豬圓環病毒3的流行及應對措施研究
2017-08-28 19:46:01王彩霞馮春燕劉丹丹張永寧杜方原林祥梅吳紹強
中國獸醫雜志 2017年7期
關鍵詞:檢測
王彩霞 , 馮春燕 , 劉丹丹 , 張永寧 , 杜方原 , 林祥梅 , 吳紹強
(中國檢驗檢疫科學研究院動物檢疫研究所 , 北京大興100176)
豬圓環病毒3的流行及應對措施研究
王彩霞 , 馮春燕 , 劉丹丹 , 張永寧 , 杜方原 , 林祥梅 , 吳紹強
(中國檢驗檢疫科學研究院動物檢疫研究所 , 北京大興100176)
2015年6月,由于豬繁殖障礙、皮炎和腎病綜合征(PDNS)的暴發,北卡羅萊納州的一個商業豬場的母豬死亡率增加了10.2%,受孕率降低了0.6%。美國堪薩斯州立大學、愛荷華州立大學和史密斯菲爾德的研究人員對該豬場的病料組織樣品進行了檢測,排除了豬圓環病毒2型(PCV-2)、高致病性藍耳病毒(PRRSV)、甲型流感病毒(IAV)、豬細小病毒(PPV)的感染,分離到1株新病毒,并將該病毒確認為新型豬圓環病毒3型(PCV-3)[1]。Palinski等的進一步研究指出,目前PCV-3在美國已有較為廣泛的分布。隨后,我國華中農業大學在我國安徽、重慶、福建、河北、河南、湖南、江蘇、江西、遼寧、沈陽、浙江等11個省市或地區也檢測到了PCV-3[2],可見PCV-3已經開始在世界范圍內蔓延開來,逐漸引起世界各國的高度重視和密切關注,本文將對該病的發生、發展、病原學、流行情況、診斷方法及防控、應對措施等研究進行綜述。
1 發生史
豬圓環病毒是目前發現的可以自主復制的最小的哺乳動物病毒。已經確定有兩種類型的PCV,即圓環病毒1型(PCV-1)和圓環病毒2型(PCV-2)。PCV-1于1974年首次在PK細胞培養物中作為一種污染物鑒定,其對豬只沒有致病性。PCV-2于1998年首次報道,其在臨床條件下能引起豬只的豬圓環病毒相關疾病。自2010年一些研究者就報道,在豬樣品中檢測到了新的圓環病毒,直到2016年,美國的Palinski等通過宏基因組測序首次將其確認為新型豬圓環病毒3型[1]。
2 病原學
2.1 分類地位 豬圓環病毒3型屬于圓環病毒科圓環病毒屬。圓環病毒科包含3個病毒屬,分別為圓環病毒屬(Cirocovirus),環形病毒屬(Gyrovirus)和指環病毒屬(Anellovirus)。進化樹分析表明PCV-3與犬圓環病毒密切相關,然而這種關系缺乏有力的證據支持。進化樹分析還表明PCV-3和犬圓環病毒與PCV-1、PCV-2和蝙蝠圓環病毒2型的進化枝具有共同的祖先[1]。
2.2 基因組結構 豬圓環病毒(Porcinecircoviruses,PCV)是單股環狀的DNA病毒,基因組長度約為1.7 kb長,是最小的動物DNA病毒之一。首先被確診為新型圓環病毒3型的毒株名稱為2164(Genbank NO.KX458235)和29160((Gen-Bank NO.KT869077)),該病毒基因組長為2.0 kb,其基因組結構如下圖1所示,共有3個開放閱讀框(ORF),分別編碼與圓環病毒同源的REP蛋白,CAP蛋白和一個特有的ORF3蛋白, BLASTP分析顯示rep和cap在相反的方向上,且在rep基因鏈上位于rep和capORFs之間的235-nt 的5′端的基因間隔區是一個預測的帶有與PCV-1(TAGTATTAC)相同的9-nt莖-環九聚體莖環結構[3]。
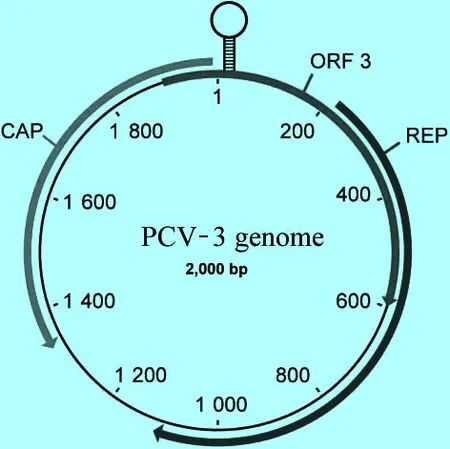
圖1 新型豬圓環病毒的基因組結構(摘自Palinski et al,2017)
最大的ORF編碼的是與圓環病毒同源的REP蛋白,含有297個氨基酸,BLASTP顯示與圓環病毒PorkNW2/USA/2009的部分復制酶蛋白的同源性為69.4%,與中國蝙蝠圓環病毒的同源性為54%。保守的圓環病毒復制酶和解旋酶區域通過BLASTP分析確定分別位于REP ORF的9~93個氨基酸和162~251個氨基酸處。進一步分析顯示,repORF蛋白序列顯示保守的滾環復制(RCR)基序和一個P-環基序,與鵝圓環病毒(GoCV)和鴿子圓環病毒(PiCV)類似[4-5]。PCV-3的3個保守的滾環復制(RCR)基序中,FTLNN基序含有單個突變,L突變為I,成為FTINN。這種突變在其他圓環病毒如GoCV中可以看到[4]。另外兩個RCR基序(HLQG和YCKK)也存在于PCV-3病毒中。而且3個基序包括WWDGY(196-200個氨基酸)、DDFYGWVP(209-2016個氨基酸)和DRYP(225-228個氨基酸)在圓環病毒復制酶蛋白中是保守的,具有未知功能。有趣的是,該ORF沒有確定的起始密碼子。在這個ORF中5′端的起始密碼子可能為GTC(編碼纈氨酸),而閱讀框中最近的ATG在下游約400 bp處。這種替代ATG起始密碼子的現象在圓環病毒PorkNW2/USA/2009中也出現過,還有在一些禽類圓環病毒,包括GoCV、PiCV和喙羽病病毒(BFDV)中也出現過[6-8]。
位于rep相反方向有一個推測的cap ORF,編碼了一個214個氨基酸的蛋白,BLASTP分析與圓環病毒PorkNW2/USA/2009的部分衣殼基因序列的同源性為87%,與PCV-2和鴨圓環病毒(DuCV)的同源性為36%~37%。同其他圓環病毒蛋白相似,N端含有許多精氨酸殘基,并且是高堿性的。保守的圓環病毒衣殼區域在26~173個氨基酸。此外,PCV-3的cap蛋白沒有預測的N-連接糖基化位點,但在第146和150個氨基酸處(分別為S和T)具有兩個預測的O-連接糖基化位點。這與PCV-2形成對比,PCV-2經試驗驗證具有兩個N-連接糖基化位點[9]。
病毒編碼的第3個ORF與預測的rep ORF在同一條鏈上,編碼一個231個氨基酸的蛋白,BLASTP分析與圓環病毒PorkNW2/USA/2009確定的ORF的同源性為94%,與鼠皰疹病毒M169,一個未知功能的蛋白的同源性為39%[10]。同rep蛋白相似,ORF3的起始密碼子不是ATG,編碼位置不清楚,5′端的密碼子是TCG(編碼絲氨酸)。也有研究推測,ORF3第55個氨基酸處的蛋氨酸可能是一個起始位點,會產生一個177個氨基酸的蛋白[1]。
對中國確診毒株的基因序列進行系統進化樹分析,結果顯示,來自中國的PCV-3分離株與美國分離株的基因組和衣殼核苷酸相似度分別為99.0%~99.1%和98.4%~98.9%。中國PCV-3分離株之間的基因組和衣殼核苷酸的相似度分別為99.0%~100%,98.4%~100%。這表明衣殼的變異程度要高于全基因組[2],中國分離株與美國分離株具有很高的同源性(見圖2)。
2.3 培養特性 PCV-3的分離培養可以采用豬睪丸細胞(ST)和豬腎細胞(PK-15)[1]進行分離。
3 流行病學
3.1 感染部位 感染了病毒的帶毒豬或病豬,死胎豬的腦、肺、淋巴結、扁桃體、精液、血液中均可檢測出PCV-3病毒[2]。
3.2 傳播途徑 死胎和精液中可檢測到PCV-3,表明PCV-3存在著垂直傳播的風險[2]。該病還可經口腔、呼吸道等途徑感染豬。
3.3 易感動物 目前的研究發現,PCV-3的易感宿主主要是豬。各種品種、年齡、不同性別的豬均可感染[11]。
3.4 發病率與死亡率
2015年6月,北卡羅萊納州的一個商業豬場與以往的數據相比,母豬死亡率增加了10.2%,受孕率降低了0.6%。每窩增加了1.19個死亡木乃伊胎,超過歷史平均流產率[1]。華中農業大學動物疾病診斷中心對收集自湖北省種豬場的14份繁殖障礙豬樣品進行了檢測,其中有12份確認為PCV-3陽性[12]。
4 臨床癥狀及病理變化
4.1 臨床癥狀 新型圓環病毒3型能引起豬的皮炎、腎病綜合征、繁殖障礙(圖3)及心臟和多系統的炎癥反應。患病母豬表現厭食,呈現多灶性丘疹、斑點和表面皮炎,豬群的死亡率增加,受孕率降低,母豬的流產率增加,生下不同胎齡的死亡木乃伊胎[1]。患病仔豬主要表現為呼吸、泌尿、腸道、淋巴、心血管、神經、繁殖系統以及皮膚的功能紊亂,對全世界的生豬養殖造成了重大的經濟損失[12]。
4.2 病理變化 組織學上,皮膚損傷的特征是急性壞死性皮炎和與淋巴漿細胞性血管套相關的表皮炎。腎臟皮質小管擴張、衰減和管狀黏膜上皮的再生,和大型集群的淋巴細胞和巨噬細胞大量地滲透到大腦皮層間質和腎小球[1]。
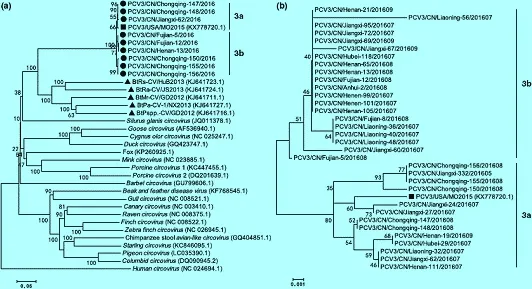
圖2 (a):不同物種圓環病毒完整基因組的系統進化樹;(b):不同的中國PCV-3分離株同美國PCV-3分離株PCV-3-USA-MO2015的部分衣殼基因的系統進化樹(摘自Ku X et al, 2017)

圖3 新型豬圓環病毒引起豬的皮膚疾病和繁殖障礙(摘自Palinski et al,2016)
5 流行情況
自2016年9月PCV-3被首次確診以來,PCV-3在美國的分布范圍已經較廣[1],在中國安徽、重慶、福建、河北、河南、湖南、江蘇、江西、遼寧、沈陽、浙江11個省市或地區也有感染報道[2]。2016年12月在中國廣東省的斷奶仔豬中也檢測到了PCV-3[13]。由于該疫病屬于新發疫病,其他國家和地區目前還未有報道。
6 診斷
6.1 鑒別診斷 豬圓環病毒相關疾病(PCVAD)臨床上表現為斷奶仔豬多系統衰竭綜合征(PMWS),豬呼吸道綜合征,繁殖障礙和豬皮炎腎病綜合征,上述癥狀并非PCV-3的特異性癥狀,因此,對患豬進行鑒別診斷時,需要首先排除PCV-2、高致病性藍耳病毒(PRRSV)、甲型流感病毒(IAV)和豬細小病毒(PPV)的感染或其他可引起上述癥狀的因素,然后通過進一步的實驗室檢驗來進行確診[1]。
6.2 病毒的分離與鑒定 病毒的分離與鑒定是確診疫病最準確的檢測方法,由于成年患病動物常缺乏臨床癥狀而導致病毒血癥期難以掌握,從而影響病毒分離。因此,通常從流產胎兒或死胎的組織樣品中分離病毒[14]。Palinski等[1]利用ST和PK-15細胞從胎兒組織勻漿中分離到了PCV-3。PCV-3可以在不同的豬組織中檢測到,例如腦、肺、淋巴結、扁桃體、精液和血清中均可檢測到[2]。
6.3 分子生物學檢測方法 目前,Palinski等[1]在實驗室研究中已經針對PCV-3建立了PCR、定量熒光PCR方法、免疫組化檢測等方法,并針對cap蛋白制備單克隆抗體建立了PCV-3的免疫熒光檢測方法。Phan等[15]利用靶向PCV-3的探針進行原位雜交調查了心臟和肺中的PCV-3,結果顯示,在呈現彌漫性肌漿/胞漿性反應的多灶性心肌細胞、發炎小動脈中膜的平滑肌細胞和心肌中的炎癥細胞(大概是巨噬細胞)中較少檢測到PCV-3 mRNA,在肺切片中未檢測到PCV-3 mRNA。中國的Ku等[2]設計PCR引物對中國11個省市或地區的35個農場的222份樣品進行了PCV-3檢測,結果發現11個省市或地區的樣品均可檢測到PCV-3。但值得一提的是,目前這些技術還未形成商品化的試劑盒,都僅停留在實驗室階段。
6.4 血清學檢測方法 Palinski等利用純化的重組衣殼蛋白包被ELISA板子,建立了PCV-3的ELISA檢測方法[1]。
7 防控
圓環病毒病是國際公認的危害養豬業的重大經濟影響性疾病,是養豬業的3大疾病(豬瘟、藍耳病、豬圓環病毒病)之一。對PCV-3的防控,首先要強化生物安全,提高相關人員的消毒意識,嚴格執行消毒規范。重視防蚊滅蠅,清潔環境,保持衛生。定期滅鼠。其次應加強飼養管理,改善豬場環境,控制營養結構,做好引種考察工作,及時隔離淘汰病豬,嚴密處理死豬及相關飼喂設施[11,16-17]。此外,PCV-2與PCV-3的衣殼蛋白之間只有30%的同源性,利用PCV-2疫苗產生交叉保護的可能性較小[1],因此,必須盡快研制優質高效的PCV-3疫苗,通過疫苗免疫來防控該病。同時還應定期對豬群進行投藥保健,增強豬群自身的抵抗力。
8 我國需采取的應對措施
8.1 加快檢測技術儲備研究 PCV-3是一種新型病毒,將可能對我國養豬業構成巨大威脅,為了及時監控疫情,防止疫情的擴散與傳播,必須加強檢測技術研究,同時進一步研究探索其生物和致病特征并監測其單一和共感染狀態,以達到精準快速的確診疫情。
8.2 做好疫病風險評估 我國的母豬及流產胎兒中已經檢測到PCV-3病原,遺傳進化樹分析,與美國PCV-3分離株的序列同源性較高。目前不但要防止外來新的疫情傳入,還要防止國內疫情的傳播,為避免疫情傳入,對進口國豬肉及其產品,以及種豬的進口,相關部門應根據該病的最新研究進展做好風險評估工作,并基于準確的風險評估結果做出最終的貿易措施。為避免疫情在國內擴散,應在國內開展廣泛的流行病學調查,及時做好疫情監控工作,做到防患于未然。
8.3 加強檢驗檢疫監管 加強對來自疫區的貨物、攜帶物、郵寄物、運輸工具的查驗和防疫消毒工作,加強出入境檢驗檢疫監管力度,嚴防不合格產品的傳入與傳出[14]。
8.4 加強疫病監測 及時跟蹤國內外疫情動態,加強對國內所有易感動物和媒介昆蟲的監測。若發現可疑病例應立即上報,并封鎖疫區,及時開展流行病學和血清學調查,并嚴格控制疫區易感動物的流動[14]。
[1] Palinski R, Pieyro P, Shang P,etal. A novel porcine circovirus distantly related to known circoviruses is associated with porcine dermatitis and nephropathy syndrome and reproductive failure[J]. J Virol. 2016,16;91(1):01 879-01 916.
[2] Ku X, Chen F, Li P,etal. Identification and genetic characterization of porcine circovirus type 3 in China[J]. Transbound Emerg Dis. 2017, 64(3):703-708.
[3] Li L, Kapoor A,Slikas B,etal. Multiple diverse circoviruses infect farm animals and are commonly found in human andchimpanzee feces [J]. J Virol, 2010, 84(4):1 674-1 682.
[4] Todd D, Weston J H,Soike D,etal. Genome sequence determinations and analyses of novel circoviruses from goose and pigeon [J].Virology. 2001, 286(2):354-362.
[5] Ilyina T V, Koonin E V. Conserved sequence motifs in the initiator proteins for rolling circle DNA replication encoded by diverse replicons from eubacteria, eucaryotes and archaebacteria[J]. Nucleic Acids Res, 1992, 20(13):3 279-3 285.
[6] Niagro F D, Forsthoefel A N, Lawther R P,etal. Beak and feather disease virus and porcine circovirus genomes: intermediates between the geminiviruses and plant circoviruses[J]. Arch Virol, 1998,143(9):1 723-1 744.
[7] Phenix K V, Weston J H, Ypelaar I,etal. Nucleotide sequence analysis of a novel circovirus of canaries and its relationship to other members of the genus circovirus of the family Circoviride[J]. J Gen Virol, 2001,82(Pt11):2 805-2 809.
[8] Bassami M R,Ypelaar I, Berryman D,etal. Genetic diversity of beak and feather disease virus detected in psittacine species in Australia[J]. Virology, 2001,279(2):392-400.
[9] Lv Q, Guo K, Zhang Y. Current understanding of genomic DNA of porcine circovirus type 2[J]. Virus Genes, 2014,49(1):1-10.
[10] Zhang W, Li L, Deng X,etal. What’s for dinner? Viral metagenomics of US store bought beef, pork, and chicken[J]. Virology, 2014, 468-470:303-310.
[11] 孫賢.豬圓環病毒2型的流行病學調查與綜合防控措施研究[D].南京:南京農業大學,2012.
[12] Fan S, Ku X, Chen F,etal.Complete Genome Sequence of a Novel Porcine Circovirus Type 3 Strain, PCV3/CN/Hubei-618/2016, Isolated from China[J]. Genome Announc,2017, 5(15): 1-2.
[13] Shen H, Liu X, Zhang P,etal.Genome characterization of a porcine circovirus type 3 in South China[J].Transbound Emerg Dis,2017,00:1-3.
[14] 張永寧,吳紹強,劉建,等.歐洲暴發施馬倫貝格病疫情及我國應采取的對策[J].中國獸醫雜志,2012, 48(4):92-95.
[15] Phan T G,Giannitti F, Rossow S,etal. Detection of a novel circovirus PCV-3 in pigs with cardiac and multi-systemic inflammation[J]. Virol J, 2016 Nov 11;13(1):184.
[16] 劉建波,張輝,劉長明.豬圓環病毒病的流行趨勢與防控對策[J].動物醫學進展,2014, 35(1): 111-115.
[17] 田曉婷,李寶玉,柳紀省.豬圓環病毒的分子生物學與檢測方法的研究現狀[J].中國農學通報,2012,28(14):66-72.
2017-05-26
十三五國家重點研發計劃課題(2016YFD0501105、2016YFD0501106)
王彩霞(1982-),女,助理研究員,碩士,從事動物檢疫工作,E-mail:friday128@sina.com
馮春燕(1982-),女,副研究員,博士,從事動物檢疫工作,E-mail:fengcy@caiq.gov.cn
林祥梅,E-mail:linxm@caiq.gov.cn;吳紹強,E-mail:sqwu@sina.com
S852.65+1
B
0529-6005(2017)07-0101-04
注:馮春燕與王彩霞對本文具有同等貢獻
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
