趨化素通過上調(diào)磷酸化JNK促進(jìn)小鼠血管平滑肌細(xì)胞的增殖*
2016-01-31 02:14:16董少紅李江華廖碧紅龐新利羅林杰
中國病理生理雜志 2015年12期
關(guān)鍵詞:細(xì)胞增殖
熊 瑋, 董少紅△, 張 鍵, 李江華, 廖碧紅, 龐新利, 羅林杰
(1暨南大學(xué)第二臨床醫(yī)學(xué)院/深圳市人民醫(yī)院心內(nèi)科,廣東 深圳 518020;2中國科學(xué)院深圳先進(jìn)技術(shù)研究院,廣東 深圳 518055)
?
趨化素通過上調(diào)磷酸化JNK促進(jìn)小鼠血管平滑肌細(xì)胞的增殖*
熊瑋1,董少紅1△,張鍵2,李江華1,廖碧紅1,龐新利1,羅林杰1
(1暨南大學(xué)第二臨床醫(yī)學(xué)院/深圳市人民醫(yī)院心內(nèi)科,廣東 深圳 518020;2中國科學(xué)院深圳先進(jìn)技術(shù)研究院,廣東 深圳 518055)
[摘要]目的: 利用慢病毒構(gòu)建趨化素基因缺陷性小鼠血管平滑肌細(xì)胞株,觀察沉默趨化素基因后血管平滑肌細(xì)胞的增殖情況并探討其機(jī)制。方法: 將正常血管平滑肌細(xì)胞、趨化素基因干擾對(duì)照血管平滑肌細(xì)胞株和趨化素基因缺陷性血管平滑肌細(xì)胞株分成正常組、增殖組、對(duì)照組和沉默組,增殖組加入血小板源性生長因子BB促進(jìn)增殖,采用細(xì)胞計(jì)數(shù)和BrdU法檢測血管平滑肌細(xì)胞增殖,采用Western blot檢測絲裂原激活蛋白激酶( MAPK)信號(hào)通路的蛋白水平。結(jié)果: 增殖組血管平滑肌細(xì)胞的細(xì)胞數(shù)目和BrdU的A值顯著高于正常組(P<0.05);與正常組相比,沉默組的血管平滑肌細(xì)胞數(shù)目和BrdU的A值顯著降低(P<0.05);與此同時(shí),對(duì)照組與正常組之間的血管平滑肌細(xì)胞增殖情況無明顯差異。各組血管平滑肌細(xì)胞之間的p-ERK1/2、ERK1/2、p-p38 MAPK和p38 MAPK水平無明顯變化,增殖組血管平滑肌細(xì)胞的p-JNK蛋白表達(dá)增加,而沉默組血管平滑肌細(xì)胞的p-JNK水平下降。結(jié)論: 趨化素具有促進(jìn)小鼠血管平滑肌細(xì)胞增殖的作用,該作用可能與p-JNK表達(dá)上調(diào)相關(guān)。
[關(guān)鍵詞]血管平滑肌細(xì)胞; 趨化素; 細(xì)胞增殖; c-Jun氨基末端激酶
再狹窄是冠狀動(dòng)脈介入治療之后的主要并發(fā)癥,其發(fā)生是由于介入損傷和血管局部炎癥反應(yīng)導(dǎo)致血管平滑肌細(xì)胞(vascular smooth muscle cells,VSMCs)增殖和遷移到內(nèi)膜下、分泌細(xì)胞外基質(zhì)的結(jié)果[1]。趨化素(chemerin)是一種新發(fā)現(xiàn)的脂肪因子,是孤獨(dú)G蛋白偶聯(lián)受體CMKLR1的內(nèi)源性配體[2]。研究發(fā)現(xiàn),人冠狀動(dòng)脈周圍的脂肪組織及冠狀動(dòng)脈病變部位的VSMCs和泡沫細(xì)胞均可高表達(dá)趨化素,冠心病患者血漿中的趨化素水平亦明顯升高,表明趨化素可能與冠狀動(dòng)脈粥樣硬化的發(fā)生相關(guān)[3- 4]。我們?cè)谇捌趯?shí)驗(yàn)中利用慢病毒構(gòu)建了趨化素基因缺陷性VSMC株,在本研究中我們檢測沉默趨化素基因后VSMCs的增殖情況并探討其機(jī)制。
材料和方法
1試劑和耗材
SPF級(jí)NIH小鼠購自廣東省實(shí)驗(yàn)動(dòng)物中心;DMEM培養(yǎng)基、0.25%胰酶和FBS購自Gibco;小鼠α-SMA抗體購自博士德公司;Lipofectamine 2000購自Invitrogen;BamHⅠ和EcoRⅠ限制性內(nèi)切酶購自Thermo;T4DNA ligase購自NEB;MinElute Reaction Cleanup Kit、Endo-free Plasmid Max Kit購自O(shè)mega;PrimeScript RT reBamHnt Kit和Lenti-X Lentiviral Expression Systems購自TaKaRa;趨化素抗體、p-ERK1/2抗體、磷酸化p38絲裂原激活的蛋白激酶(phosphorylation p38 mitogen- activated protein kinase,p-p38 MAPK)抗體和p-JNK抗體購自Abcam;ERK1/2抗體和p38 MAPK抗體購自Santa Cruz; BrdU抗體購自Biovision。引物由生工公司合成。
2趨化素基因缺陷性VSMC株構(gòu)建和鑒定
按照我們之前建立的方法[5]培養(yǎng)小鼠VSMCs,培養(yǎng)成功后分別將攜帶趨化素shRNA和趨化素shRNAc對(duì)照基因的慢病毒感染VSMCs,用嘌呤霉素對(duì)細(xì)胞進(jìn)行篩選,篩選完成后提取細(xì)胞中RNA和蛋白,分別用real-time PCR和Western blot檢測VSMCs中趨化素的mRNA和蛋白。
實(shí)驗(yàn)分成正常(normal)組、增殖(PDGF)組、對(duì)照(control)組和沉默(knockdown)組,normal和PDGF為正常VSMCs,control為趨化素shRNAc對(duì)照慢病毒VSMC株,knockdown為趨化素基因缺陷性VSMC株。PDGF組細(xì)胞在傳代后加入終濃度為20 μg/L的血小板源性生長因子BB(platelet-derived growth factor-BB,PDGF-BB)刺激VSMCs增殖;normal組、control組和knockdown組VSMCs加入同等體積的PBS。
3實(shí)驗(yàn)方法
3.1VSMCs增殖實(shí)驗(yàn)細(xì)胞消化離心后以每孔1×104個(gè)細(xì)胞接種到96孔板,每組設(shè)6個(gè)復(fù)孔,24 h后進(jìn)行轉(zhuǎn)染,4~6 h后換成100 μL完全培養(yǎng)基。在加入PDGF-BB后48 h時(shí)吸盡培養(yǎng)板中的培養(yǎng)基,用0.25%含EDTA的胰酶30 μL充分消化,加入70 μL培養(yǎng)基,充分吹打以使消化下來的細(xì)胞完全混勻,吸取10 μL細(xì)胞懸液,用計(jì)數(shù)板在顯微鏡下計(jì)算各組VSMCs數(shù)目的變化。
另行BrdU ELISA實(shí)驗(yàn)。在加入PDGF-BB后48 h時(shí)分別在每孔加入BrdU標(biāo)記溶液10 μL,輕柔混合均勻,在37 ℃、5%CO2培養(yǎng)箱中繼續(xù)培養(yǎng)10 h,加入FixDenant溶液200 μL,室溫孵育30 min后,1 500 r/min離心5 min,移除上清液,之后加入anti-BrdU-POD 工作液100 μL,室溫孵育90 min,1 500 r/min離心5 min,移除上清液,用沖洗緩沖液 250 μL反復(fù)沖洗3遍后,移除上清液;加入底物溶液100 μL,室溫孵育20 min;加入1 mol/L H2SO4溶液25 μL,300 r/min離心1 min,在15 min內(nèi)用酶聯(lián)免疫檢測儀測定450 nm波長(參考波長690 nm)的吸光度(A)值。
3.2Western blot實(shí)驗(yàn)細(xì)胞傳代生長48 h后常規(guī)提取總蛋白,每孔加入20 μL蛋白溶液,在10%的SDS-PAGE中80V、30 min及120V、1 h進(jìn)行電泳分離,半干轉(zhuǎn)液38 mA、90 min轉(zhuǎn)入PVDF膜,5%牛奶封閉2 h,加入兔抗小鼠的I 抗(p-ERK1/2稀釋比例為1∶200,ERK1/2為1∶100,p-p38 MAPK為1∶200,p38 MAPK為1∶100,p-JNK為1∶200,GAPDH為1∶1 000),4 ℃孵育過夜,1‰ TBST洗10 min 3次,加入羊抗兔 II 抗(1∶6 000)孵育1 h,1‰ TBST洗10 min 3次,加入ECL發(fā)光液100 μL在ImageQuant RT ECL冷CCD成像系統(tǒng)進(jìn)行顯影,用ImageQuant TL軟件進(jìn)行半定量分析。
4統(tǒng)計(jì)學(xué)處理
所有數(shù)據(jù)以均數(shù)±標(biāo)準(zhǔn)差(mean±SD)表示,使用SPSS 12.0統(tǒng)計(jì)學(xué)軟件分析,各組間比較采用單因素方差分析,各組均數(shù)間的兩兩比較用SNK-q檢驗(yàn),以P<0.05表示差異有統(tǒng)計(jì)學(xué)意義。
結(jié)果
1VMSCs培養(yǎng)及鑒定
用免疫熒光鑒定小鼠血管平滑肌細(xì)胞中α-SMA表達(dá),在胞漿見到大量陽性熒光顆粒,與細(xì)胞核融合,表明血管平滑肌細(xì)胞培養(yǎng)成功,見圖1。
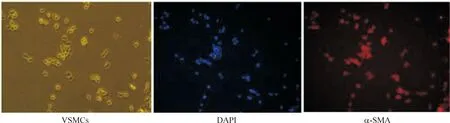
Figure 1.The immunofluorescence staining of mouse VSMCs (×100).
圖1小鼠血管平滑肌細(xì)胞免疫熒光染色
2趨化素基因缺陷性VMSC株構(gòu)建和鑒定
采用real-time PCR檢測3種細(xì)胞株中趨化素的mRNA,Western bolt檢測趨化素蛋白,結(jié)果顯示,空白對(duì)照組的VSMCs (normal)和轉(zhuǎn)染shRNAc的VSMCs (control)之間趨化素mRNA和蛋白無明顯差異,而轉(zhuǎn)染pLVX- Chn1 shRNA的基因缺陷性VMSCs(knockdown)的趨化素mRNA和蛋白水平明顯下降(P<0.01),見圖2,表明趨化素基因缺陷性VSMC株構(gòu)建成功。

Figure 2.The mRNA and protein levels of chemerin in the VSMCs with different treatments were measured by real-time PCR and Western blot. Mean±SD.n=6.**P<0.01vsnormal.
圖2Real-time PCR和Western blot檢測VSMC株的趨化素mRNA和蛋白水平
3趨化素對(duì)血管平滑肌細(xì)胞增殖的影響
分別采用BrdU和細(xì)胞計(jì)數(shù)2種方法檢測沉默趨化素基因后VSMCs的增殖情況。結(jié)果顯示PDGF組的VSMCs數(shù)目和BrdU的A值顯著高于正常VSMCs(P<0.05),表明PDGF-BB促進(jìn)了VSMCs的增殖;與normal組相比,趨化素基因缺陷性VSMC株的細(xì)胞數(shù)目和BrdU的A值顯著降低(P<0.05);與此同時(shí),作為對(duì)照的趨化素shRNAc慢病毒VSMC株與normal組之間的VSMCs增殖情況無明顯差異,見圖3、4。以上結(jié)果表明趨化素具有促進(jìn)VSMCs增殖的作用。

Figure 3.The number of VSMCs was measured by cell counting. Mean±SD.n=6.*P<0.05vsnormal.
圖3細(xì)胞計(jì)數(shù)法檢測VSMCs增殖
4Western blot實(shí)驗(yàn)結(jié)果
細(xì)胞生長48 h后測定各組VSMCs的蛋白,各組細(xì)胞p-ERK1/2、ERK1/2、p-p38 MAPK和p38 MAPK的蛋白水平無明顯差異;與normal組VSMCs相比,PDGF組VSMCs的p-JNK水平增加,而knockdown組VSMCs的p-JNK蛋白水平下降(P<0.05),見圖5。

Figure 4.The BrdU absorbance of the VSMCs with different treatments. Mean±SD.n=6.*P<0.05vsnormal.
圖4BrdU法檢測VSMCs的增殖

Figure 5.The proteins levels of MAPK pathway-related molecules were measured by Western blot.Mean±SD.n=3.*P<0.05vsnormal.
圖5Western blot檢測MAPK通路相關(guān)蛋白的水平
討論
研究表明,趨化素具有調(diào)節(jié)脂肪細(xì)胞分化、促進(jìn)脂質(zhì)代謝、調(diào)節(jié)炎癥和免疫反應(yīng)等生物學(xué)效應(yīng),被認(rèn)為是代謝綜合征的標(biāo)志物。在2型糖尿病患者中發(fā)現(xiàn)其血清趨化素水平明顯升高,在進(jìn)行減肥手術(shù)后趨化素水平明顯下降[6]。關(guān)于趨化素的研究主要集中在代謝綜合征和2型糖尿病。最近,Spiroglou等[3]研究發(fā)現(xiàn),人冠狀動(dòng)脈周圍的脂肪組織及冠狀動(dòng)脈病變部位的VSMCs和泡沫細(xì)胞均可高表達(dá)趨化素,且冠狀動(dòng)脈粥樣硬化病變的嚴(yán)重程度與趨化素的水平呈正相關(guān);在猝死患者的尸體解剖中也發(fā)現(xiàn),冠狀動(dòng)脈不穩(wěn)定斑塊中的泡沫細(xì)胞和內(nèi)皮細(xì)胞均存在趨化素高表達(dá)。Kaur等[7]發(fā)現(xiàn)人血管內(nèi)皮細(xì)胞存在趨化素受體CMKLR1,TNF-α、IL-1β、IL- 6等促炎癥因子可使其表達(dá)上調(diào);趨化素與CMKLR1受體結(jié)合后可通過活化p42/44 MEK信號(hào)通路,誘導(dǎo)血管內(nèi)皮細(xì)胞增殖和新生血管形成。Yoo等[8]檢測了58例肥胖患者的血清趨化素,發(fā)現(xiàn)趨化素與踝肱指數(shù)、體重指數(shù)、胰島素抵抗、低密度脂蛋白、甘油三酯、超敏C反應(yīng)蛋白等呈正相關(guān),與頸動(dòng)脈內(nèi)膜厚度不相關(guān),認(rèn)為循環(huán)趨化素水平升高是動(dòng)脈粥樣硬化的獨(dú)立危險(xiǎn)因素。研究[4]還發(fā)現(xiàn),冠心病患者血漿趨化素顯著升高,與冠狀動(dòng)脈病變支數(shù)、心率、血壓、白細(xì)胞數(shù)量、中性粒細(xì)胞比例、纖維蛋白原、空腹血糖和肌酐水平等密切相關(guān)。最近,Watts等[9]發(fā)現(xiàn)趨化素在血管周圍的脂肪組織中高表達(dá),而CMKLR1則在血管中膜及內(nèi)膜均有表達(dá),用CMKLR1受體的激動(dòng)劑chemerin-9可以呈濃度依賴地促進(jìn)胸主動(dòng)脈收縮,相反CMKLR1受體的拮抗劑CCX832可以抑制去甲腎上腺素和前列腺素 F2α誘導(dǎo)的胸主動(dòng)脈收縮。以上研究均表明趨化素可能與動(dòng)脈粥樣硬化的發(fā)生和進(jìn)展相關(guān),但目前對(duì)于趨化素在動(dòng)脈粥樣硬化中的作用存在爭議,如Lehrke等[10]認(rèn)為血清趨化素水平雖與血糖、血脂水平呈正相關(guān),但不能預(yù)測冠脈病變的發(fā)生。
MAPK是將信號(hào)從細(xì)胞表面轉(zhuǎn)導(dǎo)至細(xì)胞內(nèi)的重要傳遞者,在細(xì)胞增殖、遷移、凋亡、機(jī)體生長發(fā)育以及免疫應(yīng)答等多種細(xì)胞生物過程中發(fā)揮著重要作用[11]。在自發(fā)性高血壓大鼠中,MAPK的活化可促進(jìn)VSMCs的增殖[12],氧化性低密度脂蛋白促進(jìn)VSMCs分泌趨化因子也與MAPK活化有關(guān)[13],而且阻斷MAPK信號(hào)通路可以抑制動(dòng)脈損傷后的內(nèi)膜增生和血管平滑肌細(xì)胞增殖[14]。在本研究中,我們采用RNA干擾的方法,利用慢病毒載體構(gòu)建了趨化素基因缺陷性血管平滑肌細(xì)胞系,用real-time PCR檢測細(xì)胞系中趨化素的mRNA水平,與正常組和對(duì)照組VSMCs相比,該細(xì)胞系的趨化素mRNA水平顯著降低,表明趨化素基因缺陷性VSMC系構(gòu)建成功;采用細(xì)胞計(jì)數(shù)和BrdU法檢測血管平滑肌細(xì)胞增殖,與正常VSMCs相比,趨化素基因缺陷性VSMCs系的細(xì)胞數(shù)目和BrdU的A值均明顯下降,表明趨化素具有促進(jìn)VSMCs增殖的作用。對(duì)MAPK信號(hào)通路的關(guān)鍵分子檢測發(fā)現(xiàn),各組VSMCs的p-ERK1/2、ERK1/2、p-p38 MAPK和p38 MAPK水平無明顯變化,增殖組血管平滑肌細(xì)胞的p-JNK蛋白表達(dá)增加,而沉默組血管平滑肌細(xì)胞的p-JNK水平下降。
綜上所述,我們首次發(fā)現(xiàn)趨化素具有促進(jìn)血管平滑肌細(xì)胞增殖的作用,該作用可能與磷酸化JNK被活化相關(guān)。
[參考文獻(xiàn)]
[1]Curcio A, Torella D, Indolfi C.Mechanisms of smooth muscle cell proliferation and endothelial regeneration after vascular injury and stenting: approach to therapy[J].Circ J, 2011, 75(6):1287-1296.
[2]Ernst MC, Sinal CJ. Chemerin: at the crossroads of inflammation and obesity[J]. Trends Endocrinol Metab, 2010, 21(11):660- 667.
[3]Spiroglou SG, Kostopoulos CG, Varakis JN, et al. Adipokines in periaortic and epicardial adipose tissue: differential expression and relation to atherosclerosis[J]. J Atheroscler Thromb, 2010, 17(2):115-130.
[4]Dong B, Ji W, Zhang Y. Elevated serum chemerin levels are associated with the presence of coronary artery disease in patients with metabolic syndrome[J]. Intern Med,2011, 50(10):1093-1097.
[5]Dong S, Xiong W, Yuan J, et al.MiRNA-146a regulates the maturation and differentiation of vascular smooth muscle cells by targeting NF-κB expression[J].Mol Med Rep, 2013, 8(2):407- 412.
[6]Weigert J, Neumeier M, Wanninger J, et al. Systemic chemerin is related to inflammation rather than obesity in type 2 diabetes[J]. Clin Endocrinol, 2010, 72(3):342-348.
[7]Kaur J, Adya R, Tan BK, et al. Identification of chemerin receptor (ChemR23) in human endothelial cells: chemerin-induced endothelial angiogenesis[J]. Biochem Biophys Res Commun, 2010, 391(4):1762-1768.
[8]Yoo HJ, Choi HY, Yang SJ, et al. Circulating chemerin level is independently correlated with arterial stiffness[J]. J Atheroscler Thromb,2012, 19(1):59- 68.
[9]Watts SW, Dorrance AM, Penfold ME,et al. Chemerin connects fat to arterial contraction[J]. Arterioscler Thromb Vasc Biol, 2013, 33(6):1320-1328.
[10]Lehrke M, Becker A, Greif M, et al. Chemerin is asso-ciated with markers of inflammation and components of the metabolic syndrome but does not predict coronary atherosclerosis[J]. Eur J Endocrinol, 2009, 161(2):339-344.
[11]Burotto M, Chiou VL, Lee JM,et al. The MAPK pathway across different malignancies: a new perspective[J].Cancer, 2014, 120(22):3446-3456.
[12]王旭開,王燕,何作云,等.絲裂原活化蛋白激酶參與胰島素對(duì)自發(fā)性高血壓大鼠血管平滑肌細(xì)胞的增殖研究[J].中國病理生理雜志, 2000, 16(7):596-599.
[13]徐斌,張延斌,許旭光,等.TLR4/MAPKs信號(hào)通路在ox-LDL誘導(dǎo)的血管平滑肌細(xì)胞分泌MCP-1中的作用[J].中國病理生理雜志, 2010, 26(5):848- 852.
[14]Ohashi N,Matsumori A,Furukawa Y,et al.Role of p38 mitogen-activated protein kinase in neointimal hyperplasia after vascular injury[J].Arterioscler Thromb Vasc Biol, 2000, 20(12):2521- 2526.
(責(zé)任編輯: 盧萍, 羅森)

*[基金項(xiàng)目]衛(wèi)生部公益性行業(yè)科研專項(xiàng)(No. 200802173)
Chemerin promotes proliferation of mouse vascular smooth muscle cells by up-regulating p-JNKXIONG Wei1, DONG Shao-hong1, ZHANG Jian2, LI Jiang-hua1, LIAO Bi-hong1, PANG Xin-li1, LUO Lin-jie1
(1DepartmentofCardiology,SecondClinicalMedicalCollegeofJinanUniversity,ShenzhenPeople’sHospital,Shenzhen518020,China;2ShenzhenInstitutesofAdvancedTechnology,ChineseAcademyofSciences,Shenzhen518055,China.E-mail:dsh266@medmail.com.cn)
[ABSTRACT]AIM: To investigate the proliferation property of stable chemerin gene knockdown vascular smooth muscle cells (VSMCs) and to explore its mechanism. METHODS: The normal VSMCs, chemerin gene interfering control VSMCs and stable chemerin gene knockdown VSMCs were divided into normal group, PDGF group, control group and knockdown group. The VSMCs in PDGF group were given platelet-derived growth factor-BB (PDGF-BB) to initiate proli-feration. The cell counting and BrdU assay were employed to investigate the proliferation property of VSMCs. The mitogen-activated protein kinase (MAPK) signal pathway was determined by Western blot. RESULTS: The cell number and BrdU A value in PDGF-BB-treated VSMCs significantly increased as compared with the normal VSMCs(P<0.05).Compared with the normal VSMCs, the chemerin knockdown VSMCs showed obviously decreased cell number and BrdU A value (P<0.05).Simultaneously, no significant difference in the proliferation of VSMCs between the normal VSMCs and the control VSMCs was observed. No significant difference of the protein levels of p-ERK1/2, ERK1/2, p-p38 MAPK and p38 MAPK among 4 kinds of VSMCs was found. The protein level of p-JNK in PDGF-BB-treated VSMCs was up-regulated, while it was down-regulated in chemerin knockdown VSMCs compared with the normal VSMCs. CONCLUSION: Chemerin promotes the proliferation of mouse vascular smooth muscle cells by up-regulating p-JNK production.
[KEY WORDS]Vascular smooth muscle cells; Chemerin; Cell proliferation; c-Jun N-terminal kinase
通訊作者△Tel: 0851-28609838; E-mail: zyyutian@126.com
[收稿日期]2015- 07- 01[修回日期] 2015- 09- 25
[文章編號(hào)]1000- 4718(2015)12- 2287- 05
doi:10.3969/j.issn.1000- 4718.2015.12.029
[中圖分類號(hào)]R363
[文獻(xiàn)標(biāo)志碼]A
猜你喜歡
中國當(dāng)代醫(yī)藥(2017年2期)2017-03-18 18:17:33
右江醫(yī)學(xué)(2016年4期)2017-01-05 16:26:48
中國民族民間醫(yī)藥·下半月(2016年6期)2016-11-02 20:27:22
中國醫(yī)藥科學(xué)(2016年9期)2016-07-25 23:04:03
湖南師范大學(xué)學(xué)報(bào)·自然科學(xué)版(2016年3期)2016-06-25 06:18:59
學(xué)周刊·下旬刊(2016年7期)2016-05-20 18:48:49
課程教育研究·學(xué)法教法研究(2016年6期)2016-04-26 10:06:57
新課程·中學(xué)(2015年9期)2015-10-26 19:00:20
