靶向YWHAE基因shRNA慢病毒表達質粒的構建及功能驗證
2017-08-11 13:01:48張曉艷溫春燕呂玉華佘菲菲
福建醫科大學學報 2017年3期
關鍵詞:胃癌
張曉艷, 溫春燕, 呂玉華, 陳 豪, 佘菲菲
?
靶向YWHAE基因shRNA慢病毒表達質粒的構建及功能驗證
張曉艷, 溫春燕, 呂玉華, 陳 豪, 佘菲菲
目的 構建靶向YWHAE基因的shRNA慢病毒表達質粒,并驗證其對AGS胃癌細胞增殖的影響。 方法 設計并合成5對針對人YWHAE基因的shRNA序列,分別克隆入pLentiLox3.7(pLL3.7)慢病毒表達質粒,接著將重組的慢病毒表達質粒和包裝質粒PHR、包膜質粒VSVG一起采用磷酸鈣法轉染293T細胞包裝慢病毒,收集制備的慢病毒感染AGS細胞,用抗生素Puromycine進行篩選。Western-blot檢測感染細胞YWHAE蛋白的表達。選取干擾效率最高的慢病毒表達質粒,針對性設計siRNA的點突變引物,重疊延伸PCR法擴增獲得點突變的YWHAE基因,克隆入pcDNA3.1/myc-His(-)A載體,構建YWHAE點突變的表達質粒,轉染YWHAE沉默效果最好的AGS細胞株,Western-blot檢測轉染細胞YWHAE蛋白的回復表達。采用MTS法檢測YWHAE-shRNA慢病毒對AGS細胞增殖的影響。 結果 5對針對人YWHAE基因的shRNA序列構建的慢病毒中,pLL3.7-siYWHAE-5包裝成的慢病毒抑制AGS細胞的YWHAE蛋白表達的效果最明顯,僅為對照組相對表達量的(0.269±0.083)倍;pLL3.7-siYWHAE-5包裝的慢病毒,其干擾效應可經由YWHAE點突變表達質粒回復。與對照組比較,YWHAE-shRNA組AGS細胞增殖能力明顯下降。 結論 成功構建了靶向YWHAE基因的shRNA慢病毒表達質粒,獲得YWHAE基因表達顯著下調的AGS細胞株;探明YWHAE表達的下調可有效抑制AGS細胞的增殖,提示YWHAE在胃癌中的致癌潛能。
慢病毒屬; RNA干擾; 胃腫瘤; 細胞增殖
在全球高發腫瘤中,胃癌位居第4位,死亡率居癌癥死亡率的第2位,嚴重威脅人類的健康,探討胃癌的發病機制成為當前研究的熱點[1]。酪氨酸3-單加氧酶/色氨酸5-單加氧酶激活蛋白ε肽(tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, epsilon, YWHAE)是14-3-3蛋白家族的ε亞型,是生物進化中最保守的蛋白之一[2]。在胃癌的相關研究中,YWHAE扮演的角色目前尚存在爭議。幽門螺桿菌是公認的致癌原,Nagappan等用該菌感染Gulo-/-(L-gulono-gamma-lactone oxidase基因缺失)小鼠,在胃組織中檢測到YWHAE高表達[3-4]。Gong等通過免疫組織化學染色發現,YWHAE在胃癌組織中高表達[5]。但Leal等發現,YWHAE蛋白在胃癌組織中的表達明顯低于癌旁組織[6]。在胃癌組織中,YWHAE究竟是抑癌還是促癌,單純從蛋白的表達水平上判定存在一定的局限性。從實質上探討YWHAE的功能作用以明確YWHAE與胃癌的關系,從而探明YWHAE在胃癌的發生發展中所起的作用,對胃癌的防治有重大的意義。本研究擬構建靶向YWHAE基因shRNA慢病毒表達質粒,建立YWHAE表達下調的胃癌細胞模型,并檢測其對細胞增殖的影響,為調控YWHAE在胃癌中的相關功能的深入研究奠定基礎。
1 材料和方法
1.1 材料
1.1.1 質粒、菌株和細胞株 pLentiLox3.7(pLL3.7)、PHR和VSVG(消化道惡性腫瘤教育部重點實驗室保存);pcDNA3.1/myc-His(-)A、大腸桿菌Stable3和DH5α(美國Invitrogen公司);人胃腺癌上皮細胞系AGS(中國中科院上海細胞庫);293T細胞(美國ATCC公司)。
1.1.2 試劑 胎牛血清(FBS,德國PAN公司);Ham’s-F12培養液(美國HyClone公司);DMEM高糖培養液和Puromycine(美國Gibco公司);XhoI、HpaI、EcoR I、XbaI和KpnI(美國NEB公司);高保真DNA聚合酶(美國Invitrogen公司);Western及IP細胞裂解液、BCA法定量試劑(中國Beyotime公司);anti-14-3-3ε(T-16)(美國Santa Cruz公司);Myc-Tag (9B11) Mouse mAb(美國CST公司);goat anti-mouse IgG-AP(德國Merck公司);anti-β-actin(美國Sigma公司);FuGENE 6 Transfection Reagent和Cell Titer 96 AQueous One Solution Reagent(美國Promega公司)。
1.2 方法
1.2.1 細胞培養 人胃腺癌上皮細胞系AGS培養于含10% FBS的Ham’s-F12培養液中,293T細胞培養于含10% FBS的DMEM高糖培養液中。二者均置于37 ℃、體積分數為0.05的CO2、飽和濕度的細胞培養箱中孵育培養,2~3 d換液傳代1次。
1.2.2 重組慢病毒pLL3.7-YWHAE-shRNA表達質粒的構建 根據GenBank數據庫提供的YWHAE(NM_006761.4)基因編碼序列,利用美國Invitrogen公司提供的在線RNA干擾序列設計軟件及BLOCK-iTTMInducible H1 Lentiviral RNA System手冊中提供的設計規則,按照正義鏈模板:5′-T-(GN18)-(TTCAAGAGA)-(81NC)-TTTTT
TC-3′,反義鏈與正義鏈互補,并在其5′端加入XhoI酶切位點(TCGAG),設計5對針對人YWHAE基因的shRNA序列引物,1對無義的陰性對照序列引物由上海Invitrogen生物公司合成,具體序列如下(F:正義鏈;R:反義鏈)。將合成的引物退火,形成雙鏈DNA oligo,克隆入經XhoI和HpaI雙酶切后的pLL3.7慢病毒表達質粒中,構建重組的pLL3.7-YWHAE-shRNA表達質粒和pLL3.7-NC-shRNA陰性對照質粒,轉化大腸桿菌感受態細胞Stable3,挑取重組陽性菌落抽提質粒行雙酶切及測序(福州鉑尚生物技術有限公司)鑒定。
siYWHAE-1:
F:5′-TGCTGAGCAGTTGTCCGCGTTTCAAGAGAACGCGGACAACTGCTCAGCTTTTTTC-3′
R:5′-TCGAGAAAAAAGCTGAGCAGTTGTCCGCGTTCTCTTGAAACGCGGACAACTGCTCAGCA-3′
siYWHAE-2:
F:5′-TGCAGTTGTCCGCGTGCGCATTCAAGAGATGCGCACGCGGACAACTGCTTTTTTC-3′
R:5′-TCGAGAAAAAAGCAGTTGTCCGCGTGCGCATCTCTTGAATGCGCACGCGGACAACTGCA-3′
siYWHAE-3:
F:5′-TGGAAGGAGGCTGCGGAGAATTCAAGAGATTCTCCGCAGCCTCCTTCCTTTTTTC-3′
R:5′-TCGAGAAAAAAGGAAGGAGGCTGCGGAGAATCTCTTGAATTCTCCGCAGCCTCCTTCCA-3′
siYWHAE-4:
F:5′-TGAAGAAAGCTATAAGGACTTTCAAGAGAAGTCCTTATAGCTTTCTTCTTTTTTC-3′
R:5′-TCGAGAAAAAAGAAGAAAGCTATAAGGACTTCTCTTGAAAGTCCTTATAGCTTTCTTCA-3′
siYWHAE-5:
F:5′-TGAGGAGAAGACAAGCTAAATTCAAGAGATTTAGCTTGTCTTCTCCTCTTTTTTC-3′
R:5′-TCGAGAAAAAAGAGGAGAAGACAAGCTAAATCTCTTGAATTTAGCTTGTCTTCTCCTCA-3′
siRNA-NC:
F:5′-TGTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGACTTTTTTC-3′
R:5′-TCGAGAAAAAAGTCTCCGAACGTGTCACGTTCTCTTGAAACGTGACACGTTCGGAGACA-3′
1.2.3 慢病毒的包裝 用磷酸鈣轉染法轉染293T細胞進行慢病毒包裝。具體步驟如下:轉染前1天,將293T細胞鋪于35 mm板中,當密度達到匯合度60%~70%即可進行轉染。在轉染前1 h,用新鮮無血清的DMEM培養液進行細胞換液。按下列劑量配制轉染試劑體系:質粒(pLL3.7 2 μg,PHR 1.5 μg,VSVG 1 μg),2.5 mol/L的CaCl210 μL,補充水至100 μL,混勻,再加入2×HBS 100 μL,靜置20~30 min。將混合物混勻后均勻滴到293T細胞中,輕輕搖動,置培養箱內培養8 h。更換新鮮的完全培養液,并加NEAA(100×) 20 μL,繼續培養至48 h(從轉染時開始計算時間),收集上清分裝入EP管中,6 500 r/min離心5 min,吸取上清,用0.45 μm濾膜過濾,收集濾液即病毒液。
1.2.4 慢病毒感染AGS細胞 將AGS細胞以匯合度為50%~60%均勻鋪于35 mm板,培養14~16 h。將收集到的病毒液均勻滴加至AGS細胞中,輕輕搖勻,放置培養箱培養。24 h后換新鮮完全培養液。待細胞匯合度為80%~90%時,將細胞從35 mm板消化轉移至2個6 cm板(一板用于篩選細胞株,一板用于驗證表達情況),培養液分別補足至5 mL,加抗生素Puromycine,用最低致死量0.5 μg/mL篩選。待6 cm板細胞匯合度為80%~90%,轉移至10 cm板,繼續用最低致死量的Puromycine培養1周。1周后抗生素劑量減半,再篩選1周(時間從感染開始計算),收集細胞,凍存于-80 ℃。
1.2.5 YWHAE-shRNA 慢病毒干擾細胞蛋白表達功能的鑒定 Puromycine篩選結束后,各感染組收集部分細胞,用Western及IP細胞裂解液分別提取總蛋白,BCA法定量測定蛋白濃度后,取等量蛋白進行Western-blot驗證[一抗anti-14-3-3ε(T-16),1∶200稀釋;二抗goat anti-mouse IgG-AP,1∶2 000稀釋;內參anti-β-actin,1∶2 000稀釋],觀察慢病毒介導的RNAi對AGS細胞YWHAE表達的沉默效果。用Image J 1.48軟件分析蛋白條帶的灰度,并以β-actin為內參,計算YWHAE蛋白的表達量。
AGS-siYWHAE細胞YWHAE蛋白相對表達水平=YWHAE表達量siYWHAE-5組/YWHAE表達量siNC組
1.2.6 YWHAE-shRNA慢病毒干擾細胞蛋白表達功能的回復驗證 為了證明實驗篩獲的YWHAE-shRNA慢病毒對細胞YWHAE的干擾未出現off-target效應(脫靶效應),進一步采用干擾回復實驗進行驗證。
1.2.6.1 YWHAE點突變質粒pcDNA3.1-myc-YWHAEm的構建 根據GeneBank上公布的YWHAE基因(NM_006761.4),設計擴增YWHAE基因(768 bp)的引物。
上游:
YWHAE-F:5′-CCGCTCGAGGCCACCATGGATGA
TCGAGAGGATCTG-3′(下劃線為XhoI位點,序列5′端加上GCCACC增強翻譯效率)
下游:
YWHAE-R:5′-CGGGGTACCCTGATTTTCGTCTTC
CA-3′(下劃線為KpnI位點)
同時,設計含針對siYWHAE-5干擾序列的點突變(下劃線標記)。
正向:
Fm:5′-GGCGGCGAGGATAAACTGAAGATGATTC
GGGAATATCGGCAAA-3′
反向:
Rm:5′-CTTCAGTTTATCCTCGCCGCCCTTGTTTT
CTTCTTTCTGTTC-3′
引物由福州鉑尚生物技術有限公司合成。利用重疊延伸PCR法,以pGADT7-YWHAE(本實驗室保存)作為模板,使用引物YWHAE-F和Rm以及Fm和YWHAE-R分別擴增YWHAE基因上游及下游片段。擴增參數為:94 ℃預變性5 min;94 ℃變性30 s→55 ℃退火30 s→72 ℃延伸20 s(上游片段)或40 s(下游片段),共25個循環;最后72 ℃延伸10 min。以YWHAE基因上游及下游片段為模板,使用引物YWHAE-F和YWHAE-R擴增YWHAE點突變基因的全長(YWHAEm)。擴增參數為:94 ℃預變性5 min;94 ℃變性30 s→55 ℃退火30 s→72 ℃延伸1 min,共25個循環;最后72 ℃延伸10 min。進而將YWHAEm克隆入pcDNA3.1/myc-His(-)A載體中,構建pcDNA3.1-myc-YWHAEm表達質粒,轉化大腸桿菌感受態細胞DH5α,挑取重組陽性菌落抽提質粒行PCR、雙酶切及測序鑒定。
1.2.6.2 pcDNA3.1-myc-YWHAEm蛋白表達驗證及干擾回復驗證 用FuGENE 6 Transfection Reagent將pcDNA3.1-myc-YWHAEm分別轉染入AGS野生株、AGS-siNC和AGS-siYWHAE細胞株。48 h后提取細胞蛋白,BCA定量,進行Western-blot檢測目的蛋白的表達[一抗Myc-Tag (9B11) Mouse mAb,1∶1 000稀釋和anti-14-3-3ε(T-16);二抗goat anti-mouse IgG-AP;內參anti-β-actin],觀察在YWHAE-shRNA慢病毒干擾的AGS 細胞中YWHAE表達的回復效果。
1.2.7 MTS法檢測YWHAE-shRNA慢病毒對AGS細胞增殖的影響 將YWHAE沉默效果最好的AGS細胞株(AGS-siYWHAE)和對照組細胞株(AGS-siNC)分別以每孔2×103個細胞接種于96孔板,每組設3個復孔,連續培養5 d。實驗結束前1 h,吸棄細胞培養液,每孔加入100 μL培養液和20 μL Cell Titer 96 AQueous One Solution Reagent,在37 ℃、體積分數為0.05的CO2環境下孵育1 h。在酶標儀上用490 nm波長測定各孔吸光度(OD)值,以空白孔調零,實驗重復5次。以OD值為縱軸、作用時間為橫軸繪制AGS細胞增殖曲線。
增殖抑制率=(1-ODsiYWHAE組/ODsiNC組)×100%
2 結 果
2.1 重組慢病毒pLL3.7-YWHAE-shRNA表達質粒的鑒定 慢病毒載體pLL3.7含有限制性核酸內切酶XhoI和HpaI的酶切位點,用于插入退火產物。5對針對人YWHAE基因的shRNA序列和1條無義的陰性對照序列的正義鏈和反義鏈,退火形成雙鏈DNA oligo,各自退火產物的兩端中均有一端為XhoI的粘性末端,另一端為平齊末端。故將退火產物克隆入pLL3.7載體中,獲得的重組質粒pLL3.7-YWHAE(或NC)-shRNA中還保留XhoI位點,而HpaI位點已被破壞,借助這種特性可對初篩的重組質粒進行酶切鑒定:即重組構建成功后,當質粒用EcoR I和XbaI酶切,可切開得到約1 800 bp的片段;而用EcoR I和HpaI酶切則無法獲得雙酶切效果(圖1)。將經雙酶切初篩的質粒進一步送測序驗證,成功構建了具有正確序列的重組質粒,分別為pLL3.7-siYWHAE-1,pLL3.7-siYWHAE-2,pLL3.7-siYWHAE-3,pLL3.7-siYWHAE-4,pLL3.7-siYWHAE-5和pLL3.7-siNC。
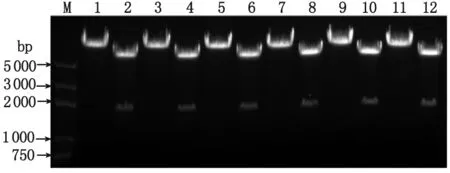
1,2:pLL3.7-siYWHAE-1質粒EcoR I/Hpa I及EcoR I/Xba I雙酶切;3,4:pLL3.7-siYWHAE-2質粒EcoR I/Hpa I及EcoR I/Xba I雙酶切;5,6:pLL3.7-siYWHAE-3質粒EcoR I/Hpa I及EcoR I/Xba I雙酶切;7,8:pLL3.7-siYWHAE-4質粒EcoR I/Hpa I及EcoR I/Xba I雙酶切;9,10:pLL3.7-siYWHAE-5質粒EcoR I/Hpa I及EcoR I/Xba I雙酶切;11,12:pLL3.7-siNC質粒EcoR I/Hpa I及EcoR I/Xba I雙酶切;M:5 kb DNA Marker.圖1 重組質粒pLL3.7-YWHAE-shRNA雙酶切鑒定Fig 1 Double digestion analysis of the recombinant plasmid pLL3.7-YWHAE-shRNA
2.2 YWHAE-shRNA 慢病毒干擾細胞蛋白表達功能的鑒定 慢病毒表達質粒(pLL3.7-siYWHAE-1,pLL3.7-siYWHAE-2,pLL3.7-siYWHAE-3,pLL3.7-siYWHAE-4,pLL3.7-siYWHAE-5和pLL3.7-siRNA-NC)和包裝質粒PHR、包膜質粒VSVG轉染293T細胞,轉染24 h后,即可觀察到細胞病理現象。在轉染后48 h收集病毒感染AGS細胞,用含Puromycine的完全培養液篩選細胞。篩選結束后,提細胞總蛋白進行Western-blot驗證。結果顯示,5對針對人YWHAE基因的shRNA序列構建的慢病毒中,pLL3.7-siYWHAE-5包裝成的慢病毒抑制AGS細胞YWHAE蛋白表達的效果最明顯,僅為對照組表達量的(0.269±0.083)倍(圖2),該穩定細胞株命名為AGS-siYWHAE細胞。
2.3 重組YWHAE點突變pcDNA3.1-myc-YWHAEm表達質粒的鑒定 以pGADT7-YWHAE為模板,利用重疊延伸PCR法擴增定點突變的YWHAEm基因,并克隆入pcDNA3.1/myc-His(-)A載體,獲得的重組質粒經PCR鑒定,可見約768 bp的DNA條帶(圖3)。重組質粒經XhoI和KpnI雙酶切后,可見約5 500和768 bp2條DNA條帶,分別與載體質粒和目的基因大小相符(圖3),進一步測序證實YWHAEm基因正確插入pcDNA3.1/myc-His(-)A載體中。
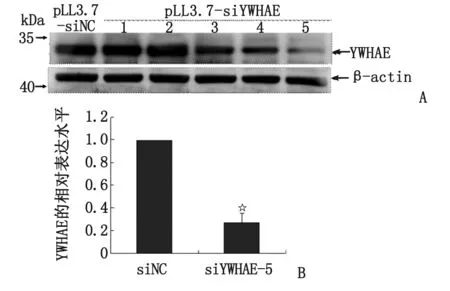
n=3. A:Western-blot鑒定YWHAE-shRNA慢病毒對AGS細胞YWHAE蛋白表達的影響;B:AGS-siYWHAE細胞YWHAE蛋白相對表達水平. 與siNC組比較,☆:P<0.05.圖2 YWHAE-shRNA慢病毒對AGS細胞YWHAE蛋白表達的影響Fig 2 Effect of YWHAE-shRNA lentivirus on the expression of YWHAE protein in AGS cells
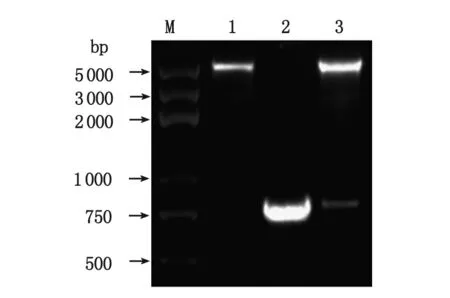
1:pcDNA3.1/myc-His(-)A質粒Xho I/Kpn I雙酶切;2:以pcDNA3.1-myc-YWHAEm為模板PCR擴增點突變的YWHAE基因;3:pcDNA3.1-myc-YWHAEm質粒Xho I/Kpn I雙酶切;M:5 kb DNA Marker.圖3 pcDNA3.1-myc-YWHAEm質粒的PCR及雙酶切鑒定Fig 3 PCR identification and double digestion analysis of the recombinant plasmid pcDNA3.1-myc-YWHAEm
2.4 pcDNA3.1-myc-YWHAEm對YWHAE- shRNA慢病毒干擾的回復驗證 將pcDNA3.1-myc-YWHAEm和pcDNA3.1/myc-His(-)A(對照)分別轉染AGS細胞,經Western-blot顯示,pcDNA3.1-myc-YWHAEm轉染組用Myc-Tag (9B11) Mouse mAb可檢測到目的蛋白條帶,分子量大小與預期相符(約38 kDa)(圖4);而對照組未見特異性條帶,即YWHAE點突變質粒pcDNA3.1-myc-YWHAEm可經轉染在AGS細胞中表達YWHAE。將pcDNA3.1-myc-YWHAEm質粒轉染AGS-siYWHAE細胞,經Western-blot顯示,與轉染pcDNA3.1/myc-His(-)A的AGS-siYWHAE細胞比較,轉染后AGS-siYWHAE細胞中YWHAE表達明顯上調,與AGS-siNC細胞內源性YWHAE相近(圖5)。表明YWHAE-shRNA慢病毒對AGS細胞表達YWHAE的干擾實驗是成功的,未出現off-target效應,細胞可經轉染pcDNA3.1-myc-YWHAEm重組質粒回復干擾。
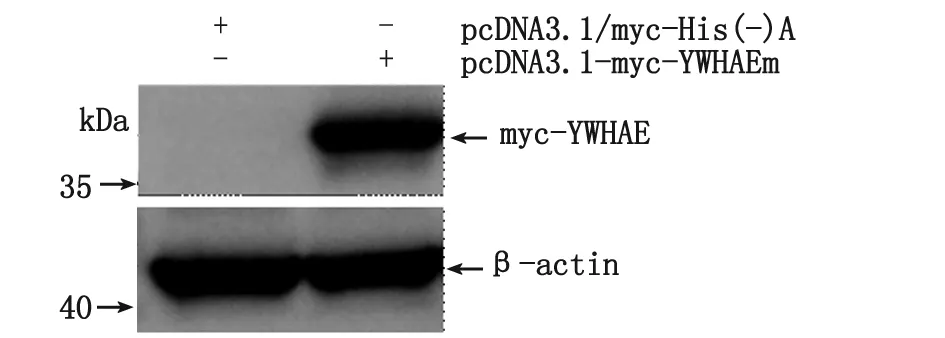
圖4 YWHAEm蛋白在AGS細胞中的表達Fig 4 YWHAEm protein expression in AGS cells
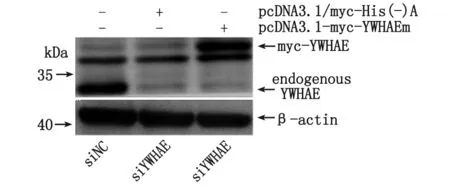
圖5 YWHAEm蛋白在AGS-siYWHAE細胞中的回復表達Fig 5 YWHAEm protein re-established expression in AGS-siYWHAE cells
2.5 YWHAE-shRNA 慢病毒對AGS細胞增殖功能的影響 用MTS比色法檢測AGS-siYWHAE和AGS-siNC(對照)1~5 d的細胞增殖情況,結果顯示,AGS-siYWHAE組與AGS-siNC組比較,從第2天開始,細胞增殖就出現差異,第3~5天這種差異逐漸加大,增殖抑制率分別為23.6%,23.6%和20.8%,AGS-siYWHAE組的增殖受到顯著抑制(圖6)。表明YWHAE-shRNA慢病毒感染使AGS細胞YWHAE表達下調后,AGS細胞的增殖能力明顯受到抑制。
3 討 論
本研究成功構建了YWHAE-shRNA慢病毒表達質粒。在構建YWHAE-shRNA慢病毒表達質粒前期,考慮到在目的基因上選擇不同起始位點的片段為模板所構建的RNAi載體對目的基因的抑制率可能會不同,課題組分別針對人YWHAE基因的5′非編碼區(UTR5)和開放讀碼框(ORF)設計了5個基因片段來構建相應表達質粒。篩選結果發現,針對UTR5設計的2個片段干擾效果不明顯,推測可能UTR5存在豐富的調控蛋白結合區域,而這些調控蛋白的結合可能影響到RNA誘導沉默復合體(RNA-induced silencing complex, RISC)與mRNA的結合,從而影響了siRNA的干擾效果;針對ORF設計的3個片段均能達到一定的下調靶基因表達的作用,并且片段5的干擾抑制效率最高[YWHAE的相對表達量為對照組的(0.269±0.083)倍]。進而課題組設計含針對siYWHAE-5干擾序列的點突變引物,構建了YWHAE點突變(同義突變)表達質粒,并將后者轉染篩獲的細胞株,順利回復了YWHAE的表達,表明課題組篩獲的YWHAE- shRNA慢病毒對細胞YWHAE的干擾未出現脫靶效應,成功構建了由shRNA慢病毒表達質粒介導的YWHAE表達下調的AGS胃癌細胞模型,可用于后續的實驗。
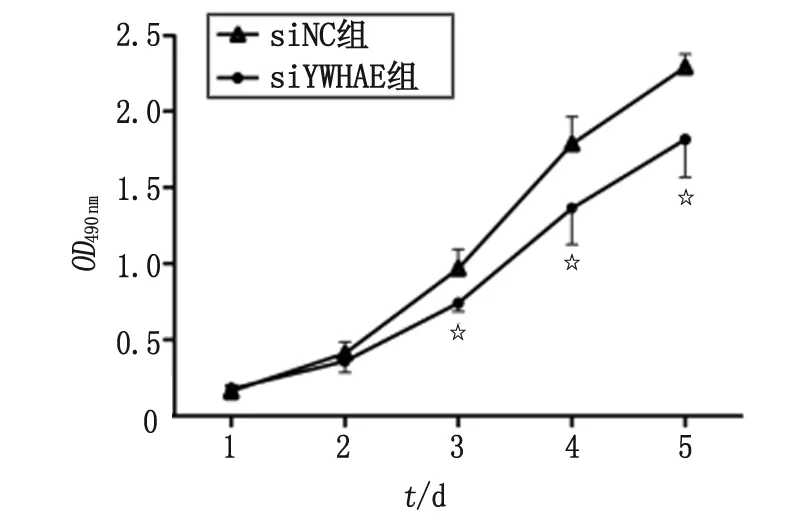
n=5. YWHAE-shRNA慢病毒感染使AGS細胞的增殖能力明顯受到抑制. 與siNC組比較,☆:P<0.05.圖6 YWHAE-shRNA慢病毒對AGS細胞增殖的影響Fig 6 Effect of YWHAE-shRNA lentivirus on the proliferation of AGS cells
與Gong等采用shRNA質粒轉染篩選YWHAE表達下調的SGC7901細胞相比[5],本研究采用慢病毒載體感染篩選YWHAE表達下調的AGS細胞具有自身的優勢:(1)慢病毒載體源于人類免疫缺陷病毒-1,能感染非分裂期細胞和分裂期細胞,宿主范圍廣;(2)病毒感染細胞的效率通常比質粒轉染細胞的效率高;(3)當感染細胞時,病毒的基因可整合到宿主細胞的基因組中,隨細胞分裂傳給子代,表達穩定;(4)病毒自身的致病基因已被剔除,難以重組野生病毒株,應用較為安全。因此,本研究用慢病毒載體表達YWHAE-siRNA能在胃癌細胞中高效、安全、穩定持久地發揮干擾下調YWHAE基因表達的作用,并且不僅可用于后續的細胞實驗,還可用于動物體內的實驗研究。此外,Gong等采用的shRNA質粒為商品化的試劑,不僅成本高,而且具體序列未公布,使得干擾回復實驗的開展受限[5]。
在YWHAE與胃癌的相關研究中,Yan等通過在SGC7901胃癌細胞中過表達YWHAE,發現YWHAE可激活ERK/MAPK信號通路,導致細胞增殖[7]。Gong等在下調YWHAE表達的SGC7901細胞中發現了抑制細胞增殖的現象[5]。本課題組借助MTS法檢測,發現課題組構建的YWHAE-shRNA慢病毒介導AGS細胞YWHAE表達下調后,細胞的增殖能力也明顯受到抑制。由此表明,YWHAE在胃癌細胞中具有促進細胞增殖的功能,很可能促進了胃癌的發生發展。但是關于YWHAE在胃癌發生發展中的作用目前仍觀點不一。例如,Leal團隊研究發現,YWHAE蛋白在胃癌組織中的表達明顯的低于癌旁組織,提示YWHAE可能是種抑癌蛋白[6]。遺憾的是,該團隊并未進一步探討YWHAE在胃癌細胞內的功能機制。在胃癌的發生發展中不僅發生細胞增殖異常,而且在細胞凋亡、侵襲轉移、血管生成等多方面都可能發生異常,YWHAE是促癌還是抑癌,則還有待更多體內外實驗的深入驗證。
本研究成功構建YWHAE-shRNA慢病毒表達質粒,建立YWHAE表達下調的AGS胃癌細胞模型,探明YWHAE表達下調對AGS細胞增殖的影響,為調控YWHAE在胃癌細胞內的功能研究和動物體內實驗的開展奠定了基礎。
[1] Van Cutsem E, Sagaert X, Topal B,etal. Gastric cancer[J].Lancet, 2016,388(10060):2654-2664.
[2] Aitken A. 14-3-3 proteins: a historic overview [J].SeminCancerBiol, 2006,16(3):162-172.
[3] Chen S Y, Zhang R G, Duan G C. Pathogenic mechanisms of the oncoprotein CagA inH.pylori-induced gastric cancer [J].OncolRep, 2016,36(6):3087-3094.
[4] Nagappan A, Park H S, Park K I,etal.Helicobacterpyloriinfection combined with DENA revealed altered expression of p53 and 14-3-3 isoforms in Gulo-/-mice [J].ChemBiolInteract, 2013,206(2):143-152.
[5] Gong X, Yan L, Gu H,etal. 14-3-3ε functions as an oncogene in SGC7901 gastric cancer cells through involvement of cyclin E and p27kip1[J].MolMedRep, 2014,10(6):3145-3150.
[6] Leal M F, Calcagno D Q, Demachki S,etal. Clinical implication of 14-3-3 epsilon expression in gastric cancer [J].WorldJGastroenterol, 2012,18(13):1531-1537.
[7] Yan L, Gu H, Li J,etal. RKIP and 14-3-3ε exert an opposite effect on human gastric cancer cells SGC7901 by regulating the ERK/MAPK pathway differently [J].DigDisSci, 2013,58(2):389-396.
(編輯:何佳鳳)
Construction and Function Identification of Short Hairpin RNA Lentiviral Expression Plasmid Targeting YWHAE
ZHANG Xiaoyan, WEN Chunyan, LYU Yuhua, CHEN Hao, SHE Feifei
Key Laboratory of Ministry of Education for Gastrointestinal Cancer, Fujian Key Laboratory of Tumor Microbiology, Fujian Medical University, Fuzhou 350122, China
Objective To construct a short hairpin RNA (shRNA) lentiviral expression plasmid targeting YWHAE (Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, epsilon) gene and then detect the effect of shRNA on the proliferation of AGS cells. Methods Five shRNA interference sequences targeting YWHAE gene were designed, synthesized and cloned into the pLentiLox3.7(pLL3.7) expression vector. Then the expression plasmids, PHR and VSVG were co-transfected into 293T cells using calcium phosphate to package virus. AGS cells were infected with the extracted virus, and screened with Puromycine. Silencing effect was verified via Western-blot. The lentiviral expression plasmid with the best silence of YWHAE was selected, then YWHAE gene with point mutation was constructed by overlap-extension PCR, and cloned into pcDNA3.1/myc-His(-)A vector. The pcDNA3.1-myc-YWHAEm was transfected into AGS cells with the best silence of YWHAE, then re-expression of YWHAE was detected by Western-blot. MTS assay was used to detect the proliferation of AGS cells. Results Among the five packaged lentivirus, the pLL3.7-siYWHAE-5 lentivirus were selected with the best effect. The relative level of YWHAE protein expression [(0.269±0.0 83)-fold] of AGS cells infected with the selected virus was significantly lower than that of AGS cells infected with the pLL3.7-siNC lentivirus. After pcDNA3.1-myc-YWHAEm was transfected into the AGS cells with the best silence of YWHAE, however YWHAE protein expression were re-established. Compared to negative control group (infected with NC-shRNA lentivirus), cell proliferation in YWHAE-shRNA group was significantly inhibited. Conclusion The recombinant shRNA lentivirus targeting the YWHAE gene was constructed successfully, and the AGS cells with knock-down of YWHAE were obtained. Knocking down YWHAE gene has noticeable effects on the proliferation inhibition of AGS cells, which implies YWHAE may be a putative oncoprotein in AGS cells.
lentivirus; RNA interference; stomach neoplasms; cell proliferation
2017-03-07
國家自然科學基金(81271784);福建省教育廳項目(JA12150);福建醫科大學重大科研項目基金(09ZD018);福建醫科大學苗圃科研基金(2010MP029)
福建醫科大學 消化道惡性腫瘤教育部重點實驗室,福建省腫瘤微生物重點實驗室,福州 350122
張曉艷,女,講師,醫學博士
佘菲菲. Email: shefeifei@yeah.net
R394.2; R735.2
A
1672-4194(2017)03-0139-07
猜你喜歡
昆明醫科大學學報(2022年1期)2022-02-28 07:43:36
昆明醫科大學學報(2021年5期)2021-07-22 07:32:22
基層中醫藥(2020年2期)2020-07-27 02:46:06
中國組織化學與細胞化學雜志(2016年3期)2016-02-27 11:15:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:46
中外醫療(2015年18期)2016-01-04 06:51:55
吉林大學學報(醫學版)(2015年4期)2015-12-17 07:48:25
醫學研究雜志(2015年6期)2015-07-01 17:40:49
醫學研究雜志(2015年9期)2015-07-01 17:28:27
中國當代醫藥(2015年20期)2015-03-01 02:04:29
