eEF1A2在原發(fā)性肝細胞癌的表達*
2016-01-31 02:13:52邱福南陳敦雁伍嚴安黃肖利吳文冰
中國病理生理雜志 2015年12期
關(guān)鍵詞:細胞增殖
黃 毅, 邱福南, 陳敦雁, 伍嚴安, 李 峰, 黃肖利, 吳文冰
(福建醫(yī)科大學省立臨床醫(yī)學院 1檢驗科, 2肝膽外科, 3病理科,福建 福州 350001)
?
eEF1A2在原發(fā)性肝細胞癌的表達*
黃毅1△,邱福南2,陳敦雁1,伍嚴安1,李峰3,黃肖利1,吳文冰1
(福建醫(yī)科大學省立臨床醫(yī)學院1檢驗科,2肝膽外科,3病理科,福建 福州 350001)
[摘要]目的: 研究真核翻譯延長因子1A2(eEF1A2)在原發(fā)性肝細胞癌(HCC)組織的表達,以及eEF1A2過表達對HCC細胞生物學行為的影響。方法: 應(yīng)用real-time PCR法和免疫組化法分別檢測62例HCC癌組織與配對癌旁組織、20例正常肝臟組織eEF1A2的mRNA和蛋白表達,應(yīng)用real-time PCR法與Western blot法分別檢測幾種HCC細胞株eEF1A2的mRNA與蛋白表達。構(gòu)建GV287-eEF1A2表達慢病毒感染低表達eEF1A2的HCC細胞,應(yīng)用real-time PCR法和Western blot法分別檢測細胞eEF1A2 的mRNA和蛋白表達;應(yīng)用MTT法、DNA倍體法和real-time PCR法分別檢測細胞活力、細胞周期和白蛋白mRNA的表達。結(jié)果: HCC癌組織eEF1A2的mRNA表達水平和蛋白表達陽性率明顯高于配對癌旁組織與正常肝臟組織(P<0.01);eEF1A2的mRNA和蛋白在HCC細胞株SMMC-7721和BEL-7402高表達,在SK-HEP-1低表達。構(gòu)建的GV287-eEF1A2表達慢病毒可感染SK-HEP-1細胞使eEF1A2過表達;與陰性對照組相比,GV287-eEF1A2組SK-HEP-1細胞的活力升高,白蛋白的mRNA表達水平降低,G0/G1期的細胞比例顯著減少,而S期和G2/M期的細胞比例顯著升高。結(jié)論: eEF1A2在HCC癌組織存在異位高表達;eEF1A2可能是HCC的一種潛在癌蛋白,其過表達可增強HCC細胞的增殖能力,降低HCC細胞的分化程度,并促使細胞周期通過G0/G1期,進入S期和G2/M期。
[關(guān)鍵詞]真核翻譯延長因子1A2; 肝細胞癌; 細胞增殖; 細胞周期
原發(fā)性肝細胞癌(hepatocellular carcinoma,HCC)是我國常見的惡性腫瘤,全世界每年發(fā)生50萬例HCC中一半以上在中國,其相關(guān)致癌機制一直是國內(nèi)外研究的難點。長期以來,對腫瘤的研究多集中在癌基因突變、活化、抑癌基因失活及轉(zhuǎn)錄因子異常等方面,而相對忽略與蛋白質(zhì)合成進程相關(guān)蛋白的致癌作用[1]。人類真核翻譯延長因子1A2(eukaryotic elongation factor 1A2,eEF1A2)參與蛋白質(zhì)翻譯過程肽鏈的延長,傳統(tǒng)觀念被認為是一種管家蛋白。相比其異構(gòu)體eEF1A1,eEF1A2表達具有組織特異性,只局限于心臟、腦和骨骼肌。但近年來的研究發(fā)現(xiàn),eEF1A2在多種其它組織來源的腫瘤中存在著異位高表達[2-7],與腫瘤的發(fā)生發(fā)展密切相關(guān)。本研究旨在觀察eEF1A2在HCC組織與細胞株的表達情況,并構(gòu)建有效表達eEF1A2的慢病毒,探討eEF1A2過表達對HCC細胞增殖的影響。
材料和方法
1研究對象
收集2012年10月~2013年12月期間在福建省立醫(yī)院接受手術(shù)切除并經(jīng)術(shù)后病理證實的HCC患者62例,其中男性52例,女性10例,年齡(54.3±12.2)歲(18~73歲)。所有患者術(shù)前均未行放、化治療。分別取每位患者手術(shù)切除的肝癌組織及配對癌旁組織標本,標本切除后部分置4%甲醛固定,備用免疫組化法檢測;部分置RNA保存液浸泡過夜,然后置于-80 ℃低溫冰箱保存,備用real-time PCR法檢測。正常肝臟組織對照來自2012年10月~2013年12月期間在福建省立醫(yī)院接受手術(shù)切除的20例肝臟血管瘤患者,其中男性12例,女性8例,年齡(40.8±10.4)歲(25~66歲)。所有患者均排除肝炎與肝硬化。取患者手術(shù)切除的肝臟血管瘤周邊組織標本,處理方式同肝癌組織及配對癌旁組織標本。
2細胞培養(yǎng)
人HCC細胞株SMMC-7721、BEL-7402和SK-HEP-1購自中科院細胞所,用含10%胎牛血清的DMEM培養(yǎng)液(Gibco),置37 ℃、5% CO2、飽和濕度的培養(yǎng)箱中培養(yǎng),2~3 d換液傳代1次;實驗用細胞為狀態(tài)良好的對數(shù)生長期細胞。
3實驗方法
3.1Real-time PCR法檢測HCC肝臟癌組織標本與細胞株eEF1A2的 mRNA表達用TRIzol試劑(Invitrogen)提取經(jīng)充分研磨的62例HCC肝臟癌組織和配對癌旁組織、20例正常肝臟組織標本以及處于對數(shù)生長期HCC細胞株SMMC-7721、BEL-7402和SK-HEP-1的總RNA,并對其進行DNase 處理后,分別取2 μg反轉(zhuǎn)錄成cDNA,以SYBR GreenⅠ(ABI)為熒光染料,在real-time PCR儀(ABI 7300)上分別行eEF1A2和磷酸甘油醛脫氫酶(glyceraldehyde phosphate dehydrogenase,GAPDH)基因的擴增。eEF1A2的上游引物為5’-GTCAAGGAAGTCAGCGCCTAC-3’,下游引物為5’-TGAACCACGGCATGTTGGG-3’,擴增產(chǎn)物為124 bp;GAPDH的上游引物為5’-TGACTTCAACAGCGACACCCA-3’,下游引物為5’-CACCCTGTTGCTGTAGCCAAA-3’,擴增產(chǎn)物為121 bp。Real-time PCR反應(yīng)條件為95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,共40個循環(huán)。PCR反應(yīng)結(jié)束后,擴增產(chǎn)物均經(jīng)熔解曲線分析驗證特異性,熔解曲線分析條件為95 ℃ 1 min,20 ℃/s;55 ℃ 1 min,20 ℃/s;55~95 ℃,0.5 ℃/s。以GAPDH為內(nèi)參照,采用 2-ΔΔCt法比較各組eEF1A2 的mRNA相對表達量。
3.2免疫組化法檢測HCC肝臟癌組織標本eEF1A2 蛋白的表達62例HCC肝臟癌組織和配對癌旁組織、20例正常肝臟組織標本經(jīng)4%甲醛固定后,制作成石蠟切片,采用免疫組化SP法染色(試劑盒購于福州邁新生物技術(shù)有限公司),用1∶50稀釋的兔抗人eEF1A2多克隆抗體(Novusbio)進行檢測,嚴格按照說明書操作,陽性判斷標準:細胞漿中出現(xiàn)棕黃色顆粒為陽性細胞,高倍鏡下計數(shù)500個細胞,染色細胞≥10%為陽性,<10%為陰性。
3.3Western blot法檢測HCC細胞株eEF1A2蛋白的表達按1×105∶1的比例加入細胞裂解液提取處于對數(shù)生長期HCC細胞株SK-HEP-1的總蛋白,經(jīng)紫外分光光度計(Beckman DU640)測定含量后,分別取等量總蛋白99 ℃變性5 min,進行12% SDS-聚丙烯酰胺凝膠垂直電泳,電轉(zhuǎn)移至NC膜上;封閉NC膜1 h,加入相應(yīng)抗體4 ℃孵育過夜,其中GAPDH兔抗購自CST,eEF1A2兔抗購自Novusbio;TBS洗滌后,加辣根過氧化物酶標記的羊抗兔IgG(KPL),室溫孵育1 h;TBS洗滌后加LumiGLO Chemiluminescent Substrate(KPL)室溫下孵育1 min,曝光,洗片,然后在GelDoc凝膠圖像分析儀(Bio-Rad)上掃描條帶的灰度,并以GAPDH為內(nèi)參照,計算eEF1A2蛋白的相對表達量。
3.4構(gòu)建eEF1A2表達慢病毒(1)eEF1A2表達慢病毒載體的構(gòu)建:根據(jù)GenBank數(shù)據(jù)庫提供的eEF1A2基因(NM_001958)編碼序列,設(shè)計出含交換配對堿基和酶切位點,并含有目的基因5’端部分序列用于PCR釣取目的基因的一對引物,正向引物為 5’-GAGGATCCCCGGGTACCGGTCGCCACCATGGGC-AAGGAGAAGACCCAC-3’,反向引物為5’-TCCTTGTAGTCCATACCCTTGCCCGCCTTCTGCGCCTT CTGC-GCCGACTTG-3’。經(jīng)PCR擴增后,將得到的PCR產(chǎn)物(1.43 kb)交換入線性化表達載體GV287(AgeI/AgeI酶切,購自上海吉凱基因化學技術(shù)有限公司)中,構(gòu)建重組的GV287-eEF1A2慢病毒表達載體,預(yù)期表達產(chǎn)物為50 kD的eEF1A2蛋白,然后以線性環(huán)化的空載體作為陰性對照,轉(zhuǎn)化感受態(tài)大腸桿菌Dpα,挑取重組陽性菌落行PCR 及測序鑒定。(2)慢病毒包裝:通過Lipofectamine 2000(Invitrogen)將構(gòu)建的重組慢病毒表達載體和相對應(yīng)的包裝質(zhì)粒pHelper 1.0載體、pHelper 2.0載體共轉(zhuǎn)染293T細胞進行慢病毒包裝,48 h后收集上清液,濃縮,測定慢病毒的滴度,分裝,-80oC保存?zhèn)溆谩?3)細胞感染及實驗分組:根據(jù)預(yù)先摸索的MOI值,將獲得的慢病毒感染對數(shù)生長期的SK-HEP-1細胞,12 h后吸去含病毒和聚凝胺(polybrene)的上清液,加入新鮮含10%胎牛血清的DMEM培養(yǎng)液,繼續(xù)培養(yǎng),感染3 d后在熒光顯微鏡下觀察慢病毒上報告基因GFP 的表達情況,感染效率大于80%者繼續(xù)后續(xù)實驗。每次實驗設(shè)3組,分別為:過表達組(over-expression,OE)組,感染GV287-eEF1A2表達慢病毒;陰性對照(negative control,NC)組,感染GV287空載體;空白對照(control,CON)組,未感染慢病毒。(4)GV287-eEF1A2表達慢病毒功能鑒定:慢病毒感染5 d后,收集各組細胞分別提取RNA和總蛋白,采用real-time PCR法檢測eEF1A2的 mRNA,Western blot法檢測eEF1A2的蛋白,觀察GV287-eEF1A2表達慢病毒在SK-HEP-1細胞內(nèi)表達eEF1A2的效果。
3.5MTT法繪制細胞活力曲線收集感染效率大于80% 的SK-HEP-1細胞并接種于96孔板,每孔100 μL,每孔接種細胞數(shù)為2×103,實驗設(shè)OE組、NC組和CON組,每組3個復(fù)孔,連續(xù)培養(yǎng)5 d;實驗結(jié)束前4 h加入5 g/L的MTT(Sigma)10 μL,37 ℃孵育4 h后小心吸去上清,加入100 μL二甲基亞砜(DMSO),置酶標儀(Stat Fax-2100)上振蕩溶解結(jié)晶后用490 nm波長比色,以吸光度A值為縱軸,作用時間為橫軸繪制SK-HEP-1細胞活力變化曲線。
3.6白蛋白(albumin)mRNA表達的檢測慢病毒感染5 d后,分別收集OE組、NC組和CON組SK-HEP-1細胞,采用real-time PCR法檢測白蛋白的mRNA。白蛋白的上游引物為5’-TGCAACTCTTCGTGAAACCTATG-3’,下游引物為5’-ACATCAACCTCTGGTCTCACC-3’,擴增產(chǎn)物為135 bp。Real-time PCR的反應(yīng)條件及擴增產(chǎn)物相對表達量的計算方法見3.1。
3.7 細胞周期的檢測慢病毒感染5 d后,分別收集OE組、NC組和CON組SK-HEP-1細胞,PBS洗滌;加入4 ℃預(yù)冷的70% 乙醇固定細胞至少1 h,1 500 r/min 離心5 min去除固定液,PBS洗滌細胞沉淀1次;按DNA倍體檢測試劑盒(Sigma)說明加入碘化丙碇溶液,避光作用15 min,F(xiàn)ACSCalibur流式細胞分析儀(BD)上檢測細胞周期情況,每次至少計數(shù)10 000個細胞。
4統(tǒng)計學處理
所有實驗均重復(fù)3次,對符合正態(tài)分布的定量數(shù)據(jù),多組間均值的比較采用單因素方差分析(one-way ANOVA)檢驗,兩組間均值的比較采用SNK-q檢驗;對不符合正態(tài)分布的定量數(shù)據(jù),組間的比較采用Nonparametric Tests的Mann-WhitneyU檢驗。陽性率的比較采用χ2檢驗。所有統(tǒng)計學處理均在SPSS 13.0統(tǒng)計軟件包上進行,以P<0.05為差異有統(tǒng)計學意義。
結(jié)果
1HCC肝臟癌組織eEF1A2的表達
以real-time PCR法檢測62例HCC肝臟癌組織與配對癌旁組織、20例正常肝臟組織的eEF1A2的mRNA表達,結(jié)果顯示癌組織eEF1A2的mRNA表達量的中位數(shù)為35.27,明顯高于配對癌旁組織和正常肝臟組織(P<0.01),見圖1。免疫組化染色結(jié)果顯示eEF1A2的陽性表達主要位于胞質(zhì)(圖2)。62例HCC癌組織有47例呈eEF1A2陽性表達,陽性率達75.8%;62例配對癌旁組織僅5例呈eEF1A2弱陽性表達,陽性率為8.1%,20例正常肝臟組織無一例eEF1A2陽性。HCC癌組織的eEF1A2蛋白表達陽性率明顯高于配對癌旁組織與正常肝臟組織(P<0.01)。

Figure 1.The mRNA expression of eEF1A2 in the HCC tissues and cells detected by real-time PCR. The mRNA of GAPDH was used as an internal control. The expression data was plotted as 2-ΔΔCtrelative to normal liver tissues. A: the relative mRNA expression level of eEF1A2 in the HCC tissues; B: the relative mRNA expression level of eEF1A2 in the HCC cells. Mean±SD.**P<0.01vsHCC;##P<0.01vsnormal liver tissues and SK-HEP-1.
圖1HCC肝臟癌組織與細胞eEF1A2 mRNA的表達

Figure 2.The protein expression of eEF1A2 in the HCC tissue and matched liver tissue from an HCC patient, and normal liver tissue analyzed by immunohistochemical staining (SP,×200).
圖2HCC患者癌組織、配對肝臟組織及正常肝臟組織eEF1A2蛋白的表達
2HCC細胞株eEF1A2的表達
Real-time PCR法與Western blot法檢測結(jié)果顯示eEF1A2的mRNA和蛋白在HCC細胞株SMMC-7721和BEL-7402中存在明顯高表達,且以BEL-7402細胞的表達量最高;在HCC細胞株SK-HEP-1中則呈低表達,見圖1、3。
3GV287-eEF1A2表達慢病毒在SK-HEP-1細胞內(nèi)表達eEF1A2的效果
構(gòu)建GV287-eEF1A2表達慢病毒感染SK-HEP-1細胞,使eEF1A2過表達,與NC組相比較,OE組細胞eEF1A2的mRNA和蛋白表達水平分別升高了88.3%和69.2%,兩組之間差異有統(tǒng)計學意義(P<0.01),見圖4。
4eEF1A2過表達對SK-HEP-1細胞活力的影響
MTT檢測結(jié)果顯示,OE組、NC組和CON組SK-HEP-1細胞的細胞活力在培養(yǎng)的第2天開始增加,第3天進入對數(shù)生長期,生長曲線良好;相比NC組,OE組細胞活力增加趨勢更為明顯,在培養(yǎng)第5天的細胞活力明顯升高(P<0.05),見圖5。

Figure 3.The protein expression of eEF1A2 in the HCC cells. GAPDH protein was used as an internal control. Mean±SD.n=3.**P<0.01vsBEL-7402 and SMMC-7721.
圖3HCC細胞eEF1A2蛋白的表達

Figure 4.The effect of GV287-eEF1A2 on the expression of eEF1A2 in SK-HEP-1 cells. GAPDH was used as an internal control. Mean±SD.n=3.**P<0.01vsNC and CON.
圖4GV287-eEF1A2對SK-HEP-1細胞eEF1A2表達的影響

Figure 5.The effect of GV287-eEF1A2 on the activity of SK-HEP-1 cells. Mean±SD.n=3.*P<0.05vsNC and CON.
圖5GV287-eEF1A2對SK-HEP-1細胞活力的影響
5eEF1A2過表達對SK-HEP-1細胞白蛋白mRNA表達的影響
Real-time PCR法檢測結(jié)果顯示,OE組細胞的白蛋白mRNA呈低表達,與NC組相比較其表達水平減低了53.5%,兩組間差異有統(tǒng)計學意義(P<0.01),見圖6。

Figure 6.The effect of GV287-eEF1A2 on the mRNA expression of albumin in the SK-HEP-1 cells. Mean±SD.n=3.**P<0.01vsNC and CON.
圖6GV287-eEF1A2對SK-HEP-1細胞白蛋白mRNA表達的影響
6eEF1A2過表達對SK-HEP-1細胞周期的影響
DNA倍體法檢測結(jié)果顯示慢病毒感染后,OE組處于G0/G1期的SK-HEP-1細胞比例為(38.17±1.49)%,明顯低于NC組的(62.37±2.06)%(P<0.01);而處于S期和G2/M期的細胞比例升高,與NC組相比較差異有統(tǒng)計學意義(P<0.01),見圖7。
討論
eEF1A能與GTP、氨基酰-tRNA形成3聚體,再與核糖體結(jié)合,傳送氨基酸-tRNA入A位,使肽鏈延長,保證蛋白質(zhì)合成的準確性和速度。eEF1A有2個異構(gòu)體,分別為eEF1A1與eEF1A2,兩者序列同源性大于90%,而且功能相同;但在哺乳動物中,兩者的表達模式存在明顯差異:eEF1A1在體內(nèi)組織中廣泛表達,而eEF1A2的表達只限于心臟、腦和骨骼肌。研究表明除了翻譯延長因子的基本功能外,eEF1A2在細胞中還發(fā)揮著其它多種作用,如促進細胞骨架重排、細胞增殖、核內(nèi)轉(zhuǎn)錄活動,抑制衰老,并可通過與一些信號分子如磷脂酰肌醇-4-激酶、磷脂酶Cγ、鈣調(diào)蛋白的相互作用參與磷脂酰肌醇、鈣等信號轉(zhuǎn)導(dǎo)途徑的調(diào)節(jié)[8]。這種多功能蛋白的特性使eEF1A2具有潛在的致癌作用,并在多種腫瘤中存在著異位高表達[2-7]。Anand等[2]最早提出eEF1A2可能在卵巢癌的發(fā)展過程中起重要作用。研究顯示eEF1A2在正常卵巢上皮細胞不表達,但1/3的卵巢腫瘤組織及細胞株中存在eEF1A2 mRNA的異位高表達。同時eEF1A2還具有誘導(dǎo)正常細胞惡性轉(zhuǎn)化的作用,將外源性eEF1A2轉(zhuǎn)染入小鼠成纖維細胞NIpT3,表達eEF1A2的NIpT3細胞可在軟瓊脂糖中呈集落樣生長,且生長速率明顯大于對照組;另外,將高表達eEF1A2的NIpT3細胞植入裸鼠體內(nèi),可引起移植瘤的形成。除卵巢癌外,eEF1A2的異常高表達被發(fā)現(xiàn)也見于乳腺癌、前列腺癌、肺癌、消化道腫瘤中[3-7]。
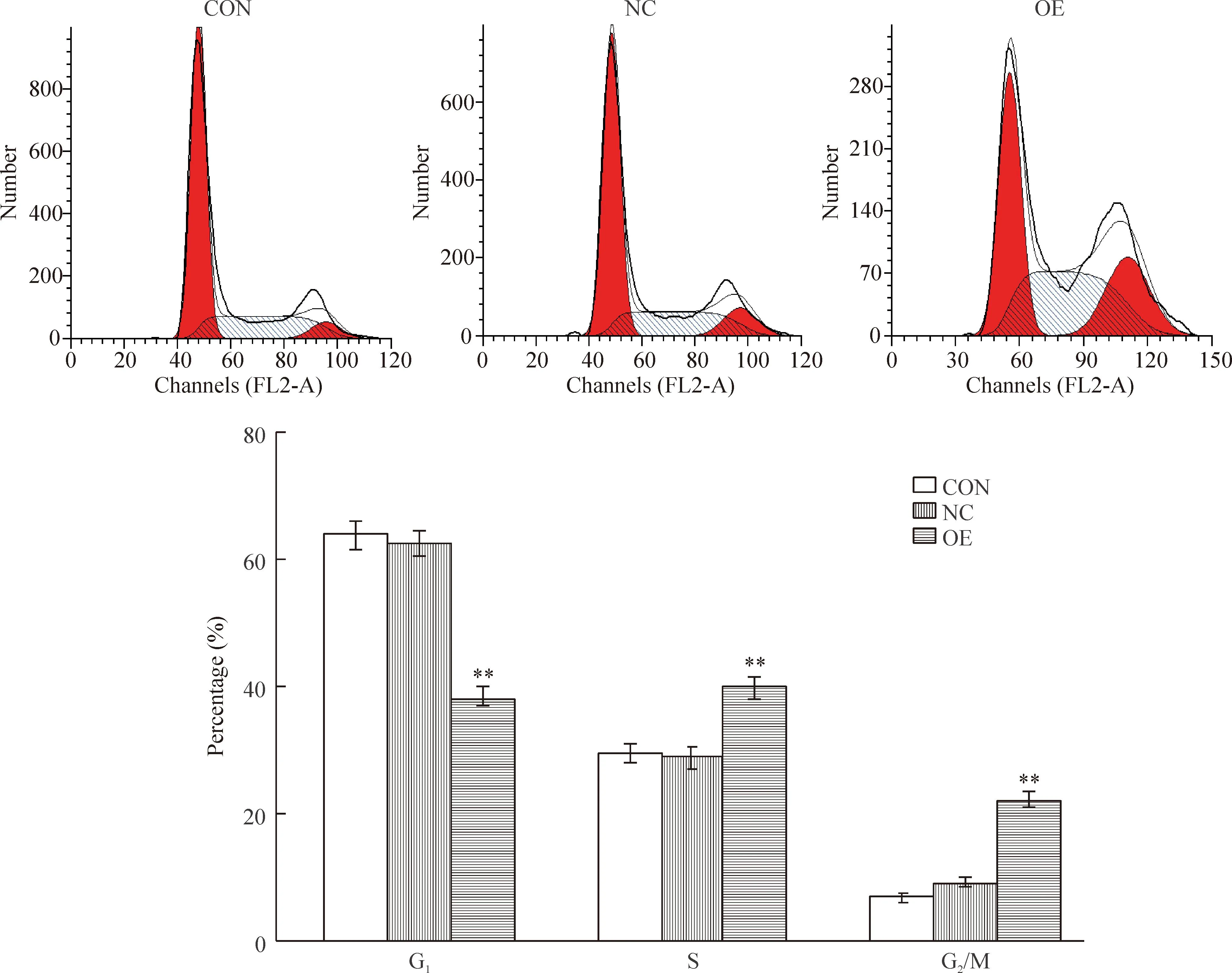
Figure 7.The effect of GV287-eEF1A2 on the cell cycle of SK-HEP-1 cells. Mean±SD.n=3.**P<0.01vsNC and CON.
圖7GV287-eEF1A2對SK-HEP-1細胞周期的影響
我們的研究結(jié)果顯示,eEF1A2的mRNA在HCC癌組織的表達量明顯高于配對癌旁組織與正常肝臟組織;其蛋白的免疫組化陽性率高達75.8%,而配對的癌旁組織僅5例呈弱陽性表達,正常肝臟組織無一例陽性,提示eEF1A2在HCC組織存在著異位高表達。文獻報道eEF1A2能與鋅指蛋白1結(jié)合,再通過與一組具有酪氨酸激酶活性的受體結(jié)合,將信號從胞漿傳到胞核,有效促進細胞的增殖[9]。此外,eEF1A2還可有效結(jié)合蛋白激酶B(Akt),通過提高人乳腺癌BT-549細胞[10]與小鼠漿細胞瘤ABPC4細胞[11]內(nèi)Akt的磷酸化水平,促進這些腫瘤細胞的增殖、侵襲性與遷移能力,增強細胞的惡性度。因此,eEF1A2在HCC的異位高表達是否參與HCC的發(fā)展值得探討。為篩選用于eEF1A2過表達研究的HCC工具細胞,本研究檢測了eEF1A2的mRNA和蛋白在幾種HCC細胞株的表達,發(fā)現(xiàn)相比SMMC-7721和BEL-7402細胞的高表達,SK-HEP-1細胞的eEF1A2表達量低,可作為一種理想的工具細胞。在此工作基礎(chǔ)上,我們以GV287為載體,構(gòu)建了GV287-eEF1A2重組表達慢病毒,通過real-time PCR及Western blot方法檢測顯示GV287-eEF1A2慢病毒在SK-HEP-1細胞內(nèi)可有效表達eEF1A2,表現(xiàn)為與NC組相比較,OE組細胞eEF1A2 mRNA和蛋白的表達水平分別明顯升高了88.3%和69.2%。將該慢病毒感染SK-HEP-1細胞,MTT結(jié)果顯示SK-HEP-1細胞的活力增強,在培養(yǎng)第5天的細胞活力明顯高于NC組,結(jié)合細胞周期分析可認為eEF1A2過表達能促進HCC細胞的增殖。Real-time PCR檢測結(jié)果發(fā)現(xiàn)eEF1A2的過表達還可明顯抑制HCC細胞合成白蛋白的能力,使其mRNA表達水平的減少高達53.5%,而合成白蛋白能力被認為與肝臟細胞分化程度密切相關(guān)[12],提示eEF1A2的過表達可能降低HCC細胞的分化程度,從而增加其惡性表型;對增殖能力增強以及白蛋白合成能力減低的HCC細胞行細胞周期分析,結(jié)果顯示GV287-eEF1A2的感染使G0/G1期的細胞比例顯著減少,而S期、G2/M期的細胞比例顯著升高。以上結(jié)果說明eEF1A2的過表達可增強HCC細胞的增殖能力,降低HCC細胞的分化程度,并促使細胞周期通過G0/G1期、進入S期和G2/M期,提示eEF1A2可能參與了HCC疾病的發(fā)生發(fā)展進程。細胞的增殖、分化及周期調(diào)控異常是腫瘤形成的重要生物學基礎(chǔ),我們認為在HCC異位高表達的eEF1A2可能是HCC的一種潛在癌蛋白,其致癌作用與機制可能是多方面的,除了本身對蛋白質(zhì)翻譯的調(diào)節(jié)外,eEF1A2在HCC細胞是否還參與Akt等信號通路的調(diào)節(jié),以及eEF1A2表達對凋亡、遷移、侵襲等其它腫瘤細胞生物學行為的影響,有待本研究后續(xù)探討。
[參考文獻]
[1]Caraglia M, Budillon A, Vitale G, et al. Modulation of molecular mechanisms involved in protein synthesis machinery as a new tool for the control of cell proliferation[J]. Eur J Biochem, 2000, 267(13):3919-3936.
[2]Anand N, Murthy S, Amann G, et al. Protein elongation factor eEF1A2 is a putative oncogene in ovarian cancer[J]. Nat Genet, 2002, 31(3):301-305.
[3]Kulkarni G, Turbin DA, Amiri A, et al. Expression of protein elongation factor eEF1A2 predicts favorable outcome in breastcancer[J]. Breast Cancer Res Treat, 2007, 102(1): 31-41.
[4]Scaggiante B, Dapas B, Bonin S, et al. Dissecting the expression ofEEF1A1/2 genes in human prostate cancer cells: the potential of EEF1A2 as a hallmark for prostate transformation and progression[J]. Br J Cancer, 2012, 106(1):166-173.
[5]Zhu H, Lam DC, Han KC, et al. High resolution analysis of genomic aberrations by metaphase and array comparative genomic hybridization identifies candidate tumour genes in lung cancer cell lines[J]. Cancer Lett, 2007, 245(1-2):303-314.
[6]Duanmin H, Chao X, Qi Z. eEF1A2 protein expression correlates with lymph node metastasis and decreased survival in pancreatic ductal adenocarcinoma[J]. Hepatogastroenterology,2013, 60(124): 870-875.
[7]Grassi G, Scaggiante B, Farra R, et al. The expression levels of the translational factors eEF1A 1/2 correlate with cell growth but not apoptosis in hepatocellular carcinoma cell lines with different differentiation grade[J]. Biochi-mie, 2007, 89 (12):1544-1552.
[8]Ejiri S. Moonlighting function of polypeptide elongation factor 1: from actin bundling to zinc finger protein R1-associated nuclear localization[J]. Biosci Biotechnol Biochem, 2002, 66(1):1-21.
[9]Panasyuk G, Nemazanyy I, Filonenko V, et al. A2 isoform of mammalian translation factor eEF1A displays increased tyrosine phosphorylation and ability to interact with different signalling molecules[J]. Int J Biochem Cell Biol, 2008, 40(1):63-71.
[10]Amiri A, Noei F, Jeganathan S,et al. eEF1A2 activates Akt and stimulates Akt-dependent actin remodeling, invasion and migration[J]. Oncogene, 2007, 26(27):3027-3040.
[11]Li Z, Qi CF, Shin DM, et al.Eef1a2 promotes cell growth, inhibits apoptosis and activates JAK/STAT and AKT signaling in mouse plasmacytomas[J]. PLoS One, 2010, 5(5): e10755.
[12]Rothschild MA, Oratz M, Schreiber SS. Albumin synthesis[J]. Int Rev Physiol, 1980,21: 249-274.
(責任編輯: 盧萍, 羅森)

*[基金項目] 重慶市衛(wèi)生局重點項目(No. 2013-1-013)
Expression of eEF1A2 in hepatocellular carcinomaHUANG Yi1, QIU Fu-nan2, CHEN Dun-yan1, WU Yan-an1, LI Feng3, HUANG Xiao-li1, WU Wen-bing1
(1DepartmentofClinicalLaboratory,2DepartmentofHepatobiliarySurgery,3DepartmentofPathology,ProvincialClinicalCollege,FujianMedicalUniversity,Fuzhou350001,China.E-mail:hyi8070@126.com)
[ABSTRACT]AIM: To study the expression of eukaryotic elongation factor 1A2 (eEF1A2) in the hepatocellular carcinoma (HCC) tissues and the effects of eEF1A2 over-expression on the biological behaviors of the HCC cells. ME-THODS: The expression of eEF1A2 at mRNA and protein levels in the HCC tissues and matched liver tissues from 62 HCC patients, and 20 normal liver tissues were detected by the methods of real-time PCR and immunohistochemical staining, respectively. The mRNA and protein expression of eEF1A2 in the HCC cells was also determined by real-time PCR and Western blot, respectively. The lentivirus containing eEF1A2 gene was constructed, and was used to infect the HCC cells with low eEF1A2 expression. The expression of eEF1A2 at mRNA and protein levels in the infected cells was detected by real-time PCR and Western blot, respectively. The cell activity, cell cycle and mRNA expression of albumin were measured by MTT assay, DNA ploid analysis and real-time PCR, respectively.RESULTS: The mRNA expression levels and protein expression positive rates of eEF1A2 in the 62 cases of HCC tissues, were significantly higher than those of 62 matched liver tissues and 20 normal liver tissues (P<0.01). eEF1A2 mRNA and protein were highly expressed in SMMC-7721 cells and BEL-7402 cells, and expressed in SK-HEP-1 cells at low level. The expression of eEF1A2 at mRNA and protein levels in the SK-HEP-1 cells was significantly enhanced by infection of GV287-eEF1A2 expression lentivirus.Compared with negative control group (transfected with negative control lentivirus), the cell activity in eEF1A2 over-expression group (transfected with GV287-eEF1A2 expression lentivirus) was significantly enhanced, the mRNA expression of albumin was remarkably reduced, and the cells in G0/G1phase were significantly decreased with increased percentage of the cells in S and G2/M phases.CONCLUSION: eEF1A2 is selectively over-expressed in human HCC cancer tissues. eEF1A2 might be a putative oncoprotein in HCC. eEF1A2 over-expression has noticeable effects on the HCC cell proliferation enhancement, differentiation inhibition, and cell cycle acceleration through the G0/G1phase to S phase and G2/M phases.
[KEY WORDS]Eukaryotic elongation factor 1A2; Hepatocellular carcinoma; Cell proliferation; Cell cycle
通訊作者△Tel: 023-89011536; E-mail: 13220327680@163.com
[收稿日期]2015- 06- 16[修回日期] 2015- 08- 30
[文章編號]1000- 4718(2015)12- 2151- 07
doi:10.3969/j.issn.1000- 4718.2015.12.006
[中圖分類號]R730.23
[文獻標志碼]A
猜你喜歡
中國當代醫(yī)藥(2017年2期)2017-03-18 18:17:33
右江醫(yī)學(2016年4期)2017-01-05 16:26:48
中國民族民間醫(yī)藥·下半月(2016年6期)2016-11-02 20:27:22
中國醫(yī)藥科學(2016年9期)2016-07-25 23:04:03
湖南師范大學學報·自然科學版(2016年3期)2016-06-25 06:18:59
學周刊·下旬刊(2016年7期)2016-05-20 18:48:49
課程教育研究·學法教法研究(2016年6期)2016-04-26 10:06:57
新課程·中學(2015年9期)2015-10-26 19:00:20
