幅4%~18%。不同植物對水體凈化效果不同:刺果澤瀉、紫芋、千屈菜、蘆竹、風車草、菰、菖蒲對氨氮去除率達到20%以上;刺果澤瀉、紫芋、梭魚草、風車草、千屈菜、菰、蘆竹和水蔥對總氮去除率達10%以上;刺果澤瀉、菰、水蔥、紫芋對總磷去除率達15%以上。添加微納米固定化凈水菌劑(W13)可促進水體總氮、總磷的去除,但對總磷去除率的促進效果(-0.2百分點~8.5百分點)不及總氮(4百分點~15百分點)。
生態浮島; 水生植物; 微生物制劑
近幾十年來,我國社會經濟迅猛發展,水環境問題日益突出,水污染形勢非常嚴峻。據《2015年中國環境狀況公報》,全國423條主要河流、62座重點湖泊(水庫)的967個國控地表水監測斷面IV類、V類和劣V類水質斷面分別為21.1%、5.6%和8.8%[1]。在浙江,2013年監測數據表明,221個省控斷面中Ⅴ類和劣Ⅴ類水質斷面占20.8%,水環境惡化現象十分嚴重。2013年底,浙江省提出“五水共治,治污先行”,水污染的治理成為社會改革發展和經濟產業升級的關鍵舉措。經過幾年的努力,據2016年的《浙江省環境狀況公報》,全省221個省控斷面中,V類和劣V類水質斷面占11.3%[2]。
一般地,生態功能健康的河流生態系統大多具有完整的水生植被,而生態功能退化的河流則大多缺乏水生植被。水生植物是水生態系統的初級生產者,不僅能通過自身同化減低水體中N、P濃度,促進水體營養的輸出[3],而且可以改變水體流向與流強,影響水體與底泥之間的物質交換平衡[4],同時對水體中藻類生長有明顯抑制作用,具有調控水生態系統物質循環和能量流動的作用[5];因此,恢復水生植被是控制水體富營養化的一種重要的生態方法[6]。目前,在污染嚴重的水體中常采用生態浮島的方式構建水生植被來凈化水體,如上海白蓮涇世博園區河道的生態浮島對修復水質作用明顯[7]。基于微生物的修復技術在污染水體治理中也已取得不少成功的例子,如上海上澳塘、蘇州河等[8-9]。此外,生物膜載體技術也被用于水體污染治理中,提供更多的附著面以利于微生物的生長[10]。總體來看,水環境治理技術的發展越來越傾向于復合治理。傳統的生態浮島缺乏與其他生態修復技術的有效耦合,如何將其他技術耦合于以水生植物為主體的生態治理系統是近年來研發的熱點。為此,本研究擬以污染河道水體為對象,構建微生物與水生植物耦合的生態浮島,同時添加生物膜載體,探索多功能生態浮島對水質改善的作用,旨在為污染水體的綜合治理提供有益參考。
1 材料與方法
1.1 多功能生態浮島設計
生態浮島由2部分構成:上面是植物部分,下面為彈性立體填料部分(圖1)。用于承載植物和懸掛立體填料的浮體為拼接式塑料空心板,空心板為邊長32 cm的正方形,中心為直徑16 cm的可放置種植籃的空穴。立體填料懸掛于塑料空心板的四周,每根立體填料長1 m,直徑10 cm。每個塑料空心板四角設有4個小孔,利用該小孔通過按扣式連接片可將不同塑料空心板拼接在一起。試驗中將36個塑料空心板連接在一起組成面積約4 m2的浮島單元。種植籃中放置球形立體填料和碎石用于固定植物材料。8月中旬向立體填料中添加研制的增效型微納米固定化凈水菌劑(W13)。
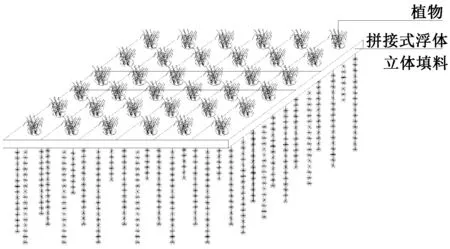
圖1 生態浮島的模型
1.2 中試試驗
試驗地位于三垟濕地半封閉河道中(27°57′20.98″N,120°41′20.14″E),半封閉水體面積450 m2。生態浮島使用的植物包括:菖蒲(Acoruscalamus)、風車草(Cyperusalternifoliusssp.flabelliformis)、菰(Zizanialatifolia)、蘆竹(Arundodonaxvar.versicolor)、千屈菜(Lythrumsalicaria)、水蔥(Scirpusvalidus)、梭魚草(Pontederiacordata)、再力花(Thaliadealbata)、刺果澤瀉(Echinodoruscordifolius)、紫芋(Colocasiatonoimo)。每個浮島單元種植1種植物,并在每個浮島單元的中心設置水樣采集標識。每種植物設置4個浮島單元,生態浮島試驗面積160 m2。生態浮島于2016年3月28日布設完畢。
1.3 水質取樣和分析
2016年5月20日至7月26日,間隔半月采集水樣1次。于2016年8月25日向生態浮島立體填料中添加增效型微納米固定化凈水菌劑(W13),因生態浮島中菰、千屈菜、菖蒲、紫芋部分植株被破壞,為使試驗數據具有有效的可比性,種植不同植物的生態浮島單元均包括36株受試植株,添加凈水菌劑的受試植物僅包括風車草、梭魚草、再力花、水蔥、蘆竹、刺果澤瀉,9月10日至11月25日間隔半月采集水樣1次。水樣采集時,用一橡膠軟管連接500 mL注射器,吸取浮島水樣采集標識處的根系周圍水樣,水樣采集深度約15 cm,每個浮島單元的水樣采集量100 mL,然后將4個種植同樣植物的浮島單元水樣混合為待測水樣,每次采樣時采集4份浮島周圍間距10 m處同深度的水樣,混合作為同期的對照水樣。樣品采集后放置于冰箱中冷藏,次日分析水質情況,測定指標包括氨氮、硝態氮、總氮、總磷,所有水質參數使用便攜式多參數分光光度計和DR2800消解器(哈希DR1900,USA)完成。
2 結果與分析
2.1 對水體氮的凈化效果
除6月3日、11月11日的監測結果外,浮島內水體氨氮監測濃度均低于浮島外(圖2)。5月20日至11月25日,浮島內所有受試植物監測位點的水體氨氮濃度均值為1.7 mg·L-1,顯著低于浮島外水體(2.2 mg·L-1),下降了22.7%。在未添加增效型微納米固定化凈水菌劑(W13)之前,浮島內所有受試植物監測位點的水體氨氮濃度均值為1.9 mg·L-1,顯著低于同期的浮島外水體氨氮濃度(2.4 mg·L-1);添加增效型微納米固定化凈水菌劑之后,浮島內所有受試植物監測位點的水體氨氮濃度(1.6 mg·L-1)雖低于同期的浮島外水體氨氮濃度(2.0 mg·L-1),但兩者差異不顯著。
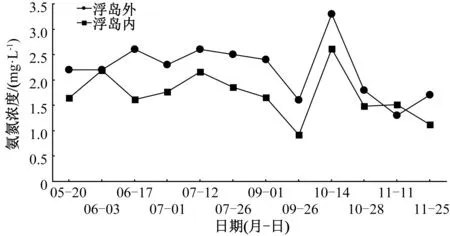
圖2 生態浮島內外氨氮濃度變化
未添加增效型微納米固定化凈水菌劑(W13)之前,除再力花之外,其余植物種植區的水體氨氮含量相比對照均有顯著降低,各種植物對氨氮的去除率分別為刺果澤瀉(38%)>紫芋(32%)>千屈菜(29%)>蘆竹、風車草(24%)>菰、菖蒲(20%)>水蔥(17%)>梭魚草(11%)>再力花(5%)(表1)。添加增效型微納米固定化凈水菌劑(W13)之后,受試植物對氨氮的去除率分別為刺果澤瀉(36%)>蘆竹(33%)>水蔥、再力花(20%)>梭魚草(17%)>風車草(13%)。蘆竹、水蔥、梭魚草、再力花相比未添加菌劑W13之前,對水體氨氮的去除率有所升高,其中再力花對氨氮的去除率增幅最大,但風車草相比未添加菌劑W13之前,對氨氮的去除率有所降低(表1)。
添加增效型微納米固定化凈水菌劑(W13)之后,即9月10日至11月25日期間,浮島內外硝態氮濃度顯著增加,浮島外硝態氮濃度相比5—7月均值增加了42.6%,浮島內硝態氮濃度增加了49.7%(圖3)。無論是否添加凈水菌劑,浮島內水體硝態氮濃度均值相比同期的浮島外水體均有所升高。未添加凈水菌劑期間,不同浮島植物單元水樣監測點硝態氮含量增加幅度為4%~18%(表2),增幅由高到低依次為水蔥、菖蒲(18%)>旱傘苗、蘆竹(16%)>千屈菜(15%)>再力花、刺果澤瀉(14%)>菰(11%)>紫芋(6%)>梭魚草(4%);添加凈水菌劑期間,不同浮島植物單元水樣監測點硝態氮含量增加幅度為11~32%(表2),增幅由高到低依次為風車草(32%)>水蔥(25%)>梭魚草(18%)
>蘆竹(16%)>再力花、刺果澤瀉(11%)。增效型微納米固定化凈水菌劑(W13)對受試植物硝態氮濃度具有增加效應的包括風車草、水蔥、梭魚草,增幅為8百分點~16百分點,具有降低效應的有蘆竹、再力花和刺果澤瀉,降幅為1百分點~3百分點。
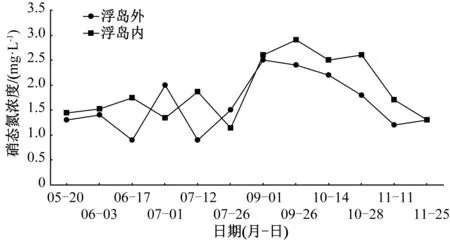
圖3 生態浮島內外硝態氮濃度變化
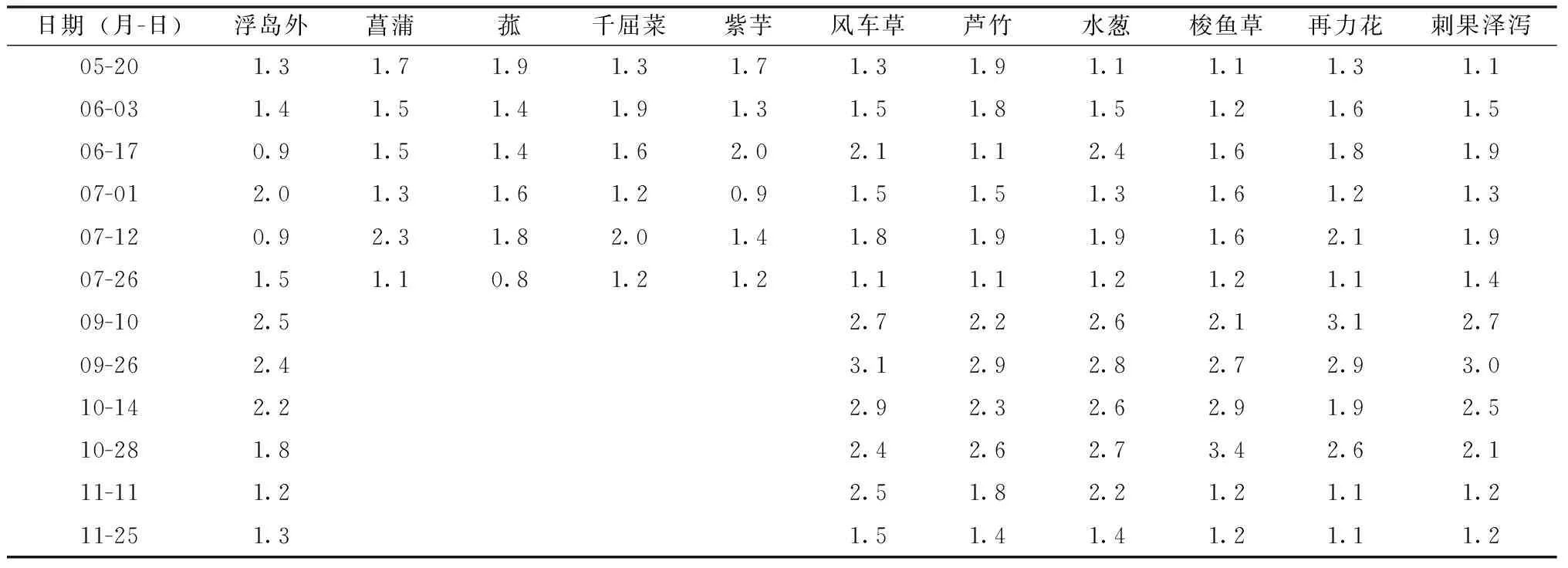
表2 生態浮島不同植物取樣點水體硝態氮含量變化 mg·L-1
試驗期間,浮島內取樣點水體平均總氮濃度(4.2 mg·L-1)顯著低于浮島外(5.2 mg·L-1),下降了19.2%(圖4)。在未添加增效型微納米固定化凈水菌劑(W13)期間,不同植物取樣點水體總氮含量相比對照,降幅由高到低依次為刺果澤瀉(27%)>紫芋(20%)>梭魚草(16%)>風車草(13%)>千屈菜(12%)>菰、蘆竹、水蔥(10%)>菖蒲(7%)>再力花(3%);添加凈水菌劑期間,不同浮島植物單元水樣監測點總氮含量降幅依次為刺果澤瀉(31%)>蘆竹(26%)>水蔥(21%)>梭魚草(19%)>風車草(18%)>再力花(15%)(表3)。增效型微納米固定化凈水菌劑(W13)對受試植物總氮去除率有促進作用,增幅為4百分點~15百分點,增幅最高的為蘆竹,最低的為梭魚草和刺果澤瀉。
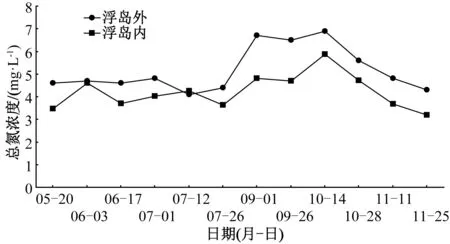
圖4 生態浮島內外總氮濃度變化
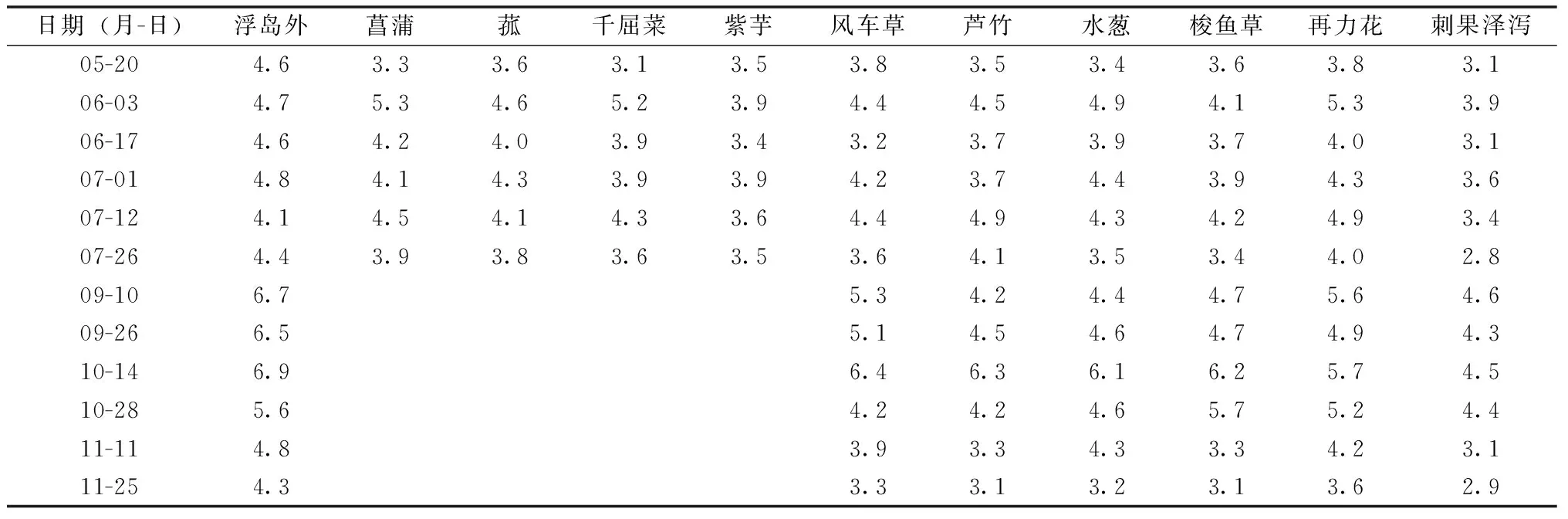
表3 生態浮島不同植物取樣點水體總氮含量變化 mg·L-1
2.2 生態浮島對水體磷的凈化效果
試驗期間,浮島內取樣點水體總磷濃度(0.73 mg·L-1)顯著低于浮島外(0.89 mg·L-1),下降了18.0%(圖5)。在未添加增效型微納米固定化凈水菌劑(W13)期間,不同植物取樣點水體總磷去除率為11%~19%(表4),總磷去除率由高到低依次為刺果澤瀉、菰(19%)>水蔥(16%)>紫芋(15%)>千屈菜(14%)>蘆竹、風車草、再力花(13%)>菖蒲(12%)>梭魚草(11%),相比同期浮島外水體,菖蒲、菰、風車草和刺果澤瀉處理下水體總磷濃度顯著(P<0.05)下降,而其余植物取樣點無顯著差異。添加增效型微納米固定化凈水菌劑(W13)之后,不同植物取樣點水體總磷去除率為13%~26%,總磷去除率由高到低依次為刺果澤瀉(26%)>水蔥(22%)>梭魚草(20%)>再力花(18%)>蘆竹(14%)>風車草(13%),相比同期浮島外水體,6種受試植物取樣點水體總磷濃度均顯著(P<0.05)下降。增效型微納米固定化凈水菌劑(W13)對增加受試植物總磷去除率的作用效果為-0.2百分點~8.5百分點,其中,效果最明顯的為梭魚草。

表4 生態浮島不同植物取樣點水體總磷含量變化 mg·L-1
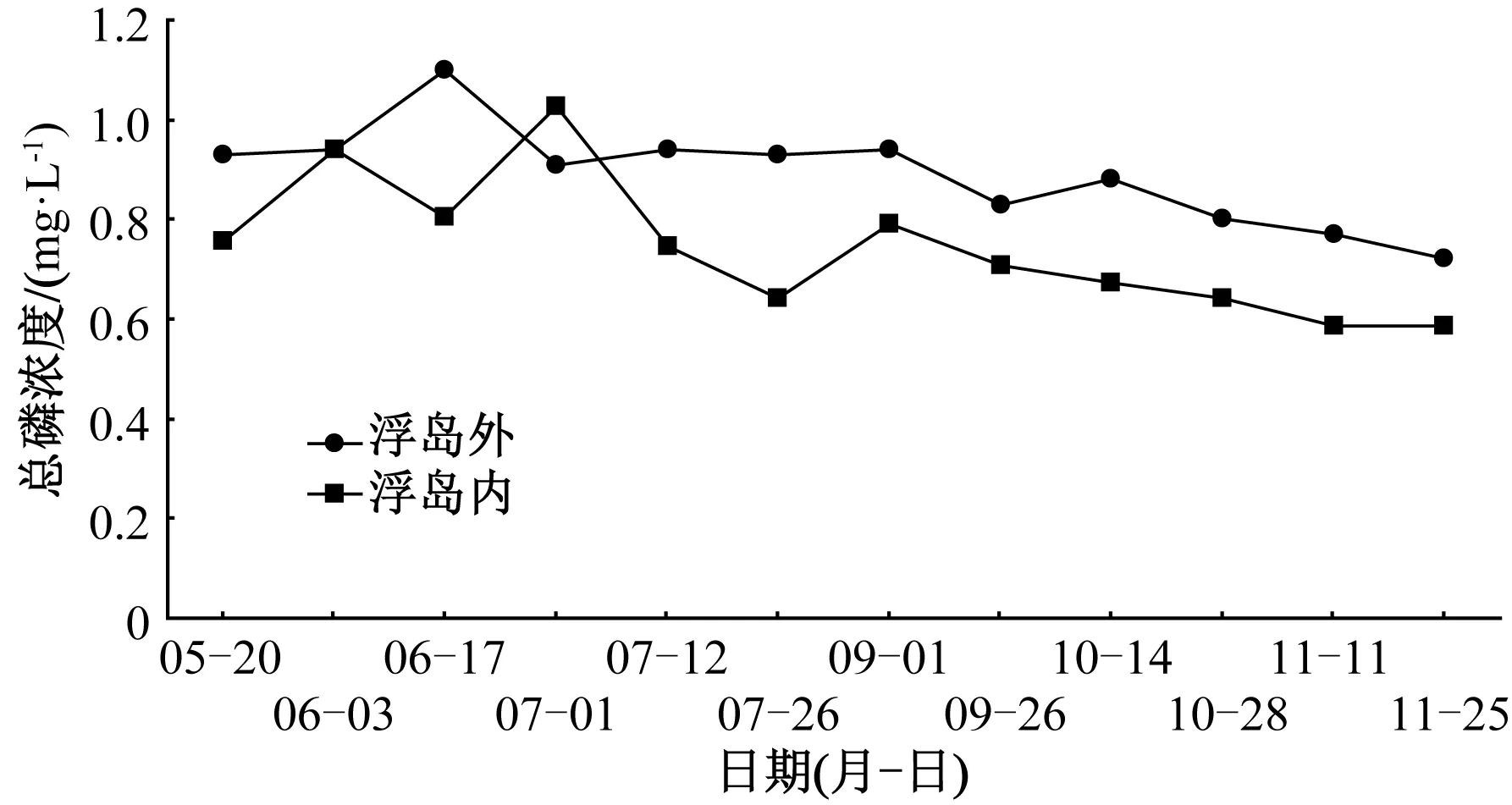
圖5 生態浮島內外總磷濃度變化
3 討論
水體富營養化是當今水環境面臨的主要問題,一般是由于水體氮磷過剩所致[3]。控制水體富營養化的方法很多,試驗中采用的新型生態浮島是一種生物-生態修復技術手段,其原理是利用植物、微生物的活動對水體中污染物進行轉移、轉化和降解[11-12],從而達到水質凈化的目的。在試驗中,未添加增效型微納米固定化凈水菌劑(W13)之前,浮島內所有受試植物監測位點的水體氨氮濃度均值均顯著低于同期的浮島外水體,與此同時,浮島內水體硝態氮濃度相比同期的浮島外水體有所升高,這可能是由于植物優先吸收氨氮和其他還原態氮,而硝態氮濃度則隨著氨氮發生硝化被轉化為硝態氮而增加[13-14]。浮島試驗區水體總氮、總磷濃度低于對照水體,是植物通過吸附沉淀、吸收代謝等途徑實現的[15]。受試植物作為水體生態系統中的初級生產者,吸收、轉化和積累了水體氮磷等營養元素,在滿足自身生長需求的同時降低了水體氮磷濃度;但不同植物對營養的需求和吸收能力不同,因而對水體氮磷的去除效果存在較大差異。陳秋夏等[16]研究了溫州三垟濕地18種水生植物生物量及對氮磷的吸收和積累能力,發現風車草、千屈菜、水芋等對氮磷去除具有較強的能力;唐藝璇[17]比較了杭州39種挺水植物的地上生物量和植物組織氮磷含量,發現澤薹草、紫芋、美人蕉、野芋等對水體氮有較強的去除能力,其組織平均氮含量超過10 000 mg·kg-1,慈姑、窄葉澤瀉、澤薹草、千屈菜、澤瀉等植物對磷的去除具有較強的能力,其組織磷含量超過3 000 mg·kg-1,可以看出,澤瀉科植物對水體氮磷的去除具有較好的效果。在本試驗中,刺果澤瀉和紫芋對總氮的去除率超過20%,刺果澤瀉、紫芋、千屈菜、蘆竹、風車草、菰和菖蒲對氨氮的去除率超過20%,刺果澤瀉、菰、水蔥、紫芋對總磷的去除率達到15%以上。刺果澤瀉為澤瀉科植物,無論是對氮還是對磷的去除,刺果澤瀉均具有較好的效果,這和上述研究結果結論相似,但各研究中不同物種對氮磷去除能力的排序可能存在一定差異,這與水體富營養化狀態及微生境條件均有一定關系[18],同時,在不同富營養化狀態水體中,植物的凈化能力亦存在較大差異[19]。
植物對水體營養物質的吸收作用相對有限,水體中污染物的降解還依賴于微生物的降解作用[20-22]。浮島植物的根系常形成一種網絡狀結構,可沉降氮磷等營養物質,同時也為微生物的附著提供了基礎,而且可改變根際周圍微生境,為微生物降解營養物質提供條件[23-24]。在試驗中,通過人工添加立體填料和增效型微納米固定化凈水菌劑(W13),氨氮總體去除率略有升高(23%>22%)。增效型微納米固定化凈水菌劑(W13)是一種蠟狀芽孢桿菌[25],兼性好氧,若W13大量增殖可代謝消耗水體溶氧使之呈現厭氧狀態,這種厭氧狀態利于反硝化作用,可提高水體硝態氮的去除。在試驗中,風車草、水蔥、梭魚草3個浮島單元,添加W13促進了硝態氮的去除;然而在蘆竹、再力花和刺果澤瀉浮島單元中則無此效果,可能是因為W13并未大量增殖,或者是添加的W13調節了水體鄉土微生物群落的結構,各種微生物菌群共同作用降解了水體營養物質,但水體的反硝化作用并不明顯,因而引起硝態氮含量的升高,其調控機制有待進一步研究。
[1] 中華人民共和國環境保護部. 圖解:2015年《中國環境狀況公報》[EB/OL]. (2016-06-03) [2017-04-11]. http://www.zhb.gov.cn/hjzl/tj/201606/t20160603_353508.shtml.
[2] 浙江省環保廳. 2015年浙江省環境狀況公報[EB/OL]. (2016-06-02) [2017-04-11]. http://www.zjepb.gov.cn/root14/xxgk/hjzl/hjzkgb/201606/t20160602_414196.html.
[3] 吳振斌,邱東茹,賀鋒,等. 水生植物對富營養水體凈化作用研究[J]. 武漢植物學研究, 2001, 19(4): 299-303.
[4] 郭輝,黃國兵. 水生植物、水流及底泥再懸浮耦合影響規律研究進展[J]. 長江科學院院報, 2013, 30(8): 108-112,116.
[5] 俞子文,孫文浩,郭克勤,等. 幾種高等水生植物的克藻效應[J]. 水生生物學報, 1992, 16(1): 1-7.
[6] 種云霄,胡洪營,錢易. 大型水生植物在水污染治理中的應用研究進展[J]. 環境污染治理技術與設備, 2003, 4(2): 36-40.
[7] 黃央央,江敏,張飲江,等. 人工浮島在上海白蓮涇河道水質治理中的作用[J]. 環境科學與技術, 2010, 33(8): 108-113.
[8] 余瑞彰,張慧,蔣俊,等. 微生物在生物柵植物根系和填料生物膜上的數量分布差異[J]. 華東師范大學學報(自然科學版), 2010(4):58-66.
[9] 徐亞同,袁磊. 上澳塘水體生物修復試驗[J]. 上海環境科學, 2000, 19(10): 480-484.
[10] 李海英,楊海華,柯凡,等. 微曝氣生態浮床的凈化效果與生物膜特性研究[J]. 中國給水排水, 2009, 25(7): 35-40.
[11] ANDERSON T A, GUTHRIE E A, WALTON B T. Bioremediation in the rhizosphere[J]. Environmental Science & Technology, 1993, 27(13):2630-2636.
[12] 張建梅. 植物修復技術在環境污染治理中的應用[J]. 環境科學與技術, 2003, 26(6): 55-57.
[13] 陳景榮,王立志. 云蒙湖前置庫淺水生態凈化區植物對水質凈化特征分析[J]. 生態科學, 2016, 35(1): 136-142.
[14] 萬曉紅,李旭東,王雨春,等. 不同水生植物對濕地無機氮去除效果的模擬[J]. 湖泊科學, 2008, 20(3): 327-333.
[15] BRIX H. Functions of macrophytes in constructed wetlands[J]. Water Science and Technology, 1994, 29(4): 71-78.
[16] 陳秋夏,鄭堅,金川,等. 水生植物對N、P的富集作用研究[J]. 江西農業大學學報, 2008, 30(3): 437-442.
[17] 唐藝璇. 杭州市富營養化河道生態浮島修復的植物選擇與示范[D]. 杭州:浙江大學,2011.
[18] 高光. 伊樂藻、輪葉黑藻凈化養魚污水效果試驗[J]. 湖泊科學, 1996, 8(2):184-188.
[19] 葛瀅,王曉月,常杰. 不同程度富營養化水中植物凈化能力比較研究[J]. 環境科學學報, 1999, 19(6): 690-692.
[20] FUHRMAN J A. Microbial community structure and its functional implications[J]. Nature, 2009, 459(7244): 193-199.
[21] JANKOWSKI K, SCHINDLER D E, HORNER-DEVINE M C. Resource availability and spatial heterogeneity control bacterial community response to nutrient enrichment in lakes[J]. Plos One, 2014, 9(1): e86991.
[22] 王新,吳逸飛,姚曉紅,等. 微生態制劑對養殖后期蝦池水質及細菌群落的影響[J]. 浙江農業學報, 2014, 26(1): 40-47.
[23] 田淑媛,王景峰,郎鐵柱,等. 水生維管束植物處理污水及其綜合利用[J]. 城市環境與城市生態, 2000(6): 54-56.
[24] 馬強,高明瑜,譚偉,等. 新型生態浮島在改善水質中的作用及生物膜載體微生物特征研究[J]. 環境科學, 2011, 32(6): 1596-1601.
[25] 王青松,柳永,王新,等. 基于質構量化分析的凈水膠囊制備及其性能研究[J]. 浙江大學學報(農業與生命科學版), 2015, 41(6): 712-722.
收入日期:2017-05-12
溫州水專項(S20140021)
王金旺(1980—),男,湖北棗陽人,副研究員,博士,從事植物學、生態學研究工作,E-mail:kingwwang@163.com。
陳秋夏(1975—),女,浙江泰順人,研究員,博士,從事林業研究工作,E-mail:yzscqx@163.com。
文獻著錄格式:王金旺,楊升,夏海濤,等. 新型生態浮島對改善水質的效果[J].浙江農業科學,2017,58(10):1837-1841,1845.
10.16178/j.issn.0528-9017.20171048
X52
A
0528-9017(2017)10-1837-05
(責任編輯高 峻)
猜你喜歡
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
