兩種不同方法制備南極磷蝦富脂蛋白的對比研究
2017-08-09 02:48:32尹利昂薛長湖田利利姜曉明尹利端李兆杰中國海洋大學食品科學與工程學院山東青島66003煙臺新時代健康產業有限公司山東煙臺64006
食品工業科技 2017年14期
尹利昂,薛長湖,田利利,姜曉明,尹利端,李兆杰,薛 勇,*(.中國海洋大學食品科學與工程學院,山東青島 66003; .煙臺新時代健康產業有限公司,山東煙臺 64006)
?
兩種不同方法制備南極磷蝦富脂蛋白的對比研究
尹利昂1,薛長湖1,田利利1,姜曉明1,尹利端2,李兆杰1,薛 勇1,*
(1.中國海洋大學食品科學與工程學院,山東青島 266003; 2.煙臺新時代健康產業有限公司,山東煙臺 264006)
以冷凍南極磷蝦為原料,通過優化自溶條件,采用控制自溶的方式,制備一種富含ω-3的復合物,并就其基本組成、蛋白質變化與分布、營養評價等,與等電點促溶/促沉法制備的富脂蛋白進行對比研究。優化的自溶條件為55 ℃和pH7.5條件下自溶3 h。實驗結果表明,兩種蛋白均為富含ω-3多不飽和脂肪酸的蛋白-脂質復合物,含有全部的人體必需氨基酸,超過FAO/WHO/UNU對食品蛋白質中必需氨基酸的要求,而自溶法制備的富脂蛋白在總脂、磷脂、ω-3多不飽和脂肪酸含量,以及必需氨基酸總量上均優于后者,平均值分別達到了43.88%干基、48.84%總脂、31.52%脂質和517.4 mg/g蛋白。由此可見,控制自溶是一種高效的制備富脂蛋白的方式,有助于磷蝦富脂蛋白作為功能性海洋食品的開發。
南極磷蝦,富脂蛋白,自溶,ω-3多不飽和脂肪酸
南極磷蝦是一種生活在南極海域的浮游生物,在南極食物鏈系統中擔任重要角色[1]。作為地球上生物蘊藏量最大的物種,南極磷蝦資源的開發利用引起全世界越來越多的關注。南極磷蝦,一般指南極大磷蝦(Euphausiasuperba),除了蝦油產品外,其作為人類日常食物的來源尚受到一定的成本和技術上的限制,但它有著均衡的營養組成,整蝦含有77.9%~83.1%的水分、0.4%~3.6%的脂質、11.9%~15.4%的蛋白質,以及約2%的甲殼質和碳水化合物[2]。南極磷蝦蛋白的營養高于一般畜禽肉蛋白和牛乳蛋白,含有全部的人體必需氨基酸,必需氨基酸的總量達到212.1 mg/g 蛋白質[3]。南極磷蝦脂質含有比例可觀的多不飽和脂肪酸,其中,DHA和EPA含量分別為16.6%~36.5%和11.1%~24.8%脂質[4],且多以磷脂型存在[5]。
南極磷蝦加工的限制性因素較多:含有較高比例的可溶性蛋白成分;其蛋白質在本體死亡和冷凍后極易發生變性;南極磷蝦肌肉中含有4.5~570 mg/kg的氟[6];南極磷蝦體內,尤其是頭胸部的消化系統,含有豐富的和較強活性的自溶酶系,主要是蛋白酶類和酯酶類。南極磷蝦的這些特征增加了傳統途徑的加工成本,使磷蝦資源的大宗利用受到限制,而尚未規模化的作為人類食物的來源。
有學者研究發現,等電點促溶/促沉法制備低氟南極磷蝦基料時,所形成的蛋白復合物,在一定程度發生了脂質的富集,可以認為是一種富脂蛋白[7]。本實驗意在利用南極磷蝦自溶酶的生化特點,嘗試制備一種新的富脂蛋白,以推動磷蝦加工的研究和相關產品的進一步開發。迄今,相關學者對磷蝦自溶酶的生化性質進行了較為深入的研究[8-12],但利用自溶酶作用制備南極磷蝦制品的開發尚未見文獻報道。本研究利用冷凍南極磷蝦為原料,經脫氟、優化條件和控制自溶,制備了一種富含ω-3多不飽和脂肪酸(polyunsaturatedfattyacids,PUFAs)的蛋白-脂質復合物,通過與原料蝦肉、等電點促溶/促沉法制備的富脂蛋白相比較,以期證明其是一種營養價值高、安全性好的富脂蛋白。
1 材料與方法
1.1 材料與儀器
南極磷蝦(E.superba) 2015年3月捕撈于南極FAO 48.1海域,中國水產有限公司;三羥甲基氨基甲烷(Tris) 北京索萊寶科技有限公司;對硝基苯酚(p-NP)、對硝基苯酚丁酸酯(p-NPB)、苯甲酰-L-精氨酰-對硝基苯胺(BApNA) 美國Sigma-Aldrich公司;石油醚、乙醚、乙酸、氯仿、甲醇、油酸、乙醇、十二烷基硫酸鈉(SDS) 國藥集團化學試劑有限公司;福林-酚乙液 上海君瑞生物技術有限公司;其他試劑等均為分析純。
JB-1A型磁力攪拌器 上海精密科學儀器有限公司;JJ-1精密增力電動攪拌器、HH-4型數顯恒溫水浴鍋 常州國華電器有限公司;GL-20M型高速冷凍離心機 上海盧湘儀離心機儀器廠;016-240改進型魚肉分離機 中國水產科學研究院上海漁業機械研究所;PF-1氟選擇電極、232甘汞參比電極 上海雷磁精密儀器有限公司;自動電位滴定儀 濟南海能儀器股份有限公司;Biochrom 30+全自動氨基酸分析儀 英國Biochrom公司;6980N氣相色譜儀、5973質譜儀、石英毛細管柱HP-INNOWAX(30 m×0.32 mm×0.25 μm) 美國安捷倫公司;BRUKER AV500核磁共振波譜儀 德國布魯克公司。
1.2 實驗方法
1.2.1 去殼南極磷蝦蝦肉的制備 脫氟:將冷凍南極磷蝦在4 ℃的2 mol/L磷酸溶液中解凍6 h,以降低蝦殼中的氟,防止其向蝦肉中遷移。
采肉:瀝干磷蝦表面水分,以魚肉分離機采取蝦肉,得到去殼南極磷蝦肉。
1.2.2 自溶法制備富脂南極磷蝦蛋白
1.2.2.1 南極磷蝦粗酶液的制備 收集冷凍南極磷蝦蝦頭,加入2倍質量的Tris-HCl緩沖液(0.05 mol/L,pH7.5,4 ℃),高速勻漿后,勻漿液4 ℃、10000 r/min離心30 min,收集上清液,得南極磷蝦粗酶液[13]。粗酶液于-80 ℃冰箱凍存。
1.2.2.2 內源酶最適溫度的測定 取pH7.5的粗酶液,參考Erlanger等的方法[14],以苯甲酰-L精氨酰-對硝基苯胺(BApNA)為底物,以吸光值最高的pH所對應的酶活力為100%,進行粗酶液中蛋白酶在不同溫度條件(5~65 ℃)下相對活力的測定;采用對硝基苯酚(p-NP)法[15],以對硝基苯酚丁酸酯(p-NPB)為底物,以吸光值最高的pH所對應的酶活力為100%,進行粗酶液中脂肪酶在不同溫度條件(5~65 ℃)下相對酶活力的測定。
1.2.2.3 內源酶最適pH的測定 以55 ℃為反應溫度,BApNA為底物,以吸光值最高的pH所對應的酶活力為100%,進行粗酶液中蛋白酶在不同pH(pH3~11)下相對活力的測定;以p-NPB為底物,以吸光值最高的pH所對應的酶活力為100%,進行粗酶液中脂肪酶在不同pH(pH3~11)下相對酶活力的測定。
1.2.2.4 自溶過程及自溶時間選擇 準確稱取8~10 g的去殼南極磷蝦肉,加入3倍體積的Tris-HCl緩沖液(0.05 mol/L,pH7.5,4 ℃),于冰浴條件下高速勻漿。保持pH穩定,加入南極磷蝦肉質量1/20的上述南極磷蝦粗酶液,分別于55 ℃水浴、溫和攪拌條件下自溶1、2、3、4 h,快速升溫滅酶(90 ℃保持5 min),并于冷卻后離心(5000 r/min,10 min),收集沉淀部分即自溶法制備的富脂蛋白。
以Folch方法[16]提取其中的脂質,以薄層色譜法(TLC)分析不同的自溶時間節點時,游離脂肪酸(FFA)、溶血卵磷脂(LPC)等脂質品質劣化代表產物含量的變化趨勢。以油酸(C18∶1)為參照,中性展開劑為石油醚∶乙醚∶乙酸=85∶15∶1(v/v/v),極性展開劑為氯仿∶甲醇∶水=65∶25∶4(v/v/v)。
1.2.3 等電點促溶/促沉法制備富脂南極磷蝦蛋白 準確稱取8~10 g的去殼南極磷蝦肉,加入3倍體積的4 ℃的蒸餾水,高速勻漿。以10 mol/L和4 mol/L NaOH調節勻漿液pH至11.5,在冰浴條件下攪拌30 min以溶解南極磷蝦蛋白,5000 r/min離心10 min,收集沉淀部分,重復一次以上的蛋白溶解操作。將先后兩次離心的上清混合,以10 mol/L和4 mol/L HCl調節混合液pH至4.5,冰浴靜置30 min,5000 r/min離心10 min,收集沉淀部分,得等電點促溶/促沉法制備的富脂蛋白[7]。
1.2.4 基本成分分析 水分含量:采用直接干燥法測定,參照GB/T 5009.3-2010。總脂含量:參照Folch[16]的方法,采用氯仿/甲醇(2/1,v/v)提取磷蝦油,以蝦油質量除以樣品質量,即得總脂含量;該磷蝦油用于脂肪酸組成分析和磷脂、游離脂肪酸含量測定。蛋白質含量:采用凱氏定氮法測定,參照國標GB/T 5009.5-2010。灰分含量:參照國標GB/T 5009.4-2010測定。總氟含量:精確稱量2 g樣品,采用堿熔固定-高溫灰化-氟離子選擇電極法測定[17],以0.1 mol/L高氯酸為氟的提取試劑。
1.2.5 蛋白質的雙向二維電泳分析 提取和純化兩種富脂蛋白,采用雙向二維電泳(2-DE)分析幾種蛋白的分布特征[18]。
1.2.5.1 電泳樣品的制備 在研缽中倒入液氮預冷,然后將樣品置于研缽中,再次迅速加入液氮,待液氮揮發完之后再倒一次液氮,至研缽中液氮停止沸騰后迅速研磨,直至組織變成粉末,均勻而沒有明顯顆粒。在研缽中加入800 μL RB(使用前加入1×PMSF),充分溶解粉末后轉移至1.5 mL EP管中。
將EP管中樣品以超聲破碎儀進行超聲破碎,頻次為8次/輪,共超聲8~9輪。每完成一輪的超聲破碎,將樣品立即冰浴1 min,然后進行離心(4 ℃,12000 r/min,20 min),收集上清液。將上清液分裝成3管,每管約250 μL,每管再加入4倍樣本體積丙酮-20 ℃沉淀過夜。沉淀完的蛋白質進行離心(4 ℃,12000 r/min,20 min),倒去上清液后,平放于干凈的紙巾上自然干燥,即可得到處理后的蛋白質團塊,-80 ℃保存備用。
1.2.5.2 二維電泳條件 一向等電聚焦電泳(IEF):pH3~10;二向垂直電泳(SDS-PAGE):15~170 kDa;染色:銀染。以ImageMaster 2D platinum 5.0進行圖像分析。
1.2.6 氨基酸組成分析 參照GB/T5009.124-2003。
1.2.7 脂肪酸組成分析 蝦油脂肪酸甲酯化條件:取磷蝦油10 mg,加入1 mL 10%濃硫酸-甲醇溶液(或濃鹽酸∶甲醇=1∶5),于90 ℃水浴甲酯化30 min,冷卻后加入正己烷1 mL振蕩,靜置分層,取上清液供GC/MS分析[19]。
色譜條件:HP-INNOWax石英毛細管柱(30 m×0.32 mm×0.25 μm),高純氦氣為載氣,采用恒壓模式,壓力為54 kPa,分流比為25∶1。進樣口溫度為230 ℃,檢測器溫度為250 ℃,柱溫以3 ℃/min由140 ℃升到210 ℃,然后在210 ℃下保持10 min,整個分析過程為33 min。
質譜條件:GC/MS接口溫度280 ℃,EI離子源,電離能量70 eV,離子源溫度230 ℃,掃描周期2.84次/s,質量掃描范圍m/z 50~500 u。
1.2.8 磷脂和游離脂肪酸含量測定 磷脂含量測定[20]:核磁共振法(31PNMR)。以三氯甲烷∶氘代甲醇(2∶1)溶劑為溶劑,磷酸三苯酯(TPP)為內標物進行核磁共振分析。
游離脂肪酸含量測定:采用自動電位滴定法[21],以石油醚∶乙醇(2∶1)為混合溶劑,0.1 mol/L NaOH溶液為滴定試劑。
1.3 數據處理
各數據均平行測定三次。運用分析軟件SPSS 18.0進行統計分析,采用one-way ANOVA算法進行方差分析。多重比較分析采用Tukey’s test算法判斷顯著性差異(p<0.05)。表2~表3同。
2 結果與分析
2.1 自溶條件的優化
2.1.1 自溶溫度和pH的選擇 南極磷蝦自溶酶系主要由蛋白酶和脂肪酶構成,為探索最佳的自溶效果,本實驗驗證了在不同溫度和pH條件下兩種酶類的相對活性。如圖1所示,南極磷蝦蛋白酶系的最高相對活性對應的溫度為55 ℃、pH為7.5,體現了其類胰蛋白酶的屬性。這與以往研究者報道的結果一致,Kubota等[22]認為對自溶起主要作用的是消化道內的蛋白酶,這些酶最適pH為6~8;Osnes等[9]也認為磷蝦自溶酶中起著決定作用是類胰蛋白酶,整體自溶酶活性在中性至弱堿性條件下最強;Kimoto等[10]研究顯示南極大磷蝦的類胰蛋白酶對其自溶起主要的作用;Konagaya[23]通過分離鑒定南極磷蝦體內的蛋白酶,也認為其主要的蛋白酶為類胰蛋白酶。
就脂肪酶類而言,由圖1可以看出,南極磷蝦脂肪酶系為低溫酶,其較適宜的溫度為5~40 ℃,最高相對活性對應的溫度為35 ℃、pH為7.5。Turkiewicz等[24]的研究表明,磷蝦酯酶在pH4.6和37 ℃下具有最大活性,55 ℃下5 min熱處理可以使其完全失活。尚憲明[12]進行了反應溫度和反應pH對酶促反應速率影響的實驗,結論認為,南極磷蝦脂肪酶最佳反應pH為8.0左右,其對較高溫度的穩定性差,在35 ℃具有最佳反應速率,具有低溫脂肪酶的催化特性,并分析這可能與其生活環境相關。
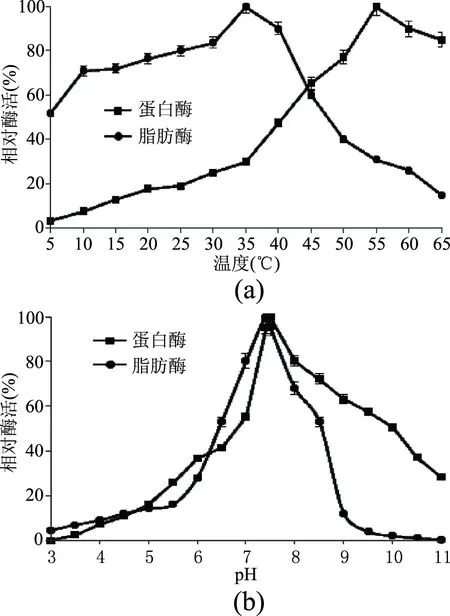
圖1 冷凍南極磷蝦自溶酶的相對活性Fig.1 Relative activity of autoenzyme from frozen Antarctic krill注:(a)和(b)分別表示自溶酶在不同溫度和pH條件下相對活性的變化。
脂肪酶類在自溶過程中,容易導致脂質的酶促降解,引起游離脂肪酸(FFA)和溶血卵磷脂(LPC)水平的上升,使自溶產物的脂質發生品質的劣化,因此有必要采取合適的自溶條件,在促進蛋白酶類作用的同時,抑制脂肪酶類的活性。鑒于蛋白酶在55 ℃和pH7.5分別顯現最高的相對活性,而在55 ℃反應溫度下,脂肪酶類的相對活性只有最高時的20%~30%,本實驗采用的自溶溫度和pH分別為55 ℃和7.5。
2.1.2 自溶時間的選擇 一般來說,隨著自溶時間的延長,脂肪酶作用對自溶產物的不利影響會逐漸增大,如圖2所示,在不同時間(1、2、3、4 h)的平行自溶產物中,隨著自溶時間的延長,作為脂質劣化標志性產物的FFA、LPC呈現增加的趨勢,在1~3 h的自溶中,二者含量的增加比較緩慢,而自溶4 h時則呈現急劇增加的特點。為避免脂質劣化給自溶產物帶來不利影響,優化的自溶時間以3 h為宜。因此,結合自溶效果,本實驗選擇自溶時間為3 h的反應產物為富脂蛋白。

表1 南極磷蝦肉和兩種富脂蛋白的基本成分Table 1 Basic composition of AKM,OEP-A and OEP-ISP
注:AKM,南極磷蝦肉;OEP-A,自溶3 h制備的富脂蛋白;OEP-ISP,等電點促溶/促沉法制備的富脂蛋白。表2~表3同
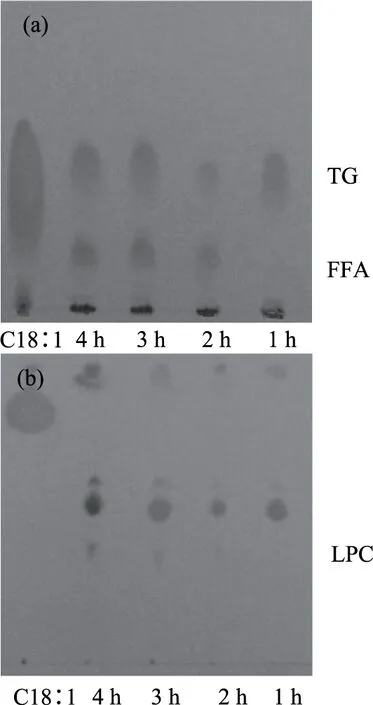
圖2 不同自溶時間所對應脂質的薄層色譜分析Fig.2 TLC analysis oflipid from OEP with different autolysis time注:(a)中性展開;(b)極性展開;TG-甘油三酯。
2.2 基本成分分析
所制備OEP-A的干物質得率平均為40.53%,OEP-ISP的干物質得率平均為41.16%。去殼南極磷蝦肉及兩種富脂蛋白的基本組成見表1。原料中蛋白質含量(68.05%±0.45%干基)遠高于總脂含量(19.72%±0.32%干基)。自溶過程中,蛋白酶能夠有效的降解蛋白質,而在溶解/等電沉淀法中,在等電點pH4.5處,半數以上的蛋白能夠析出而形成沉淀[25],這也是自溶方法的干物質得率略低于后者的原因。
由表1可知,OEP-ISP中的蛋白、總脂的比例接近2∶1,這與王靈昭等[25]使用等電點溶解-沉淀法(ISP方法)所制備的低氟南極磷蝦基料中蛋白、總脂的比例相接近。OEP-A中的蛋白、總脂的比例略高于1∶1,就總脂含量而言,自溶方式比等電點促溶/促沉方式對脂質的富集能力更強,這可能是由于在酶解過程中,蛋白酶作用較強而脂肪酶作用較弱,有相當比例的非水溶性蛋白質經酶解后,以多肽、氨基酸的形式進入到上清液中。兩種富脂蛋白中的平均干基氟含量分別為35.27 mg/kg和24.81 mg/kg,符合美國FDA對食品中氟含量100 mg/kg的限定標準要求,ISP法對氟的脫除效果較自溶方式稍好。這可能是由于ISP過程中,除了經歷水洗作用,蛋白質的完全溶解使氟釋放的比較充分,相關研究結果證實酸處理更能加速氟的脫除[6,26-28]。表1數據對比說明,自溶是有效的富集脂質的方式。同時,自溶過程所產生的上清液,含有蛋白質、多肽、氨基酸及部分脂質,經有效的脫氟后,可以作為后續開發風味類產品的原料。
2.3 蛋白質的分布情況
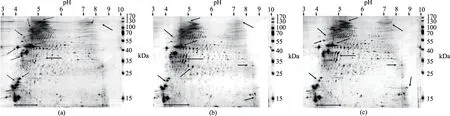
圖3 南極磷蝦蛋白與兩種富脂蛋白的雙向二維電泳圖譜Fig.3 2-DE spectrum of protein from AKM,OEP-A and OEP-ISP注:(a)、(b)、(c)分別為AKM、OEP-A和OEP-ISP蛋白的雙向二維電泳圖。
由于制備原理的不同,OEP-A及OEP-ISP的蛋白分布情況有所差異,如圖3所示。由圖3可以看出,南極磷蝦蛋白的2-DE圖譜上的斑點集中在pH4~5,尤其以pH4.5處最為集中,說明南極磷蝦蛋白的等電點在pH4.5附近,這與已報道的南極磷蝦蛋白等電點pH4.6相近[25],同時說明實驗中制備OEP-ISP所采用的等電點是正確的。南極磷蝦蛋白的分子量集中在35~55 kDa和70~100 kDa。由圖3(a)~圖3(c)對比可以看出,南極磷蝦肉蛋白(圖3(a))在170 kDa處有少量斑點(spots),該斑點通常被認為是魚類、對蝦和磷蝦肌原纖維蛋白中的肌球蛋白重鏈(myosin heavy chain,MHC)的所在[29-30]。MHC斑點或條帶的弱化,是其所在的蛋白質凝膠性和ATPase活性減弱的表征。這說明,南極磷蝦在經歷了死亡和長期凍藏后,其蛋白質可能發生了較嚴重的降解和變性。在圖3(b)及圖3(c)中,MHC的斑點幾乎完全消失,并且代表肌動蛋白的40~55 kDa的斑點也明顯的減少和弱化,這可能是肌原纖維蛋白受溫度、酸堿作用的影響,發生了進一步的降解[31-32]。據文獻報道,在自溶過程中,肌球蛋白與肌動蛋白是蛋白酶作用的主要對象[33]。與此同時,低于35 kDa的小分子量蛋白斑點增加,圖3(c)的增加尤其明顯,說明酸堿對南極磷蝦蛋白質的降解作用較強。
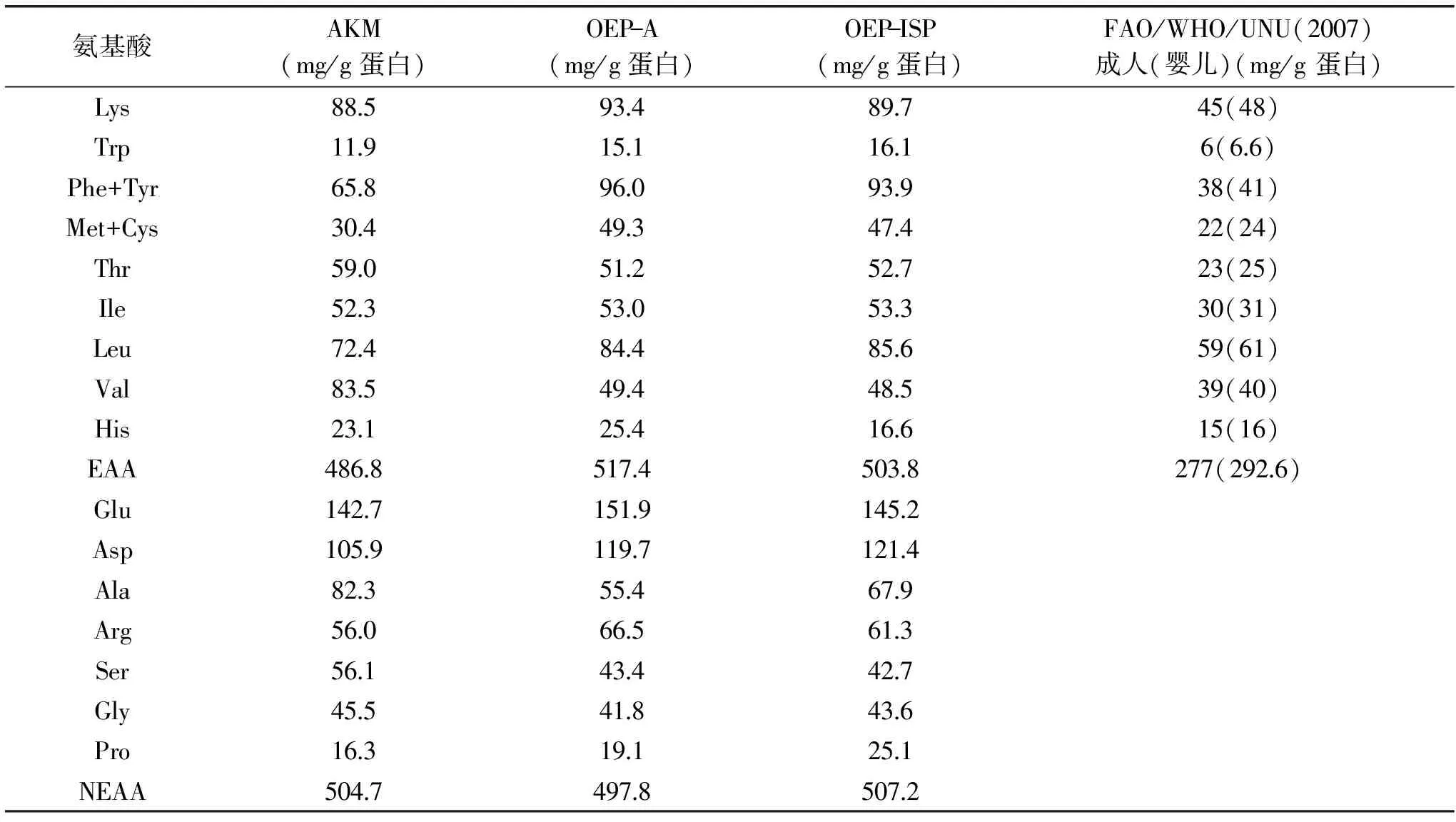
表2 南極磷蝦肉蛋白和兩種富脂蛋白的氨基酸組成Table 2 Amino acid composition of protein from AKM,OEP-A,and OEP-ISP
注:Lys,賴氨酸;Trp,色氨酸;Phe,苯丙氨酸;Tyr,絡氨酸;Met,甲硫氨酸;Cys,半胱氨酸;Thr,蘇氨酸;Ile,異亮氨酸;Leu,亮氨酸;Val,纈氨酸;His,組氨酸;Glu,谷氨酸;Asp,天冬氨酸;Ala,丙氨酸;Arg,精氨酸;Ser,絲氨酸;Gly,甘氨酸;Pro,脯氨酸;EAA,必需氨基酸;NEAA,非必需氨基酸。
2.4 氨基酸組成分析
氨基酸組成特別是必需氨基酸(EAA)的含量,是衡量蛋白質營養價值的重要指標。由表2可以看出,3種南極磷蝦蛋白中必需氨基酸的含量,分別達到486.8、517.4和503.8 mg/g蛋白,均超過FAO/WHO/UNU對成人(嬰兒)食品蛋白中必需氨基酸的要求[277(292.6) mg/g蛋白][34]。OEP-A和OEP-ISP蛋白的必需氨基酸總量在AKM蛋白的基礎上得到提升,OEP-A蛋白的必需氨基酸總量更高。經過兩種方式的處理,賴氨酸、谷氨酸、天冬氨酸和精氨酸的含量增加明顯,其中標志蛋白質營養價值的賴氨酸(Lys),在OEP-A中的含量達到93.4 mg/g蛋白,高于在AKM和OEP-ISP中的含量(88.5、89.7 mg/g蛋白)。
2.5 脂肪酸組成分析
3種脂質的脂肪酸組成如表3所示。OEP-A的PUFAs含量超過了其它2種脂質,其中EPA(C20∶5 n-3)和DHA(C22∶6 n-3)的含量之和達到30.44%,明顯高于AKM的28.39%和OEP-ISP的28.50%,ω-3 PUFAs(C18∶3 n-3、C20∶5 n-3、C22∶6 n-3)含量達31.52%,這說明OEP-A的脂質含有豐富的ω-3 PUFAs,考慮到OEP-A中的總脂含量達到43.88%干基,故本實驗將自溶法制備的脂質-蛋白復合物稱為ω-3富脂蛋白(omega-3-enriched protein,OEP)。
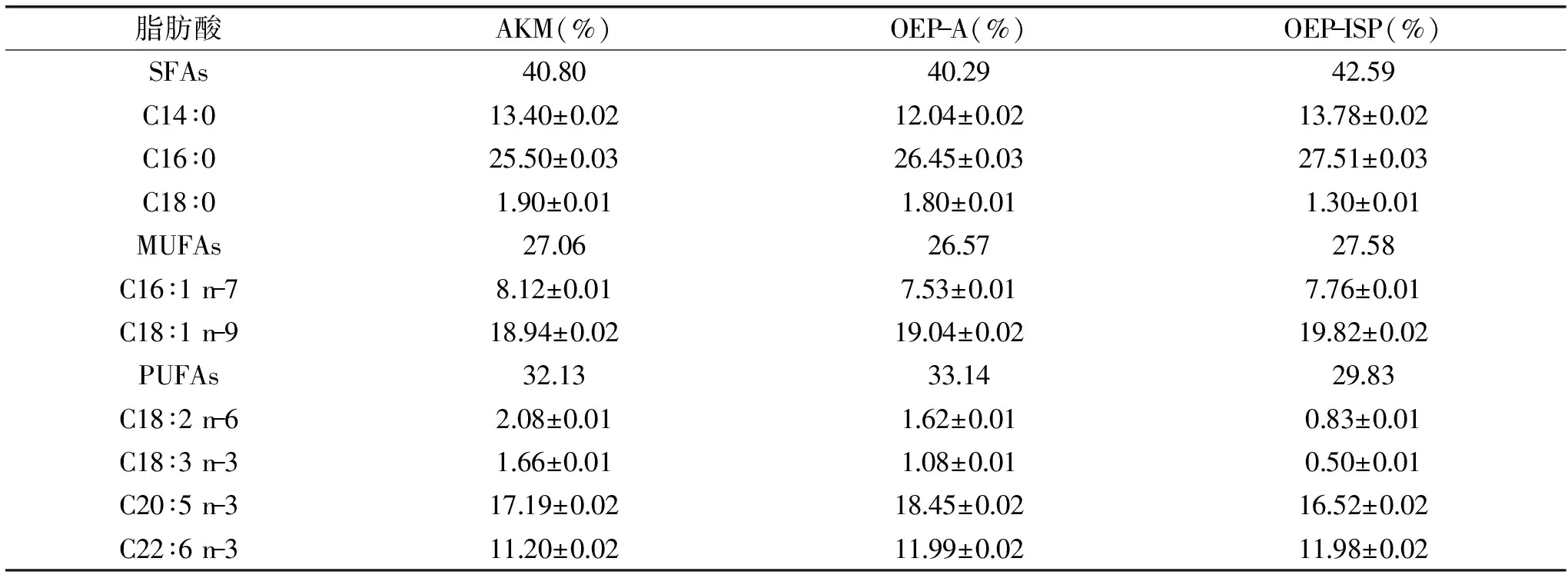
表3 南極磷蝦肉和兩種富脂蛋白脂質的脂肪酸組成Table 3 Fatty acid composition of lipids from AKM,OEP-A,and OEP-ISP
注:SFAs,飽和脂肪酸;MUFAs,單不飽和脂肪酸;PUFAs,多不飽和脂肪酸。
2.6 磷脂和游離脂肪酸含量分析
本實驗采用核磁共振方法(31P NMR)以準確定量脂肪中的磷脂含量,結果見圖4。
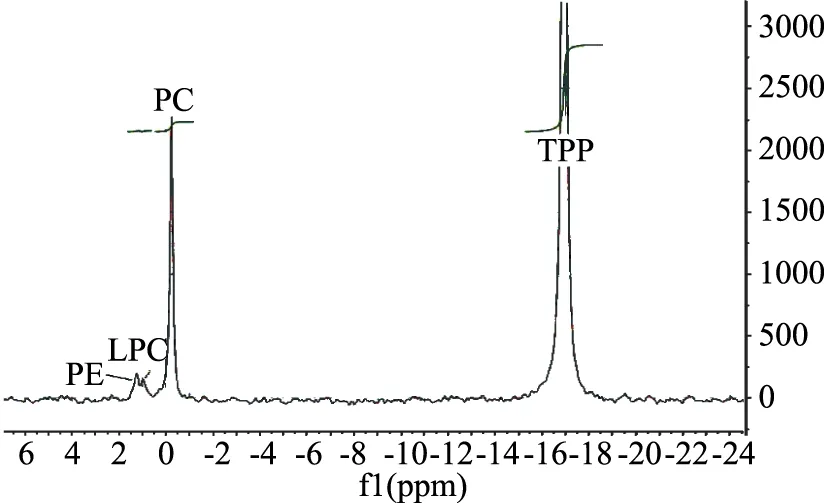
圖4 自溶法所制備的富脂蛋白脂質的31P NMR譜圖Fig.4 31P NMR spectrum of lipid from OEP prepared with autolysis
3種脂質的磷脂和游離脂肪酸含量如表4所示,OEP-A脂質的磷脂含量(48.84%±0.57%總脂)比AKM脂質中磷脂含量(33.16%±0.41%總脂)和OEP-ISP脂質中磷脂含量(34.97%±0.48%總脂)明顯提高,說明OEP-A是一種富含磷脂結合型脂質的復合物。自溶酶解法提高脂質中磷脂含量的作用機制尚未見報道,故這一發現將作為后續研究的重點。OEP-A所含的脂質,可以對人體的神經系統和心血管系統健康起到良好的維護作用[35],不僅是由于其含有豐富的ω-3 PUFAs,還有重要一點是,其脂肪酸主要以磷脂結合的形式存在,據相關研究證實,磷脂型脂質在人體內的吸收效率和安全程度更高[36-37]。因此,OEP-A可以認為是一種富含磷脂型PUFAs的復合物。OEP-A脂質中的游離脂肪酸含量較AKM的略有增加,這可能是自溶過程中脂肪酶的分解作用所致。
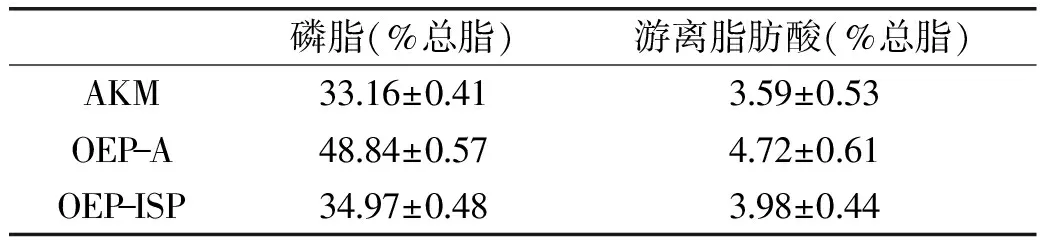
表4 三種脂質的磷脂和游離脂肪酸含量Table 4 Phospholipid and free fatty acid content of lipid from AKM and two OEP
3 結論
本研究的結果表明,可以通過優化的自溶方法:于55 ℃、pH7.5條件下經自溶酶3 h的作用,制備得到富含ω-3的蛋白-脂質復合物。該復合物具有優異的氨基酸及脂肪酸組成,其EAA、ω-3 PUFAs分別達到517.4 mg/g蛋白和31.52%脂質,并含有較高的磷脂含量(48.84%總脂),安全的氟含量(35.27 mg/kg干重)以及合理的游離脂肪酸含量(4.72%總脂)。該富脂蛋白制備過程中產生的副產物可以用于開發磷蝦風味產品,為南極磷蝦的綜合利用提供新途徑。自溶法制備的富脂蛋白對人體健康有積極的意義,具有開發功能性海洋食品的潛在價值。
[1]SuzukiT,Shibata N. The utilization of Antarctic krill for human food[J].Food Reviews International,1990,6:119-147.
[2]Grantham G J. The Southern Ocean:The Utilization of krill[M]. Rome:FAO,1977:1-61.
[3]Chen Y C,Tou J C,Aczynskia J. Amino acid and mineral composition of protein and other components and their recovery yields from whole Antarctic krill(Euphausiasuperba)using isoelectric solubilization/precipitation[J]. Journal of Food Science,2009,74:H31-H39.
[4]Virtue P,Johannes R E,Nichols P D,et al. Biochemical composition ofNyctiphanesaustralisand its possible use as an aquaculture feed source:lipids,pigments and fluoride content[J]. Marine Biology,1995,122:121-128.
[5]Nie Y C,Zhang B,Zhao X Y,et al. Seasonal Variation in Lipids and Protein Content of Antarctic Krill(Euphausiasuperba)[J]. Progress in Fishery Sciences,2016,37:1-8.
[6]Jung H R,Kim M A,Seo Y S,et al. Decreasing effect of fluoride content in Antarctic krill(Euphausiasuperba)by chemical treatments[J]. International Journal of Food Science and Technology,2013,48:1252-1259.
[7]Wang L Z,Xue C H,Wang Y M,et al. Extraction of proteins with low fluoride level from Antarctic krill(Euphausiasuperba)and their composition analysis[J]. Journal of Agricultural and Food Chemistry,2011,59:6108-6112.
[8]Sjo?dahl J,Emmer ?,Vincent J,et al. Characterization of proteinases from Antarctic krill(Euphausiasuperba)[J]. Protein Expression and Purification,2002,26:153-161.
[9]Osnes K K,Mohr V. Peptide hydrolases of Antarctic krill,Euphausiasuperba[J]. Comparative Biochemistry and Physiology B:Biochemistry and Molecular Biology,1985,82:599-606.
[10]Kimoto N. Classification of proteases in Antarctic krill[J]. Agric Biol Chem,1983,47(11):2577-2583.
[11]任艷.南極磷蝦蛋白加工利用的初步研究[D]. 青島:中國海洋大學,2009.
[12]尚憲明.南極大磷蝦脂肪酶提取純化及其酶學性質研究[D]. 青島:中國海洋大學,2011.
[13]杭虞杰,李學英,楊憲時,等.南極磷蝦自溶酶性質的初步研究[J].食品科學,2011,32(13):198-200.
[14]Erlanger B F,Kokowsky N,Cohen W. The preparation and properties of two new chromogenic substrates of trypsin[J].Archs Biochem Biophys,1961,95:271-278.
[15]鄭毅,葉海梅,吳朝娟,等.脂肪酶活力測定研究進展[J].工業微生物,2005,35(9):36-40.
[16]Folch J,Lees M,Sloane-Stanley G H. A simple method for the isolation and purification of total lipids from animal tissues[J].J Biol Chem,1957,226:497-509.
[17]Marian Kjellevold Malde,Kjell Bjorvatna,K?re Julshamn. Determination of fuoride in food by the use ofalkali fusion and fuoride ion-selective electrode[J].Food Chemistry,2001,73:373-379.
[18]O’Farrell PH. High resolution two-dimensional electrophoresis of proteins[J]. J Biol Chem,1975,250:4007-4021.
[19]樓喬明.幾種海洋動物脂質分析及EPA磷脂的活性[D]. 青島:中國海洋大學,2011.
[20]崔常樂,滕英來,汪勇,等. 核磁共振磷譜內標法測定磷脂酰膽堿的含量[J].中國糧油學報,2016,31(5):158-162.
[21]薛斌,曹文明,包杰,等.自動電位滴定法測定米糠油酸值的研究[J].中國油脂,2014,39(11):33-37.
[22]KubotaM,Sakai K. Autolysis of Antarctic krill protein and its inactivation by combined effects of temperature and pH[J]. Trans Tokyo Univ Fish,1978,2:53-63.
[23]Konagaya S.Protease activity and autolysis of Antarctic krill[J].Nippon Suisan Gakkaishi,1980,46(2):175-183.
[24]Turkiewicz M,Kalinowska H,Krystynowicz A,et al. Lipolytic activity of Antarctic krill,EuphausiasuperbaDana[J]. Polish Polar Res,1995,16:185-198.
[25]王靈昭.南極磷蝦(Euphausiasuperba)蛋白質深加工新技術的研究[D]. 青島:中國海洋大學,2012.
[26]Sands M,Nicol S,McMinn A. Fluoride in Antarctic marine crustaceans[J]. Marine Biology,1998,132:591-598.
[27]AdelungD,Buchholz F,Culik B,et al. Fluoride in tissues of krillEuphausiasuperbadana andMeganyctiphanesnorvegicaM. sars in relation to the moult cycle[J]. Polar Biol,1987,7:43-50.
[28]Xie C L,Kim H S,Shim K B,et al. Organic acid extraction of fluoride from Antarctic krillEuphausiasuperba[J]. Fish Aquat Sci,2012,15:203-207.
[29]Yoshikawa K,Inoue N,KawaiY,et al. Changes of the Solubility and ATPase activity of carp myofibrils during frozen storage at different temperatures[J].Fisheries Science,2011,61(5):804-812.
[30]Martinez I,Friis T J,Careche M. Post mortem muscle protein degradation duringice-storage of Arctic(Pandalusborealis)andtropical(PenaeusjaponicusandPenaeusmonodon)shrimps:a comparative electrophoretic and immunological study[J].J Sci Food Agric,2001,81:1199-1208.
[31]Inger V H Kjaersg?rd,Mette R N?rrelykke,Flemming Jessen. Changes in cod muscle proteins during frozen storagerevealed by proteome analysis and multivariate dataanalysis[J].Proteomics,2006,6:1606-1618.
[32]Li T T,Li J R,Hua W Z,et al. Protein changes in post mortem large yellow croaker(Pseudosciaenacrocea)monitored by SDS-PAGE and proteome analysis[J]. Food Control,2014,41:49-55.
[33]Sun L C,Chen Y L,Zhong C,et al. Autolysis of krill protein from North Pacific krillEuphausiapacificaduring protein recovery via isoelectric solubilization/precipitation[J]. Fish Sci,2014,80:839-847.
[34]WHO/FAO/UNU. Protein and amino acid requirements in human nutrition. Report of a joint WHO/FAO/UNU expert consultation[R]. WHO Technical Report Series 935,2007.
[35]Sally T,Rosanna W S,Chung E W,et al. Dietary Krill Oil Supplementation Reduces Hepatic Steatosis,Glycemia,and Hypercholesterolemia in High-Fat-Fed Mice[J].J Agric Food Chem,2009,57:9339-9345.
[36]Ruxton C,Reed S,Simpson M,et al. Health benefits of omega-3 polyunsaturated fatty acids:a review of the evidence[J]. J Hum Nutr Diet,2004,17:449-459.
[37]Gigliotti J C,Davenport M P,Beamer S K,et al. Extraction and characterisation of lipids from Antarctic krill(Euphausiasuperba)[J]. Food Chemistry,2011,125:1028-1036.
Comparative research of two omega-3-enriched protein prepared from Antarctic krill(Euphausiasuperba)with different methods
YIN Li-ang1,XUE Chang-hu1,TIAN Li-li1,JIANG Xiao-ming1,YIN Li-duan2,LI Zhao-jie1,XUE Yong1,*
(1.College of Food Science and Engineering,Ocean University of China,Qingdao 266003,China; 2.Yantai New Era Health Industry Co.,Ltd.,Yantai 264006,China)
Omega-3-enriched protein(OEP)was prepared with controlled autolysis from Antarctic krill(Euphausiasuperba)meat(AKM)with optimization,and compared with OEP prepared with isoelectric solubilization/precipitation(ISP)from basic composition,nutritional evaluation,and variations and distributions of proteins. The optimized condition was that autolysis at 55 ℃ and pH7.5 for 3 h,either protein was demonstrated to be a protein-lipid complex which was rich in ω-3 polyunsaturated fatty acids(ω-3 PUFAs),and exhibited excellent amino-acid composition in all the essential amino acids,exceeding the requirements of the FAO/WHO/UNU for food protein. Total lipid,phospholipid,ω-3 PUFAs and essential amino acids were richer in the OEP prepared with autolysis,and the average values reached 43.88% dry basis,48.84%total lipid,31.52% lipid and 517.4 mg/g protein. This study indicated that controlled autolysis was an efficient way of preparation of ω-3-enriched protein,promotingits development as a functional seafood product in future.
Antarctic krill;ω-3-enriched protein;autolysis;ω-3 PUFAs
2016-12-20
尹利昂(1983-),男,博士研究生,工程師,研究方向:水產品加工,E-mail:yinliang2011@163.com。
*通訊作者:薛勇(1976-),男,博士,教授,研究方向:水產品加工,E-mail:xueyong@ouc.edu.cn。
山東省科技重大專項海洋食品現代加工與產業鏈質量控制體系研究(2015ZDZX05003)。
TS254.4
A
1002-0306(2017)14-0075-07
10.13386/j.issn1002-0306.2017.14.015
