發酵乳中常見微生物應用研究進展
2015-12-16 07:44:06焦晶凱
中國乳品工業 2015年2期
關鍵詞:酵母菌
焦晶凱
(乳業生物技術國家重點實驗室,光明乳業股份有限公司乳業研究院,上海200436)
0 引言
世界上有非常多的發酵乳類型,如酸奶、開菲爾、維利等。它們基于乳酸菌、酵母菌和霉菌菌種的發酵,發酵菌種的多樣化致使發酵乳的風味、質構也各有千秋。使用酵母菌作為發酵劑的發酵乳產品典型代表是開菲爾,開菲爾中由于包含乳酸、乙醛、3-羥基丁酮、少量酒精等物質而呈現特殊的風味。開菲爾是由開菲爾粒發酵而成,開菲爾粒由乳酸菌、酵母菌和醋酸菌等維持一個平衡的微生物體系[1-2]。發酵乳制品種類多種多樣,這些發酵乳主要的區別方式是其中特征微生物的不同,從而導致其終產品特點的不同,除酸奶、開菲爾等常見發酵乳制品外,不同地區也具有一些特色發酵乳,如“Filmjolk”、“Ymer”和 Acidophilin等,文中將進一步闡述這些發酵乳制品的特點及特征微生物。
1 乳酸菌
1.1 酸乳
傳統酸奶主要使用德氏乳桿菌保加利亞亞種(Lactobacillus delbrueckiissp.Bulgaricus)和嗜熱鏈球菌(Streptococcus Thermophilus)作為酸奶的發酵菌種,國標也因此定義酸乳為“以生牛(羊)乳或乳粉為原料,經殺菌、接種嗜熱鏈球菌和保加利亞乳桿菌(德氏乳桿菌保加利亞亞種)發酵制成的產品”。嗜熱鏈球菌和德氏乳桿菌保加利亞亞種對于酸奶的發酵有特殊的功能,不僅單一菌種的特性和添加量對于酸奶的質地和風味有重要影響,兩種菌的添加比例在酸奶的發酵中也有一定的講究。
1.1.1 德氏乳桿菌保加利亞亞種
乳酸桿菌(Lactobacillus)是一類革蘭氏陽性、無孢子、桿狀、過氧化氫陰性、微量需氧型細菌。濃度為5%的二氧化碳可以促進它們的生長。保加利亞乳桿菌(L.Bulgaricus)是三種德氏乳桿菌亞種(Lactobacillus delbrueckiissp.)中的一種[3],另外兩個亞種分別是德氏乳桿菌乳酸亞種(Lactobacillus delbrueckiissp.lactis(Lactobacillus lactis))和德氏乳桿菌德式亞種(Lactobacillus delbrueckii ssp.delbrueckii)。保加利亞乳桿菌(L.Bulgaricus)和乳酸乳桿菌(Lactobacillus lactis)是奶和奶制品中的常見菌種。乳酸桿菌通常被劃分為三個部分:專性同型發酵、兼性異型發酵和專性異型發酵。保加利亞乳桿菌(Lactobacillus Bulgaricus)、嗜酸乳桿菌(Lactobacillus Acidophilus)、乳酸乳桿菌(Lactobacillus lactis)和瑞士乳桿菌(Lactobacillus Helveticus)均屬于專性同型發酵菌株[4]。保加利亞乳桿菌(Lactobacillus Bulgaricus)的最適生長溫度為45℃。目前,已經測序了兩株保加利亞乳桿菌(Lactobacillus Bulgaricus)的完整基因組序列,分別是保加利亞乳桿菌(Lactobacillus Bulgaricus)ATCC 11842和保加利亞乳桿菌(Lactobacillus Bulgaricus)ATCC BAA-365[5]。
1.1.2 嗜熱鏈球菌
嗜熱鏈球菌(Streptococcus Thermophilus)被廣泛應用于乳制品的發酵應用中,因為它具有重要的商業意義,因此嗜熱鏈球菌的物理、生理和代謝特征被廣泛研究。如對碳水化合物發酵過程分子基礎的研究,包括乳糖轉移和代謝的特殊途徑研究。由嗜熱鏈球菌產生的胞外多糖在乳制品發酵過程中具有重要的功能特性,一些胞外多糖已被提取和鑒定。嗜熱鏈球菌抵御細菌噬菌體的抗性機制是目前學者們的研究熱點,特別是CRISPR系統。近來也成功鑒定和測序了一些嗜熱鏈球菌的基因組序列[6]。
1.1.3 菌株間的相互作用
等量的嗜熱鏈球菌(Streptococcus Thermophilus)和保加利亞乳桿菌(L.Bulgaricus)可以得到風味更佳的酸奶產品。酸奶的主要發酵劑在發酵期間具有專性共生關系,并且已經得到了大眾的廣泛認可。混合菌株發酵所產生的酸和風味物質的速率遠遠高于單獨菌株的發酵。酸奶發酵菌株生長需要碳源和氮源,保加利亞乳桿菌(L.Bulgaricus)產生的細胞結合蛋白(特別是prtB)可以產生小肽和氨基酸,主要是纈氨酸,由乳桿菌生成的多肽和氨基酸可以被嗜熱鏈球菌(Streptococcus Thermophilus)的生長所利用[7]。嗜熱鏈球菌(Streptococcus Thermophilus)代謝產生的肽、嘌呤、草酞乙酸、磷酸二鈉、延胡索酸、酪蛋白水解產物、二氧化碳等可以促進保加利亞乳桿菌(Streptococcus Thermophilus)的生長。嗜熱鏈球菌(Streptococcus Thermophilus)的蛋白酶活力遠低于保加利亞乳桿菌(L.Bulgaricus)[8]。但是嗜熱鏈球菌(Streptococcus Thermophilus)的肽酶可以水解保加利亞乳桿菌(L.Bulgaricus)水解酪蛋白的中間產物,這是兩種酸奶發酵劑協同作用的關鍵點。研究發現乳經強熱處理后含有甲酸鈉,甲酸鈉可以促進保加利亞乳桿菌生長。因此熱處理乳可以加速保加利亞乳桿菌的產酸過程,嗜熱鏈球菌在生長過程中代謝能夠產生甲酸,甲酸可以刺激保加利亞乳桿菌的生長。此外嗜熱鏈球菌(Streptococcus Thermophilus)產生嘌呤、嘧啶、二氧化碳、草酰乙酸和反丁烯二酸,這些這些產物均會刺激保加利亞乳桿菌(L.Bulgaricus)的生長。丙酮酸、二氧化碳也能夠刺激保加利亞乳桿菌的生長。有研究認為保加利亞乳桿菌(L.Bulgaricus)的蛋白水解力還受溫度及可溶性氮含量的影響;另外保加利亞乳桿菌(L.Bulgaricus)在牛奶中生長更傾向于利用β-酪蛋白作為氮源,也就是說蛋白的類型也是保加利亞乳桿菌(L.Bulgaricus)生長的主要影響因素。協同作用所帶來的作用是這些菌種更佳快速的生長并有效將乳糖代謝為乳酸,使酸奶發酵在3.5~4 h內完成。此兩種菌發酵產生的酸奶風味與其他發酵乳明顯不同,由混合發酵劑發酵產生的主要風味物質乙醛可達到40 mg/kg。
值得注意的是并不是所有的嗜熱鏈球菌菌株(Streptococcus Thermophilus)和保加利亞乳桿菌(L.Bulgaricus)之間均存在協同共生作用。事實上,它們之間也存在不同程度的拮抗抑制作用。有學者在研究了366對保加利亞乳桿菌(L.Bulgaricus)和嗜熱鏈球菌(Streptococcus Thermophilus)之間的協同生長后發現,僅有8對菌株間存在協同共生作用,即非協同生長的比例為97%。研究的64對菌株組合,僅有12對菌株間存在共生作用,即80%的嗜熱鏈球菌和保加利亞乳桿菌之間沒有表現出協同生長的能力。有研究表明,保加利亞乳桿菌產生的過氧化氫對具有益生作用的乳酸菌有抑制作用;嗜熱鏈球菌(Streptococcus Thermophilus)能夠產生細菌素,同樣保加利亞乳桿菌(L.Bulgaricus)也能夠產生細菌素。有學者研究了嗜熱鏈球菌(Streptococcus Thermophilus)產生的細菌素347在100℃、l h加熱處理后,其抑菌活性仍存在,并使用SDS-PAGE分析驗證一個分子量在2.5~6.2 ku的蛋白類物質能夠影響其殺菌活性。嗜熱鏈球菌(Streptococcus Thermophilus)的抑菌活性也與一個分子量大約為2.5Kda的蛋白質有關。有學者研究保加利亞乳桿菌菌株(L.Bulgaricus)VI1005與德氏乳桿菌菌株(Streptococcus Thermophilus)VI1007之間的相互關系,當在菌株VI1005的培養液中添加菌株VI1007無細胞發酵液后,42℃培養4 h時,菌株VI1005的活菌數較培養開始時下降了1000倍,24 h后的活菌數為0;而VI1005菌株對照組的活菌數培養4 h后增加了40倍。如果用蛋白酶處理菌株VI1007的無細胞發酵液,則可以緩解德氏乳桿菌VI1007無細胞發酵液對保加利亞乳桿菌VI1005的殺菌作用,這意味著菌株VI1007無細胞發酵液的抑菌物質屬于蛋白類成分。多位學者已經證實嗜熱鏈球菌(Streptococcus Thermophilus)和保加利亞乳桿菌(L.Bulgaricus)也能夠代謝生成細菌素,且嗜熱鏈球菌(Streptococcus Thermophilus)對保加利亞乳桿菌(L.Bulgaricus)的抑制作用主要與細菌素物質的產生有關。細菌素屬于熱穩定的、小分子量的非羊毛硫多肽物質。因此在發酵工業中,應特別注意避免兩種菌株之間的相互抑制作用,有助于提高酸奶的產量。
1.2 Ayran發酵乳
1.2.1 Ayran來源
在土耳其,aryan是最流行的發酵乳飲品,它可歸類為稀釋或飲料制品一類,相傳aryan是在一場戰爭中因G?kiirks將酸奶用水稀釋來降低酸度而得以發明,然后aryan由中亞傳到安那托利亞、巴爾干半島和中東地區。但土耳其的aryan有其自身特殊的質地和風味。
1.2.2 Ayran微生物
酸奶發酵劑菌種嗜熱鏈球菌(Streptococcus Thermophilus)和保加利亞乳桿菌(L.Bulgaricus)同樣被用于ayran的發酵,ayran的質地和性狀很大程度取決于這些菌株的活性,因此,選擇合適的發酵菌株至關重要。
在ayran生產中普遍使用酸奶發酵菌種,產粘特性低且發酵速度慢的菌種為ayran生產優選菌株,因為此類菌株可獲得特性穩定的ayran[9],ayran發酵終點pH值也很關鍵,培養溫度和菌株活性會直接影響產品的最終pH值。有學者研究攪拌型發酵乳終點pH值為4.2~4.4相對于pH值為4.7~4.8可以獲得粘性更佳的產品,同樣也有研究證明低pH值(4.3)與高pH值(4.6)相比,低pH值發酵ayran可以獲得更好的質構。如表1所示[10],降低終點pH值會減少乳清析出,增加黏度,但同時也會帶來低pH值對后酸化的影響,導致產品過酸。
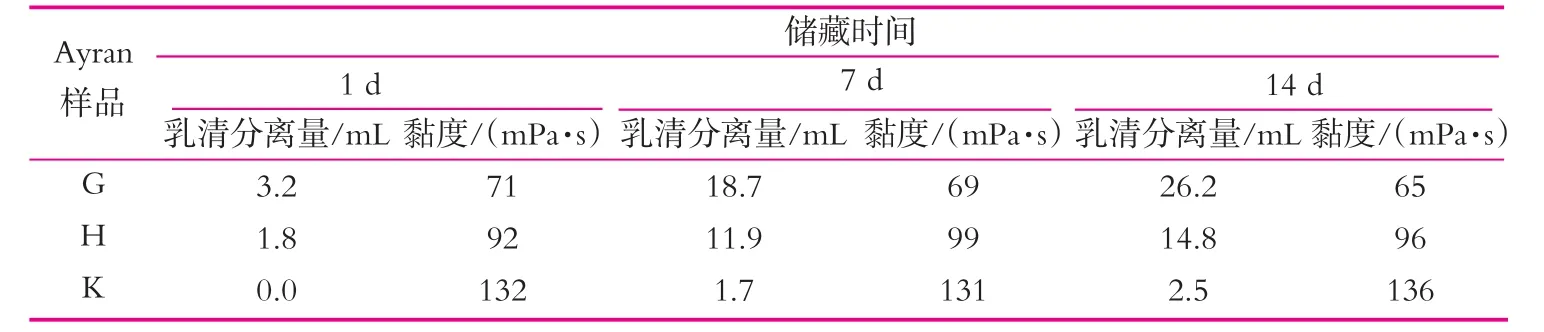
表1 發酵終點pH值對Ayran乳清析出和粘度的影響
Ayran菌種與酸奶微生物類似。工業化條件下,ayran菌種更加穩定,也就是說工業化生產ayran僅包含酸奶菌種。但是家庭手工制ayran易受雜菌污染,酵母特別是馬克思克魯維酵母(Kluyveromyces)和釀酒酵母(Saccharomyces)常見于ayran中。ayran中的菌種要比酸奶中菌種數量大。
在酸奶中,發酵劑可以產生胞外多糖(EPS),從而增加持水性[11-12],這些菌株產黏特性較好,可增加酸奶的穩定性,ayran也一樣,但是需要注意的是并不是所有產EPS菌株都可增加黏度或減少乳清析出,EPS的數量,化學結構以及與牛奶蛋白作用程度均會對黏度產生影響。如表2所示[10],一些產粘好的菌株不一定提高ayran的質構特性。換句話說產EPS高的菌株也不一定能夠提高ayran的特性,即使它們可能會提高其他發酵乳制品的特性,其原因可能為ayran中的蛋白含量減少等原因而使蛋白質之間相互作用增強。
1.3 嗜酸菌牛奶
1.3.1 嗜酸菌牛奶
酸乳桿菌(Lactobacillus Acidophilus)有益于人體健康,通過對其生物組學的研究,使我們更加清楚嗜酸桿菌在人體內的作用機制[13],它對生長條件要求苛刻,在牛奶中生長緩慢[14],因此在生產嗜酸菌牛奶時一定要注意發酵細菌的活性。在發酵過程中牛奶的pH值經常會超過嗜酸乳桿菌的最適生長酸度范圍(pH值為5.5~6.0),從而導致嗜酸菌數量的降低。較高的處理溫度有助于后續嗜酸乳桿菌的生長,因為嗜酸乳桿菌具有相當低的蛋白酶活性,高強度熱處理為嗜酸乳桿菌提供了變性蛋白和多肽等營養成分,對其后續生長極其重要。嗜酸菌接種量一般為2%~5%,發酵完成后嗜酸乳桿菌的數量大約為(2~3)×109mL-1。過長時間發酵會帶來一些不良反應,如到達消費終端時,活菌數量會減少。為了克服長時間發酵帶來的問題,Nahaisi和Robinson[15]使用嗜熱鏈球菌(Streptococcus Thermophilus)和保加利亞乳桿菌(Lactobacillus delbrueckiisubsp.Bulgaricus)混合菌替代25%的嗜酸乳桿菌進行發酵,但值得注意的是,保加利亞乳桿菌產生的過氧化氫對嗜酸乳桿菌有一定抑制作用。

表2 使用產黏和非產粘發酵劑對Ayran產品質構影響
1.3.2 甜嗜酸菌牛奶(Sweet Acidophilus Milk)
盡管嗜酸菌牛奶的益生作用被廣泛認可,但是由于其酸牛奶的風味不被大眾接受而限制了其消費量。因此,為改善嗜酸牛奶的風味使其更加符合大眾的喜好,甜嗜酸菌牛奶應運而生,并流入市場。由于嗜酸乳桿菌在低溫(<10℃)條件下不生長,在5℃下無菌條件下將嗜酸乳桿菌加入到巴氏殺菌奶中,這些菌可以存活14 d而不降低牛奶的pH值。最初加入到牛奶中的嗜酸乳桿菌推薦量為4×109cuf/mL,并在保質期內保持這些菌的數量在規定要求的益生菌作用量之上。也可以使用凍干粉,凍干粉可以在4℃下以58%的存活率存活23 d。
1.4 雙岐乳桿菌乳和嗜酸菌-雙岐乳桿菌乳
工廠加工雙歧乳桿菌乳和A/B乳與嗜酸菌牛奶相似,預先均質達到預定蛋白和脂肪的含量,將牛奶加熱到80~120℃,保持5~30 min,然后迅速冷卻到37℃;向牛奶中加入10%的兩岐雙岐桿菌(Bifidobacterium Bifidum)和長雙歧桿菌(Bifidobacterium Longum)凍干粉,發酵到終點pH值為4.5。發酵結束后,終產品迅速冷卻到10℃以下后包裝,產品有輕微酸味,乳酸和醋酸的比例為2:3,儲藏兩周后雙歧桿菌(bifidobacteria)數量大約在108~109mL。
用于生產A/B乳的牛奶一般標準化到蛋白質高于脂肪含量,標準化牛奶加熱到75℃,保持15 s或85℃保持30 min。在牛奶冷卻到37℃后,加入嗜酸乳桿菌(Lactobacillus Acidophilus)和雙歧桿菌(Bifidobacteria)凍干粉,發酵到pH值為4.5~4.6,大約發酵16 h。隨后將發酵好牛奶冷卻到10℃以下,終產品包含益生菌108~109mL-1,貨架期月20 d。A/B乳有芳香和輕微的酸味,黏度很高。
也可以通過簡單向冷的巴氏殺菌乳中加入嗜酸乳桿菌(Lactobacillus Acidoph-ilus)和兩岐雙岐桿菌(Bifidobacterium Bifidum)混合發酵劑生產益生菌牛奶。該種益生菌牛奶與甜嗜酸牛奶相似。雙岐酸奶(Bifighurt)是另一種益生菌飲品,通過兩岐雙岐桿菌(Bifidobacterium Bifidum)或長雙歧桿菌(Bifidobacterium LongumCKL 1969)和嗜熱鏈球菌(Streptococcus Thermophilus)在42℃發酵而成。接種量為6%,終點pH值為4.7。雙岐酸奶(Bifighurt)具有溫和酸味,含95%的L+乳酸。兩岐雙岐桿菌(Bifidobacterium Bifidum)數量一般高于107mL-1,冷藏1~2周后數量會降低2(對數值)。使用牛奶發酵的雙岐酸奶(Bifighurt)有特殊的風味和口感,被廣泛用于治療胃腸道疾病。
1.5 養樂多
養樂多(Yakult)是甜的具有益生作用的乳飲料,主要加入類干酪乳桿菌類干酪亞種(Lactobacillus Paracaseisubsp.Paracasei Shirota)進行發酵,由脫脂乳、乳粉、白砂糖、葡萄糖和水制成。類干酪乳桿菌類干酪亞種(Lactobacillus Paracaseisubsp.Paracasei Shirota)抗胃液和膽汁,發酵持續16-18 h,終產品含108cfu/mL的類干酪乳桿菌類干酪亞種(Lactobacillus Paracaseisubsp.Paracasei Shirota)。脫脂乳熱處理后會產生美拉德反應,使其變成咖啡色澤,產出比為99%,沒有副產物。也會加入天然香料調香,如西紅柿、芹菜、胡蘿卜、卷心菜、荷蘭芹及其他蔬菜汁。Yakult有時也加入谷物、沙冰、奶昔、芝士蛋糕和其他生冷食物,貨架期一般為冷藏條件下30 d。
Yakult Miru-Miru是Yakult一系列產品,是一款以牛奶為基質的功能性乳飲料,其與牛奶組成非常相似,含質量分數為3.1%的脂肪、3.1%的蛋白質和4.5%的乳糖,也包含一些其他糖類。Yakult Miru-Miru中加入的益生菌為兩岐雙岐桿菌(Bifidobacterium Bifidum)、短雙歧桿菌(Bifidobacterium Breve)和嗜酸乳桿菌(Lactobacillus Acidophilus)。
此外,以單獨以乳酸菌為發酵菌種的發酵乳制品還有 Mil-Mil牛奶、“Filmjolk”和“Lattfil”、“Langfil”和“Ymer”等。Mil-Mil牛奶包含的特色微生物是嗜酸乳桿菌(Lactobacillus Acidophilus)、兩岐雙岐桿菌(Bifidobacterium Bifidum)和短雙歧桿菌(Bifidobacterium Breve),常用葡萄糖和果糖來平衡產品風味,用胡蘿卜汁調色[16];“Filmjolk”和“Lattfil”以乳酸乳球菌乳酸亞種(Lactococcus lactissubsp.lactis)和乳酸乳球菌乳脂亞種(Lactococcus lactissubsp.cremoris)及乳酸乳球菌乳酸亞種丁二酮變種(Lactococcus lactissubsp.lactis biovar diacetyl)和腸膜明串珠菌乳脂亞種(Leuconostoc mesenteroidessubsp.cremoris)發酵而成;“Langfil”中加入了產粘乳酸鏈球菌(Lactic streptococci);“Ymer”使用乳酸乳球菌乳脂亞種(Lactococcus lactissubsp.cremoris)和乳酸乳球菌乳酸亞種雙乙酰變種(Lactococcus lactissubsp.lactis biovar diacetyl)發酵而成。
2 酵母菌
2.1 開菲爾發酵乳
不同種類的乳酸菌(LAB)和酵母菌賦予了開菲爾發酵乳特殊的風味,酵母菌的存在是作為開菲爾的典型代表,而乳酸菌對開菲爾風味的影響也起著至關重要的作用,有研究報告指出愛爾蘭開菲爾粒微生物多樣性由109mL-1的乳球菌(lactococci),108mL-1的明串珠菌(leuconostocs),106mL-1的乳桿菌(lactobacilli),105mL-1的醋酸菌(acetic acid bacteria)和106mL-1的酵母菌組成。開菲爾中最常報道的乳酸菌是同型發酵和異型發酵的乳桿菌(Lactobacillus)、乳球菌(Lactococcus)、明串珠菌(Leuconostoc)和醋酸菌(Acetic acid bacteria)[17],表3所示為開菲爾中常見的乳酸菌,開菲爾中也包含如蠟樣芽胞桿菌(Bacillus cereus)和蘇云金芽孢桿菌(Bacillus thuringiensis)等一些污染菌[21]。
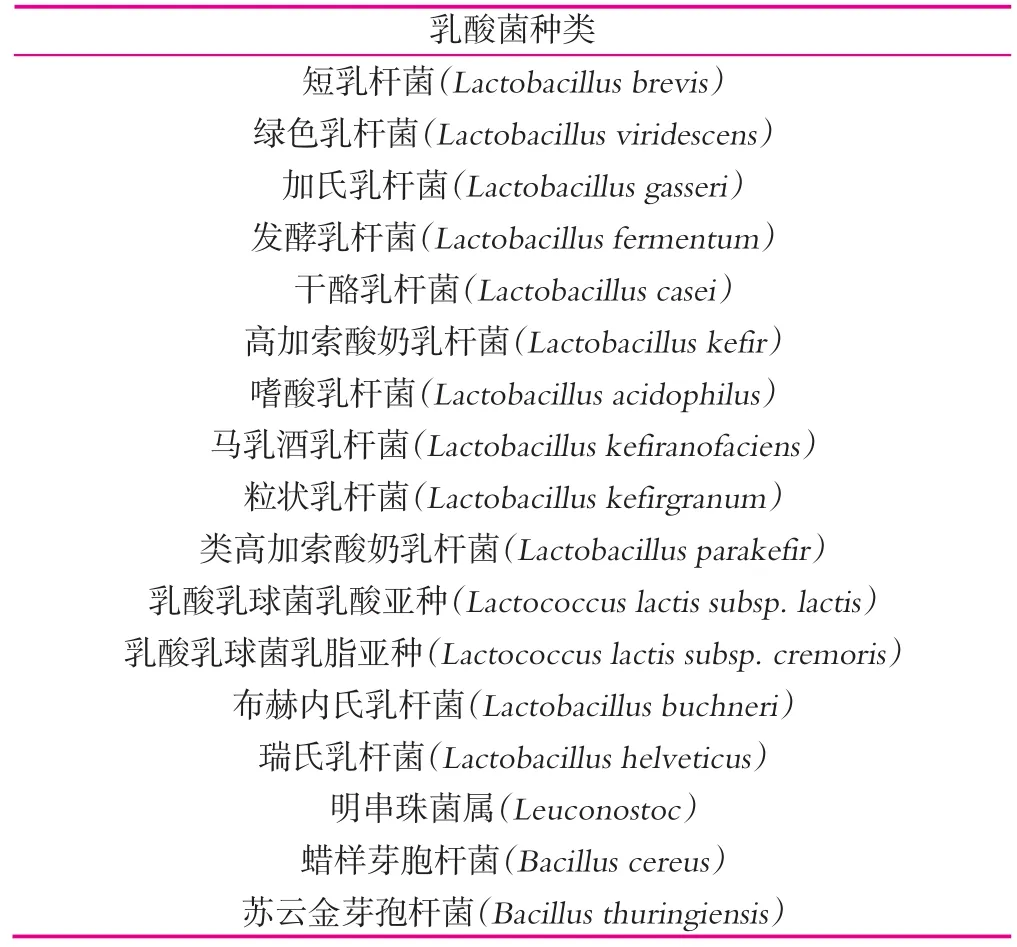
表3 開菲爾中常見的乳酸菌[18-21]
開菲爾中引人注目的還是其中的酵母菌,并且有許多學者分離和鑒定了開菲爾中的酵母菌,常見酵母菌如表4所示。

表4 開菲爾中常見的酵母菌[22-24]
開菲爾之所以有特殊的香氣和質地是由乳酸菌和酵母菌共同作用的結果,Vedamuthu[25]研究發現了開菲爾粒中LAB和酵母有共生關系。另有學者提出酵母菌為LAB提供了一個優化的生長環境,酵母可以代謝部分乳酸并為LAB提供生長刺激物。ChinWen等人[26]鑒定了臺灣開菲爾粒中的乳酸桿菌(LAB)和酵母菌,并對此做了微生物特性研究,他們分離到了瑞士乳桿菌(Lactobacillus helveticus)和腸膜明串珠菌(Leuconostoc mesenterioides)以及馬克思克魯維酵母(Kluyveromyces marxianus)和發酵畢赤酵母(Pichia fermentans)。隨后,他們將這些菌株在30℃的脫脂奶中單獨培養,瑞士乳桿菌(Lactobacillus helveticus)生長速率最快,達到94 min/代,而后是馬克思克魯維酵母(Kluyveromyces marxianus),生長速率是210 min/代,再是生長速率是241 min/代的腸系膜明串珠菌(Leuconostoc mesenterioides),最后是生長速率為300 min/代的發酵畢赤酵母(Pichia fermentans)。馬克思克魯維酵母(Kluyveromyces marxianus)產L-乳酸最多,腸膜明串珠菌(Leuconostoc mesenterioides)產D型乳酸,瑞士乳桿菌(Lactobacillus helveticus)兩種乳酸均生產,馬克思克魯維酵母(Kluyveromyces marxianus)在發酵末期產生很高濃度的酒精。Kroger[27]報告指出歐洲開菲爾發酵乳包含了3×105cfu/mL酵母,108~1010cfu/mL的鏈球菌(streptococci),105cfu/mL的嗜熱乳酸桿菌(thermophilic lactobacilli)和 102~103cfu/mL 的常溫乳酸桿菌(mesophilic lactobacillus)。
一些研究中使用掃描電子顯微鏡(SEM)檢測不同開菲爾粒中的微生物組成,發現開菲爾粒是具有層層網狀基質的海綿針狀結構,并且特別是在中心的位置有分支和相互連接的纖維組織,開菲爾粒內、外部乳桿菌和酵母組成比例不同,開菲爾粒表面由長短不一的乳酸桿菌和酵母菌組成分離式的菌落結構,乳桿菌和酵母菌分別在可繁殖開菲爾粒內部海綿狀基質上形成單獨的菌落,乳桿菌有絲狀附屬物,一般彎曲乳桿菌僅在開菲爾粒內部,而乳球菌則在開菲爾粒表面,Guzel-Seydim等人[28]研究中指出乳桿菌嵌入在土耳其開菲爾粒中,開菲爾粒微生物組成在一個纖維狀基質中,越往中心的位置酵母菌數量越多。
2.2 Koumiss發酵乳
Koumiss是傳統黑發酵乳飲品,源自中亞和俄羅斯地區的游牧部落,并盛行于哈薩克斯坦和吉爾吉斯斯坦。Koumiss是使用馬乳發酵乳制品,發酵劑中包含乳酸菌和酵母菌。有學者研究了三種發酵程度不同的Koumiss,分別是發酵較深的pH值達到3.3-3.6的Koumiss,發酵程度中等pH值達到3.9-4.5的Koumiss和發酵程度較輕的pH值為4.5-5的Koumiss。不同發酵程度的Koumiss中LAB組成不同,分離鑒定的菌株有同型發酵菌株唾液乳桿菌(Lactobacillus Salivarius)、布赫內氏乳桿菌(Lactobacillus Buchneri)和植物乳桿菌(Lactobacillus Plantarum)。Koumiss中酵母菌對其風味、質構等具有重要影響,并且可以和成人體所需的一些營養元素,Mu等人從96個Koumiss樣品中分離出了假絲酵母(Candida pararugosa)、異常德克拉酵母(Dekkera anomala)、地絲菌屬(Geotrichumsp.)、東方伊薩酵母(Issatchenkia orientalis)、單孢釀酒酵母(Kazachstania unispora)、馬克思克魯維酵母(Kluyveromyces marxianus)、畢赤酵母屬(Pichia deserticola、Pichia fermentans、Pichiamanshurica、Pichiamembranaefaciens)、釀酒酵母(Saccharomyces cerevisiae)和德爾布有孢圓酵母(Torulaspora delbrueckii)等[29]。
2.3 其他酵母菌發酵乳
除了開菲爾和Koumiss發酵乳外,使用酵母菌發酵的乳制品還有很多,只是大部分具有局部區域性,沒有得到普及,如嗜酸菌酵母菌牛奶(Acidophilus-yeast milk),在前蘇聯非常常見,是全脂或脫脂牛奶加熱到90~95℃維持10~15 min后,冷卻到35℃,用混合發酵劑嗜酸乳桿菌(Lactobacillus Acidophilus)和乳酸釀酒酵母(Saccharomyces Lactis)發酵,直到酸度達到0.8%乳酸,酒精度達到0.5%。此飲料較為粘稠,有輕微酸味及少量氣泡;此外,Acidophilin也是一款甜酵母菌發酵制品,與嗜酸菌酵母菌牛奶(Acidophilus-yeast milk)類似,用嗜酸乳桿菌(Lactobacillus Acidophilus)、乳酸乳球菌乳酸亞種(Lactococcus Lactissubsp.Lactis)和開菲爾酵母或開菲爾發酵劑發酵而成,并向牛奶中添加乳粉、蔗糖或奶油。Acidophilin已被廣泛應用治療結腸炎、小腸結腸炎、痢疾和其他胃腸道疾病。
3 霉菌
發酵乳制品中,除使用乳酸菌和酵母菌外,在芬蘭生產的維利(Viili)酸奶中使用霉菌,它是一種半固體酸奶,呈均一黏稠膠態并有適口的酸味,形成這一特性主要是發酵乳中乳酸菌及生長在表面的真菌在起作用,Viili中的乳酸菌主要是產莢膜菌株乳酸乳球菌乳酸亞種(Lactococcus lactissubsp.lactis)、乳酸乳球菌乳脂亞種(Lactococcus lactissubsp.cremoris)、乳酸乳球菌丁二酮變種5(Lactococcuslactis subsp.Lactis biovar diacetyl)和腸膜明串珠菌乳脂亞種(Leuconostoc mesenteroidessubsp.cremoris);真菌主要是白地霉(Geotrichum candidum),常被稱作乳霉,它使發酵乳形成絨毛狀表面,使產品略帶一點發霉的味道。此外Viili中還含有馬克思克魯維酵母(Kluveromyces marxianus)和發酵畢赤酵母(Pichia fermantans)。
4 結論
有數據顯示,過去8年,我國酸奶、發酵型乳酸菌飲料市場保持持續快速增長,2006年到2013年酸奶銷售額增長3.5倍,乳酸菌飲料增長9倍,呈現明顯的快速增長趨勢,其中功能酸奶是低溫酸奶主要增長驅動引擎。隨之,發酵乳的市場競爭也非常強烈,而保持發酵乳的競爭優勢的有效手段之一,即為對發酵乳菌種優勝劣汰,本文介紹了發酵乳制品中的常見微生物,包括細菌、酵母菌和真菌,隨著發酵乳行業的不斷壯大,越來越多的發酵乳制品發展起來。此外,近十年,醫療費用的昂貴使消費者不得不轉向了廉價而有效途徑來保持身體的健康,消費者對功能性食品的關注度越來越高,而乳制品特別是功能性、益生菌類乳制品占據功能性食品的巨大市場,本文為今后發酵乳制品的發展提供理論基礎。
[1]GOREK A,TRAMEK M.Quantitative examination of process parameters during kefir grain biomass production[J].International Journal of Chemical Reactor Engineering,2007,5(1):1-8.
[2]SERAFINI F,TURRONI F,RUAS-MADIEKO P,et al.Kefir fermented milk and kefiran promote growth of Bifidobacterium bifidum PRL2010 and modulate its gene expression[J].International Journal of Food Microbiology,2014,178:50-59.
[3]BERGEY D,HOLT J,KRIEG P,et al.Bergey's Manual of Determinative Bacteriology[M].Williams and Wilkins,USA,1994.
[4]STILES M E,HOLZAPFEL W H.Lactic acid bacteria of foods and their current taxonomy[J].Int J Food Microbiol,1997,36(1):1-29.
[5]VAN DE GUCHTE M,PENAUD S,GRIMALDI C,et al.The complete genome sequence of Lactobacillus bulgaricus reveals extensive and ongoing reductive evolution[J].Proc Natl Acad Sci U S A,2006,103(24):9274-9279.
[6]HUTKINS R,GOH Y J.Encyclopedia of Food Microbiology(Second Edition)[M].Oxford:Academic Press,2014.
[7]FIRA D,KOJIC M,BANINA A,et al.Characterization of cell envelope-associated proteinases of thermophilic lactobacilli[J].Journal of Applied Microbiology,2001,90(1):123-130.
[8]SETTACHAIMONGKON S,NOUT MJR,ANTUNES FERNANDES EC,et al.Influence of different proteolytic strains of Streptococcus thermophilus in co-culture with Lactobacillus delbrueckii subsp.bulgaricus on the metabolite profile of set-yoghurt[J].International Journal of Food Microbiology,2014,177(0):29-36.
[9]NILSSON L E,LYCK S,TAMIME A Y.Production of Dringking products[M].Blackwell Publishing Ltd,2007.
[10]YILDIZ F.Development and manufacture of yogurt and other functional dairy products[M].CRC Press,2010.
[11]KRISTO E,MIAO Z,CORREDIG M.The role of exopolysaccharide produced by Lactococcus lactis subsp.cremoris in structure formation and recovery of acid milk gels.Int Dairy J,2011,21(9):656-662.
[12]MENDE S,PETER M,BARTELS K,et al.Addition of purified exopolysaccharide isolates from S.thermophilus to milk and their impact on the rheology of acid gels.Food Hydrocolloid,2013,32(1):178-185.
[13]SELLE K M,KLAENHAMMER T R,RUSSELL W M.Encyclopedia of Food Microbiology(Second Edition)[M].Oxford:Academic Press,2014.
[14]SALJI J.Acidophilus milk products:foods with a third dimension[J].Food Science and Technology Today,1992,6(3):142-147.
[15]NAHAISI M,ROBINSON R.Acidophilus drinks:the potential for developing countries.Dairy Industries International,1985,50:16-17.
[16]KURMANN J A,RASIC J L,KROGER M.Encyclopedia of fermented fresh milk products.As reported by Lourens-Hattingh A and Viljoen B(2001)Yoghurt as a probiotic carrier food.International Dairy Journal,1992,11:1-17.
[17]SIMOVA E,BESHKOVA D,ANGELOV A,et al.Lactic acid bacteria and yeasts in kefir grains and kefir made from them[J].Journal of Industrial Microbiology and Biotechnology,2002,28(1):1-6.
[18]ANGULO L,LOPEZ E,LEMA C.Microflora present in kefir grains of the Galician region(North-West of Spain)[J].J Dairy Res,1993,60(2):263-267.
[19]LEITE A M O,MAYO B,RACHID CTCC,et al.Assessment of the microbial diversity of Brazilian kefir grains by PCR-DGGE and pyrosequencing analysis[J].Food Microbiol,2012,31(2):215-221.
[20]NALBANTOGLU U,CAKAR A,DOGAN H,et al.Metagenomic analysis of the microbial community in kefir grains[J].Food Microbiol,2014,41:42-51.
[21]HOSSEINI H,HIPPE B,DENNER E,et al.Isolation,identification and monitoring of contaminant bacteria in Iranian Kefir type drink by 16S rDNA sequencing[J].Food Control,2012,25(2):784-788.
[22]ROHM H,ELISKASES-LECHNER F,BR?UER M.Diversity of yeasts in selected dairy products[J].J Appl Bacteriol,1992,72(5):370-376.
[23]LEITE A M O,LEITE DCA,DEL AGUILA E M,et al.Microbiological and chemical characteristics of Brazilian kefir during fermentation and storage processes[J].J Dairy Sci,2013,96(7):4149-4159.
[24]FUQUAY J W,FOX P F,MCSWEENEY P L H.Encyclopedia of Dairy Sciences(Second Edition)[M].San Diego:Academic Press,2011.
[25]VEDAMUTHU E.Exotic fermented dairy foods[J].Journal of Food Protection,1977,40:801-802.
[26]CHINWEN L,HSIAOLING C,JERUEI L.Identification and characterisation of lactic acid bacteria and yeasts isolated from kefir grains in Taiwan[J].Aust J Dairy Technol,1999,54(1):14-18.
[27]KROGER M.Kefir[J].Cultured dairy products journal,1993:28-29.
[28]GUZEL-SEYDIM Z,WYFFELS J T,SEYDIM A C,et al.Turkish kefir and kefir grains:microbial enumeration and electron microscobic observation[J].Int J Dairy Technol,2005,58(1):25-29.
[29]MU Z,YANG X,YUAN H.Detection and identification of wild yeast in Koumiss[J].Food Microbiol,2012,31(2):301-308.xndxjjk@126.com
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
中國民間療法(2021年8期)2021-07-22 05:53:28
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
當代水產(2019年3期)2019-05-14 05:43:24
天然產物研究與開發(2018年9期)2018-10-08 03:25:32
煙草科技(2015年8期)2015-12-20 08:27:04
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
食品工業科技(2014年9期)2014-03-11 18:15:31
