凝結芽孢桿菌RY237β-半乳糖苷酶酶學性質研究
2017-08-09 02:47:53李文婷王翠萍朱友峰王世榮岳壽松山東省農業科學院生物技術研究中心山東濟南25000天津科技大學食品工程與生物技術學院天津300450
食品工業科技 2017年14期
關鍵詞:研究
李文婷,邊 斐,王翠萍,陳 高,張 燕,朱友峰,王世榮,岳壽松,*(.山東省農業科學院生物技術研究中心,山東濟南 25000; 2.天津科技大學食品工程與生物技術學院,天津 300450)
?
凝結芽孢桿菌RY237β-半乳糖苷酶酶學性質研究
李文婷1,2,邊 斐1,王翠萍1,陳 高1,張 燕1,朱友峰1,王世榮1,岳壽松1,*
(1.山東省農業科學院生物技術研究中心,山東濟南 250100; 2.天津科技大學食品工程與生物技術學院,天津 300450)
為研究乳品中具有應用價值的乳糖酶,以乳糖為唯一碳源,用鄰硝基苯酚-β-D-半乳糖苷(ONPG)法,從產乳酸的細菌中篩選出了產乳糖酶活力高的菌株RY237,其活性達4.98 U/mL,鑒定為凝結芽孢桿菌。研究了乳糖酶的酶學性質,該酶最適反應溫度和最適pH分別為50 ℃和6.0。該酶在溫度40~50 ℃具有良好的穩定性;pH5.5~8.0表現穩定,pH7.0出現酶活峰值。金屬離子Ca2+、K+、Zn2+、Mn2+、Na+、Mg2+對酶活具有抑制作用,Cu2+對酶活具有完全抑制作用,EDTA對酶活具有激活作用。凝結芽孢桿菌RY237乳糖酶活性高、性能優良,具有應用價值。
凝結芽孢桿菌,分子鑒定,β-半乳糖苷酶,酶學性質
乳糖酶又稱為β-半乳糖苷酶,廣泛存在于植物、微生物以及哺乳動物腸道中。乳糖酶可催化乳糖水解為半乳糖和葡萄糖,對解決人體普遍存在的乳糖不耐具有重要意義,引起了廣泛關注[1-4]。目前只有來源于微生物的乳糖酶有工業應用價值。產乳糖酶微生物種類具有多樣性,包括曲霉菌、酵母菌、放線菌、芽孢桿菌、乳酸菌等[5-9]。不同來源的乳糖酶性能有很大不同,用途也不一樣。霉菌產生的乳糖酶最適pH偏酸性(pH2.5~5.0),可應用于酸性乳清和奶酪的水解;酵母菌和細菌所產乳糖酶最適pH近中性(分別為pH6~7和pH6.5~7.5),適于牛乳和鮮乳清的水解[10]。
凝結芽孢桿菌(Bacilluscoagulans)既能產乳酸又能形成芽孢,是一種重要的益生菌。近年有凝結芽孢桿菌產乳糖酶研究的報道[11-14]。Batra等研究了從溫泉中分離的凝結芽孢桿菌RCS3乳糖酶性質,該菌在50 ℃產酶量最大,65 ℃酶活性最高,酶的最適pH6~7,在pH5~8范圍酶活表現穩定,該酶良好的性能具有在乳品業開發的潛力[13]。宋春雷等對凝結芽孢菌桿菌L42產乳糖酶條件進行優化,總酶活力提高了10.8倍,達到7.1 U/mL,L42產酶最佳發酵溫度為50 ℃,該酶的最適反應溫度為65 ℃,最適pH6.0[12]。Levin等報道,凝結芽孢桿菌L4的最適溫度為55 ℃,其最適pH在30 ℃和55 ℃分別為6.0和6.5[14]。由此可見,凝結芽孢桿菌乳糖酶性質因菌株不同特點不同。因此,篩選具有高活性乳糖酶的凝結芽孢桿菌資源,研究乳糖酶性質,具有理論和實踐意義。
本研究對實驗室保存的產乳酸菌株進行乳糖酶篩選,鑒定了產乳糖酶能力強的菌株,并研究了乳糖酶性質,為該菌種的開發利用提供理論依據。
1 材料與方法
1.1 材料與儀器
產乳酸菌株LP1119、LP777、LP738、LC544、LC 868、LC 727、RY 237和大腸桿菌(Escherichiacoli)DH5α本實驗室保存;pMD18-T載體 TakaRa公司;pET28a(+) Novagen公司;鄰硝基苯酚-β-D-吡喃半乳糖苷(ONPG,色譜純) Bio Basic Inc公司(上海生物工程有限公司分裝);鄰硝基苯酚(ONP,色譜純) 國藥集團化學試劑有限公司;ONPG磷酸鹽緩沖液 0.6025 g ONPG溶于100 mL磷酸鹽緩沖溶液;pH7.0磷酸鹽緩沖溶液 13.6 g磷酸二氫鈉、0.06 g硫酸鎂、0.126 g氯化錳溶于1L水;檸檬酸-磷酸氫二鈉緩沖溶液 0.1 mol/L檸檬酸與0.2 mol/L磷酸氫二鈉混合;0.1 mol/L醋酸-醋酸鈉緩沖溶液 0.2 mol/L的醋酸和0.3 mol/L的醋酸鈉混合后稀釋一倍;0.05 mol/L甘氨酸-氫氧化鈉緩沖溶液 0.2 mol/L的甘氨酸和0.2 mol/L的氫氧化鈉混合;乳糖MRS(L-MRS)培養基(g/L) 牛肉膏10,酵母膏5,蛋白胨10,檸檬酸氫二銨2,磷酸氫二鉀2,乳糖20,MnSO4·H2O 0.23,MgSO4·7H2O 0.58,無水乙酸鈉5 g,吐溫-80 1 mL,pH6.4。
冷凍高速離心機 美國Thermo Fisher公司;PCR儀 日本Takara公司;UP200s型超聲波破碎儀 德國Dr.Hielscher公司;TSQ-280型立式搖床 上海精宏實驗設備有限公司;723型可見分光光度計 上海光譜儀器有限公司。
1.2 標準曲線繪制
取6支試管分別加入200 μg/mL的ONP溶液0、0.2、0.4、0.6、0.8、1.0 mL,用磷酸鹽緩沖液補足至1 mL。37 ℃水浴10 min。分別加入4 mL 0.5 mol/L Na2CO3。以第一只試管為空白,測OD420。以ONP濃度(mmol/L)為縱坐標,OD420為橫坐標繪制標準曲線。
1.3β-D-半乳糖苷酶活性的測定
1.3.1 酶的發酵與提取 將菌株接種于L-MRS培養基,37 ℃、150 r/min振蕩培養24 h。取培養液4 mL,4 ℃、7000 r/min離心15 min,棄上清。菌體用0.1 mol/L磷酸鹽緩沖溶液(pH7.0)洗滌、7000 r/min離心15 min。重復兩次。收集菌體并用0.1 mol/L磷酸鹽緩沖溶液(pH7.0)添加至4 mL,60 W超聲破碎15 min后于4 ℃、12000 r/min離心5 min,取上清液測酶活。
1.3.2 酶活性測定 取1 mL酶提取液,37 ℃水浴預熱5 min,加入已預熱至37 ℃的含20 mmol/L ONPG磷酸鹽緩沖液1.0 mL,37 ℃水浴反應10 min,然后立即加入3 mL 0.5 mol/L Na2CO3終止反應,測OD420。以65 ℃加熱失活的酶液作為空白對照。一個酶活力單位(U)定義為:37 ℃條件下,1 min水解產生1 μmol ONP所需要的酶量。乳糖酶活力計算公式如下:
X=(C×20×N)/10,其中,X為乳糖酶活力;C為標準曲線上對應的ONP濃度;N為稀釋倍數。
1.4 菌株生理生化反應
生理生化特性測定參照《常見細菌系統鑒定手冊》中的方法[15]。
1.5 16S rDNA基因的克隆、測序
菌株基因組DNA提取參照《精編分子生物學指南》[16]的方法略有改動。擴增細菌16S rDNA通用引物。上游P1:5′-AGAGTTTGATCCTGGCTCAG-3,下游P2:5′-TACGGCTACCTTGTTACGACT-3。以菌株基因組為模板,P1、P2為引物,PCR擴增16S rDNA。PCR擴增條件為:95 ℃ 5 min;94 ℃ 40 s,55 ℃ 40 s,72 ℃ 2 min,30個循環;最后72 ℃ 10 min。PCR擴增產物經膠回收后克隆至pMD18-T載體,轉化E.coliDH5α,選取正確的陽性克隆送上海生工測序。
1.6 系統發育樹構建
將16S rDNA序列與GenBank里的已知核酸序列進行BLAST分析。選取與該序列同源性較高的已知菌株的16S rDNA序列,利用MEGA6.0軟件進行多序列比和系統發育樹構建。
1.7β-D-半乳糖苷酶酶學性能
1.7.1 最適反應溫度與熱穩定性 最適反應溫度:200 μL磷酸鹽緩沖液(pH7.0)稀釋的酶液,30、40、50、60、70 ℃保溫5 min,同時將200 μL ONPG溶液依次在相同溫度下保溫5 min,將ONPG加入到酶液中,在各溫度下反應10 min,加入600 μL Na2CO3終止反應。計算各時間酶活與最高酶活的百分比,確定酶最適反應溫度。
酶的熱穩定性:200 μL磷酸鹽緩沖液稀釋的酶液,40、50、60、65 ℃保溫0、10、20、30、40、50 min。同時將200 μL ONPG溶液依次在相同溫度下進行保溫,將ONPG加入到粗酶液中,37 ℃反應10 min,加入600 μL Na2CO3終止反應,測OD420,以65 ℃加熱失活的酶液作為空白對照,以0時間酶活為100%,計算各時間酶活與最高酶活百分比。
1.7.2 最適pH和pH穩定性 最適pH:在最適溫度下,按1.3.2酶活測定方法,測定pH4.0、4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5、9.0、9.5和10.0條件下酶活性。計算各pH酶活與最高酶活的百分比,確定酶最適反應pH。
pH穩定性:將酶液分別置于pH為4.0、4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0、8.5、9.0、9.5和10.0的緩沖溶液中,37 ℃放置18 h,測定各pH條件下的酶活,以最高酶活為100%,計算各pH酶活與最高酶活百分比,測定酶的pH穩定性[17-18]。
1.7.3 金屬離子和EDTA對酶活的影響 在酶促反應體系中加入1 mol/L的Mn2+、Mg2+、Na+、K+、Ca2+、Cu2+、Zn2+、EDTA溶液,使最終反應體系中金屬離子的濃度為1 mmol/L,測定酶活力以加熱失活的酶液作為空白對照。不加金屬離子溶液的酶活作為對照,計算相對酶活。
2 結果與分析
2.1 ONPG標準曲線
以光密度OD420為橫坐標,ONP濃度為縱坐標繪制乳糖酶活標準曲線,相關系數R2=0.998(圖1),說明該標準曲線可用于乳糖酶的測定。
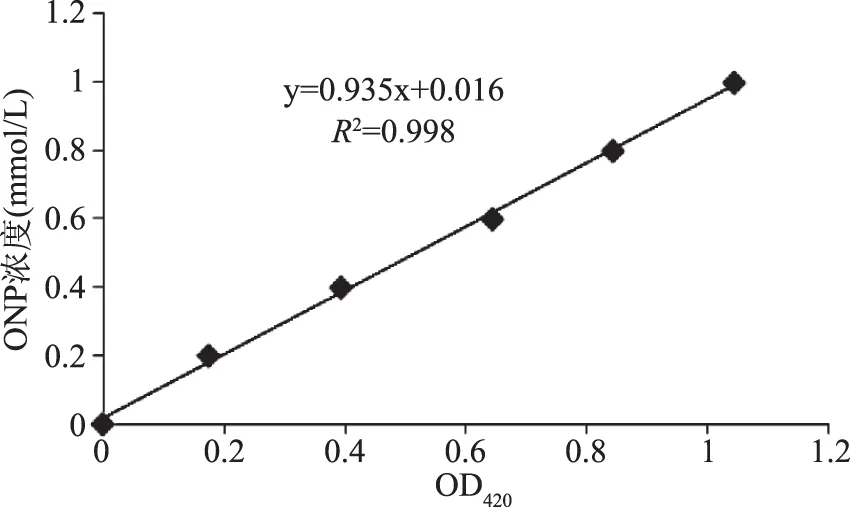
圖1 β-半乳糖苷酶活性測定標準曲線Fig.1 Standard curve of β-galactosidase
2.2 產乳酸菌株的乳糖酶活性

圖3 基于16S rDNA序列的系統發育樹Fig.3 Unrooted phylogenetic tree based on the 16S rDAN sequence
將RY237、LC777、LP272、LP119、LC868,LP554和LC738菌株L-MRS培養基液體培養24 h,由于不同菌株培養液的光密度OD600存在差異,以光密度最低的菌株OD值0.7為標準,把各菌株培養液的OD600調節為0.7,然后利用1.3.1中酶的提取方法收集菌體,磷酸緩沖液洗滌兩次,重新懸浮,超聲波破碎,離心取上清液測定各菌株酶活。結果表明,各菌株酶活高低依次為RY237(4.98 U/mL)、LC777(0.34 U/mL)、LP272(0.28 U/mL)、LP119(0.11 U/mL)、LC868(0.08 U/mL),LP554和LC738沒有檢測到乳糖酶活性(圖2)。關于乳酸菌乳糖酶報道較多,酶活一般在5 U/mL以下[17-20],RY237乳糖酶活性明顯高于已報道的乳糖酶,說明具有潛在的開發應用價值。
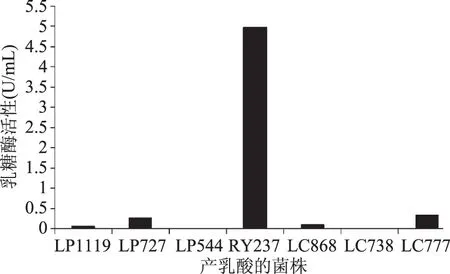
圖2 不同菌株乳糖酶活性Fig.2 Activity of β-galactosidase in different strains
2.3 菌株鑒定
2.3.1 生理生化特征 菌株生長特征為:可以在15~55 ℃下生長,NaCl 7%時生長被抑制。生理生化反應結果表明,RY237菌株能發酵葡萄糖產酸,明膠不液化,能水解酪素和淀粉,不能利用檸檬酸鹽和丙酸鹽,V-P實驗陽性,接觸酶和過氧化氫酶陽性,卵黃反應和吲哚產生實驗陰性,可發酵甘露糖、阿拉伯糖。菌株RY237生長特征和生理生化特性與凝結芽孢桿菌的特征吻合[15]。

表1 菌株RY237生理生化特征Table 1 Biological and physiological characteristics of strain RY 237
注:“+”,陽性反應;“-”,陰性反應。
2.3.2 分子鑒定 測序結果表明,RY237菌株16S rDNA序列全長1450 bp。將該序列提交NCBI進行BLAST,結果發現其與凝結芽孢桿菌(Bacilluscoagulans)同源性最高,達99%。根據同源性比對結果,選出相關性較高和芽孢桿菌屬內相關菌株的16S rDNA序列構建系統發育樹,結果顯示其與Bacilluscoagulans聚為一族(圖3)。結合生理生化特征及16S rDNA分子鑒定結果,將RY237鑒定為凝結芽孢桿菌,命名為BacilluscoagulansRY237。
2.4 RY237乳糖酶最適反應溫度與熱穩定性
酶最適反應溫度是酶的重要性能,乳糖酶商業應用中一般要求酶的最適溫度為37 ℃或更高[21]。本研究在pH7.0條件下,按照1.3.1對RY327提取乳糖酶,測定30~70 ℃范圍內乳糖酶活性變化。結果表明,酶活性隨溫度升高而先增加后減少,40 ℃酶活上升到較高水平(最高值的90.6%),50 ℃酶活最高,60 ℃酶活為最高值的72.8%,之后快速下降,至70 ℃已檢測不到酶活(圖4)。由此可以確定菌株R237乳糖酶的最適溫度為50 ℃。該值與已報道的凝結芽孢桿菌乳糖酶的最適反應溫度有所不同,鄭兆娟等報道凝結芽孢菌桿菌L42最適反應溫度為65 ℃[11],另有最適反應溫度為63 ℃[13]和55 ℃[14]的報道。說明RY327乳糖酶的性能與已報道的菌株性能不同。
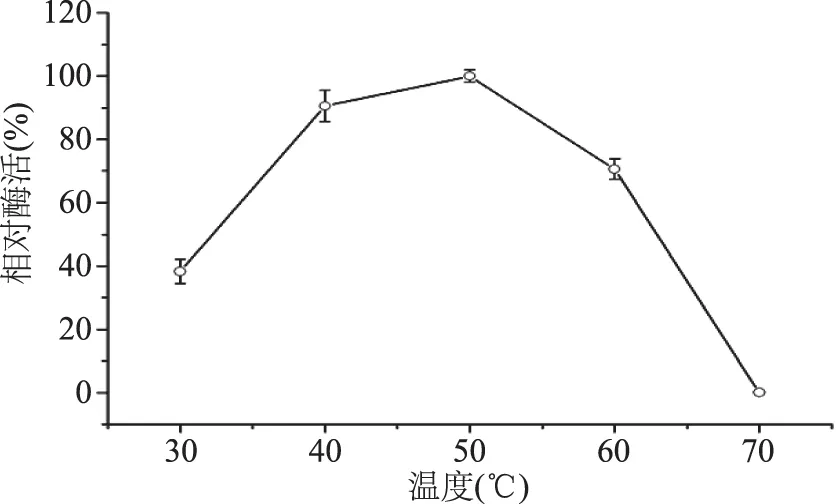
圖4 不同溫度下乳糖酶活性變化Fig.4 Effect of temperature on activity of β-galactosidase
將酶分別置于40、50、60、65 ℃不同時間(10~50 min)測定酶活性。以最高值為100%,計算各溫度不同保溫時間的相對酶活。結果表明,乳糖酶活性在40 ℃和50 ℃具有較好的穩定性,保溫30 min,酶活仍保持最高值90%以上(分別為92.1%和90.1%)。60 ℃穩定性下降,保溫30 min,酶活只有最高值的62.8%;65 ℃酶活急劇下降,保溫20 min酶活僅為最高值的2.5%,至30 min已檢測不到酶活,說明該酶對熱敏感(圖5)。

圖5 乳糖酶熱穩定性Fig.5 Termostability of β-galactosidase
2.5 RY237乳糖酶最適pH與pH穩定性
按照1.3對RY327提取乳糖酶和乳糖酶的活性測定,圖6表明酶最適pH為6.0。乳糖酶最適pH決定了其不同用途,霉菌產生的乳糖酶最適pH偏酸性(pH2.5~5.0),可應用于酸性乳清和奶酪的水解;酵母菌和細菌所產乳糖酶的最適pH近中性(分別為pH6~7和pH6.5~7.5),適于牛乳(pH6.6)和鮮乳清(pH6.1)的水解[10]。該結果說明凝結芽孢桿菌RY237在乳制品水解領域具有應用價值。
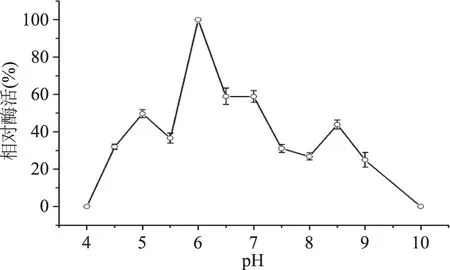
圖6 不同pH條件下乳糖酶酶活變化Fig.6 Effect of pH on activity of β-galactosidase
將酶液在不同的pH緩沖溶液中37 ℃保溫18 h,測定各pH條件下的酶活性,計算相對酶活。結果表明,酶在pH7.0酶活最高,最穩定。37 ℃、pH5.5~8.5維持18 h,酶活保持80%以上(圖7),說明該酶在pH5.5~8.0范圍內具有較高的穩定性,該酶pH適應范圍寬泛,具有潛在的應用價值。
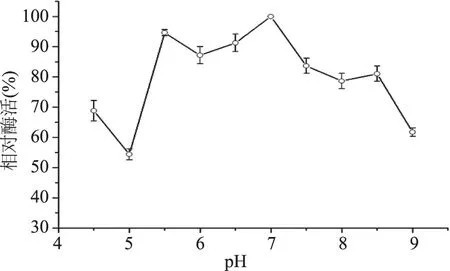
圖7 乳糖酶pH穩定性Fig.7 pH stability of β-galactosidase
2.6 金屬離子和EDTA對乳糖酶活性的影響
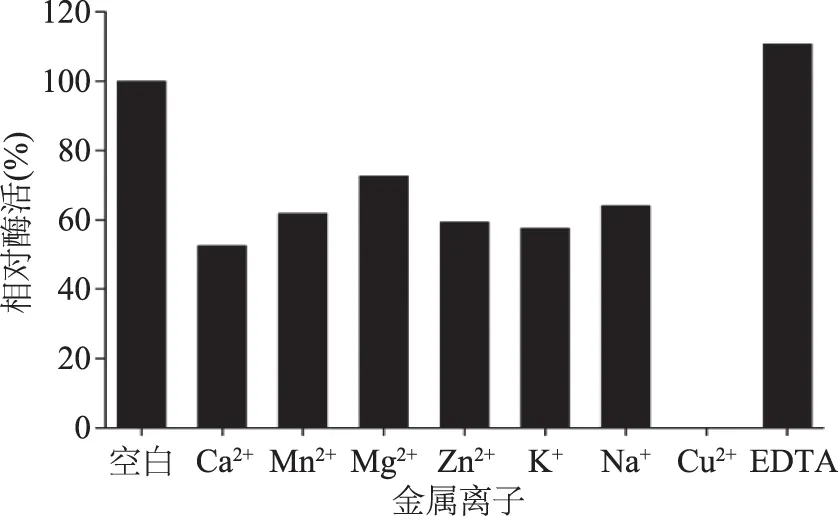
圖8 金屬離子和EDTA對乳糖酶活力的影響Fig.8 Effect of metalic ions and EDTA on β-galactosidase activity
金屬離子對酶活具有激活或抑制作用,按照1.3.1對RY327提取乳糖酶,測定了金屬離子Mn2+、Mg2+、Na+、K+、Ca2+、Cu2+、Zn2+對乳糖酶活性的影響。結果表明,上述7種金屬離子對酶活均有抑制作用,其中Cu2+對酶活具有完全抑制作用,剩余酶活依次分別為Ca2+(52.6%)、K+(57.6%)、Zn2+(59.4%)、Mn2+(61.8%)、Na+(64.1%)、Mg2+(72.7%)。EDTA對乳糖酶活具有激活作用(110.8%),可能在于EDTA對酶活性位點具有保護作用(圖8)。高曉峰研究植物乳桿菌產生的乳糖酶,金屬離子Mg2+、Mn2+對酶活性具有顯著的激活作用,而Cu2+、EDTA對酶活有較強的抑制作用[22]。說明酶的來源不同,性能不同。上述結果可作為酶活性調節與控制的理論依據。
3 結論
本研究以乳糖為唯一碳源,利用ONPG作為底物,從產乳酸的細菌中篩選出了產乳糖酶活力高的菌株RY237,鑒定為凝結芽孢桿菌(Bacilluscoagulans)RY237。研究了RY237乳糖酶的性質,結果表明,該酶在溫度40~50 ℃具有較高的酶活性(酶活為90.6%~100%),最適反應溫度為50 ℃,低于已報道的凝結芽孢桿菌乳糖酶的最適反應溫度,而且該酶在40~50 ℃具有很強的穩定性,說明其在高酶活區保持穩定的性能,具備良好生產潛力。該酶的最適pH為6.0,7.0最穩定,適于牛乳和乳清的水解。金屬離子對酶活抑制作用的影響為Cu2+>Ca2+>K+>Zn2+>Mn2+>Na+>Mg2+,Cu2+對酶活具有完全抑制作用,EDTA對乳糖酶活具有激活作用。因此,凝結芽孢桿菌RY237乳糖酶具有新穎性和較好的應用潛力,實驗結果為研究和開發乳糖酶提供了理論依據。
[1]Husain Q.β-Galactosidases and their potential applications:a review[J]. Critical Reviews in Biotechnology,2010,30(1):41-62.
[2]張敏文,顧取良,張博,等.乳糖酶研究進展[J].微生物學雜志,2011,31(3):81-86.
[3]Mlichova Z,Rosenberg M. Current trends ofβ-galactosidase application in food technology[J]. Journal of Food and Nutrition Research,2006,45(2):47-54.
[4]Park AR,Oh DK. Galacto-oligosaccharide production using microbial beta-galactosidase:current state and perspectives[J]. Appl Microbiol Biotechnol,2010,85(5):1279-1286.
[5]何熹,王彥寧,李超,等. 乳酸菌與乳糖酶[J]. 食品研究與開發,2016,36(16):202-205.
[6]Hsu CA,Yu RC,Chou CC. Production ofβ-galactosidase byBifidobacteriaas influenced by various culture conditions[J]. International Journal of Food Microbiology,2005,104(2):197-206.
[7]Nagy Z,Kiss T,Szentirmai A,et al. Beta-galactosidase ofPenicilliumchrysogenum:production,purification,and characterization of the enzyme[J]. Protein Expr Purif,2001,21(1):24-29.
[8]Zagustina NA,Tikhomirova AS,Biokhimiia. Purification and properties of beta-galactosidase from fungusCurvulariainaequalis[J]. Biokhimiya,1976,41(6):1061-1066.
[9]Fischer L,Scheckermann C,Wagner F. Purification and characterization of a thermotolerant beta-galactosidase fromThermomyceslanuginosus[J]. Appl Environ Microbiol,1995,61(4):1497-1501.
[10]張紅艷,劉成更.乳糖酶的酶學特性及其研究進展[J].食品研究與開發,2004,25(6):34-36.
[11]鄭兆娟,徐穎,石磊,等.凝結芽孢桿菌中β-半乳糖苷酶基因的克隆及表達[J].南京林業大學學報:自然科學版,2015,39(6):35-39.
[12]宋春雷,閆巧娟,江正強,等.凝結芽孢桿菌產β-半乳糖苷酶條件的優化[J].中國農業大學學報,2010,15(5):110-114.
[13]Batra N,Singh J,Banerjee UC,et al. Production and characterization of a thermostable beta-galactosidase fromBacilluscoagulansRCS3[J]. Biotechnol Appl Biochem,2002,36(1):1-6.
[14]Levin RE,Mahoney RR. Purification and characterization ofβ-galactosidase from a strain ofBacilluscoagulans[J]. Antonie Van Leeuwenhoek,1981,47(1):53-64.
[15]東秀珠,蔡妙英,等編著. 常見細菌系統鑒定手冊[M].北京:科學出版社,2001:62-63.
[16]Osborn F,Blinder R,Justin RE,et al. 顏子穎,王海林譯. 精編分子生物學實驗指南[M].第一版. 北京:科學出版社,2001:39-40.
[17]張衛兵,張炎,文鵬程,等.EnterobacterSYA2乳糖酶酶學性質研究[J].中國釀造,2012,31(11):93-98.
[18]Vivek DF,Susan TLH,Aniruddha BP. Improved cavitational cell disruption following pH pretreatment for the extraction ofβ-galactosidase fromKluveromyceslactis[J]. Biochemical Engineering Journal,2006(31):25-30.
[19]盧麗麗,肖敏,徐曉東.EnterobactercloacaeB5產轉糖基β-半乳糖苷酶發酵條件優化[J]. 應用與環境生物學報,2008,14(1):118-121.
[20]李玉梅,盧麗麗,肖敏. 產轉糖基β-半乳糖苷酶菌株 F3的鑒定、產酶條件及轉糖基活性研究[J].山東大學學報,2009,44(1):1-6.
[21]Karasov P,Spiwok V,Mal KR,et al. Beta-galactosidase activity in psychrotrophic microorganisms and their potential use in food industry[J]. Czech J Food Sci,2002,20:43-47.
[22]高曉峰,周穎,霍貴成.一株植物乳桿菌乳糖酶性質的研究[J].食品工業,2014(11):260-264.
Characterization ofβ-galactosidase fromBacilluscoagulansRY237
LI Wen-ting1,2,BIAN Fei1,WANG Cui-ping1,CHEN Gao1,ZHANG Yan1, ZHU You-feng1,WANG Shi-rong1,YUE Shou-song1,*
(1.Bio-Tech Research Center,Shandong Academy of Agricultural Sciences,Jinan 250100,China; 2.College of Food Engineering and Biotechnology,Tianjin University of Science & Technology,Tianjin 300450,China)
A novel lactase-producing bacterium RY237 was selected based onβ-galactosidase activity using lactose as sole carbon media,and was identified asBacilluscoagulansRY327. The activity of lactase reached 4.98 U/mL. The optimum temperature and optimum pH ofβ-galactosidase were 50 ℃ and 6.0,respectively. The enzyme was stable at pH5.5~8.0,with peak activity at pH7.0,and it was also stable at 40~50 ℃. The activity ofβ-galactosidase was inhibited by Ca2+,K+,Zn2+,Mn2+,Na+,Mg2+and completely inhibited by Cu2+. Its activity was promoted by ethylene diamine tetraacetic acid(EDTA). The high activity,thermostability and pH-stability make this enzyme potentially useful in dairy industry.
Bacilluscoagulans;molecular identification;β-galactosidase;enzymatic characterization
2017-01-03
李文婷(1992-),女,碩士研究生,研究方向:發酵食品與微生物資源開發,E-mail:liwentinghaoye@163.com。
*通訊作者:岳壽松(1965-),男,博士,研究員,研究方向:微生物資源,E-mail:yueshousong@163.com。
山東省農業科學院農業科技創新工程(CXGC2017A01和CXGC2016B13);山東省重點研發計劃項目(2016GGH3111)。
TS201.3
A
1002-0306(2017)14-0111-05
10.13386/j.issn1002-0306.2017.14.022
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
