誘導耐藥型宋內志賀菌中主動外排阻遏基因acrR、marR的變化
2015-05-06 08:15:21高佩增姚明曉
中國實驗診斷學 2015年10期
關鍵詞:耐藥
高佩增,姚明曉
(1.濟南市第四人民醫院檢驗科,山東濟南250031;2.山東省疾病預防控制中心病原生物研究所)
誘導耐藥型宋內志賀菌中主動外排阻遏基因acrR、marR的變化
高佩增1,姚明曉2
(1.濟南市第四人民醫院檢驗科,山東濟南250031;2.山東省疾病預防控制中心病原生物研究所)
目的分析宋內志賀菌在被環丙沙星誘導耐藥后其主動外排阻遏基因acrR、marR突變與表達量的變化。方法 通過1/2MIC逐級誘導宋內志賀菌對環丙沙星耐藥,通過提取誘導株誘導前后DNA和總RNA,采用基因測序檢測突變和實時熒光定量PCR方法檢測acrR、marR基因表達量的變化。結果 2株誘導耐藥株中acrR、marR中均不存在基因突變,相對于誘導前acrR、marR Ct值增加,基因表達量降低。結論 acrR、marR基因表達量降低間接導致主動外排基因acrAB-tolC轉錄水平提高,證實了誘導劑環丙沙星激活了菌株的主動外排系統。
誘導耐藥;主動外排;阻遏基因;acrR;marR
Key words:Induced resistance;Active efflux;Repressor gene;acrR;marR
(Chin J Lab Diagn,2015,19:1625)
隨著抗生素的應用,志賀菌耐藥現象逐年增加,近年來,宋內志賀菌分離的比率逐年增高,有逐漸取代福氏志賀菌的趨勢。志賀菌耐藥機制涉及多個方面,其中,主動外排系統,尤其是耐藥結節分化家族(resistance nodulation division,RND)成員對革蘭陰性菌獲得性耐藥起重要作用,acrAB-tolC是RND家族的典型代表,由藥物質子轉運子AcrB、膜融合蛋白AcrA和外膜通道蛋白TolC構成,抗菌藥物進入細菌的周質或細胞質后,會被內膜上的AcrB捕獲,然后AcrB與AcrA形成AcrAB復合體,再與TolC結合形成三聯復合體[1,2],AcrAB的表達受多種調節因子的調節,其中阻遏子基因acrR和mar-RAB操縱子的作用最為重要,AcrR抑制子是由位于泵基因上游的阻遏子基因arcR編碼的,對acrAB轉錄起一定程度的抑制作用,MarR是marRAB操縱子的阻遏基因,主要抑制marA,一旦MarA表達增加,同時引起acrA、acrB過量表達,使細菌的耐藥性顯著增加[3,4]。雖然國內外很多單位對AcrAB-TolC進行了研究,研究對象主要是大腸埃希菌,也有少數研究福氏志賀菌和傷寒沙門菌等[5,6],本文通過檢測acrR和marR在菌株被環丙沙星誘導耐藥后基因突變與表達水平的變化,來驗證宋內志賀菌中主動外排泵的表達變化。
1 材料與方法
1.1 菌株來源
采用我院分離的宋內志賀菌敏感菌株(編號:22、38),以及誘導成功后的宋內志賀菌耐藥菌株(22R、38R)。
1.2 主要試劑
TaKaRa Taq kit,Marker D2000,RNAiso Plus,PrimeScriptTMRT reagent Kit with gDNA E-raser(Perfect Real Time),SYBR○RPremix Ex TaqTMII(Tli RNaseH Plus)均購自寶生物工程(大連)有限公司。E-test試條(氨芐西林、頭孢噻肟、慶大霉素、環丙沙星、氯霉素、四環素、復方新諾明)購自梅里埃公司。
1.3 主要儀器
實時熒光定量PCR儀GeneLight 2400(廈門安普利),電泳儀JY300C(北京君意),紫外分析儀JY02S(北京君意),高速低溫冷凍離心機Eppendorf 5430R(艾本德中國有限公司)。
1.4 方法
1.4.1 宋內志賀菌的誘導 取對數生長期宋內志賀菌,用M-H肉湯校正濃度為0.5麥氏單位,使菌液濃度為1×108CFU/mL,稀釋200倍后終濃度為5×105CFU/mL。向菌液中加入濃度為1/2MIC0環丙沙星溶液,充分混勻后塞緊塞子,放入恒溫培養箱中37℃振搖培養至細菌生長的對數期,重復以上步驟,不斷提高誘導劑環丙沙星的濃度,當環丙沙星的濃度達到1mg/L后,濃度以1mg/L遞增,且每一濃度誘導兩代以確保耐藥,直至誘導劑環丙沙星濃度大于或等于128×MIC0。誘導過程中每誘導3代接種一次M-H瓊脂平板,置35℃孵育16-18h,按菌株鑒定步驟進行生化反應和血清型鑒定,確保誘導過程中菌株不被污染。
1.4.2 E-test法測誘導前后菌株的MIC 挑取經35℃培養16-18h后的菌落,用生理鹽水調整至0.5McF。在15min內用無菌棉試子蘸取菌液,管壁上擠去多余液體,涂布于M-H平板,平板旋轉60度涂布3次并沿邊緣涂布一圈,放置數分鐘后小心貼上E-test試條。35℃培養16h-18h后讀取結果。結果根據2013年版CLSI(M100-S19)標準判讀。
1.4.3 宋內志賀菌總DNA的提取 用無菌棉棒挑取適量孵育16-18h誘導株的菌苔溶于50μl雙蒸餾水中,95℃水浴5min,12 000rpm離心30秒,含有DNA的上清液于-20℃冰箱保存備用。
1.4.4 acrR與marR基因擴增與測序 根據Gen-Bank公布的acrR和marR基因序列,采用軟件Primer Premier 5自行設計引物,見表1中acrR-1與marR-1的引物序列,以總DNA為模板進行PCR反應,PCR反應體系為:5U/μl TaqDNA聚合酶0.5μl、10×緩沖液(Mg2+plus)5μl、10mmol/L dNTP 4μl、25mmol/LMgCl22μl、10μmol/L引物5μl、模板DNA5μl、無菌雙蒸水補足總反應體積為50μl,反應條件:95℃預變性5min;94℃變性25s,55℃退火20s,72℃延伸25s,共35個循環;最后72℃延伸5min。反應結束后進行瓊脂糖凝膠電泳,擴增產物送寶生物工程(大連)有限公司進行基因測序。
1.4.5 宋內志賀菌總RNA的提取及轉錄 宋內志賀菌總RNA提取參照RNAiso Plus試劑盒說明,取1μg總RNA按照PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time)進行DNA消除,然后進行逆轉錄。
1.4.6 宋內志賀菌主動外排基因表達水平檢測(RT-PCR) 根據GenBank公布的acrR和marR基因序列,采用軟件Primer Premier 5自行設計引物,PCR引物合成委托寶生物工程(大連)有限公司完成。RT-PCR引物序列見表1中acrR-2,marR-2。熒光定量PCR反應體系與反應條件。25μl反應體系中:SYBR○RPremix Ex Taq(Tli RNaseH Plus)12.5μl,上下游引物10μM各1μl,cDNA 2 μl,dH2O 8.5μl,RT-PCR循環參數;反應條件:95℃預變性5min;94℃變性25s,52℃退火20s,72℃延伸25s,共35個循環;最后72℃延伸5min。分別讀取每個菌株的acrR、marR和16sRNA的Ct值,acrR、marR的Ct值與16sRNA的Ct值之比為其相對含量,比較敏感株與耐藥株中acrR和marR的相對含量。
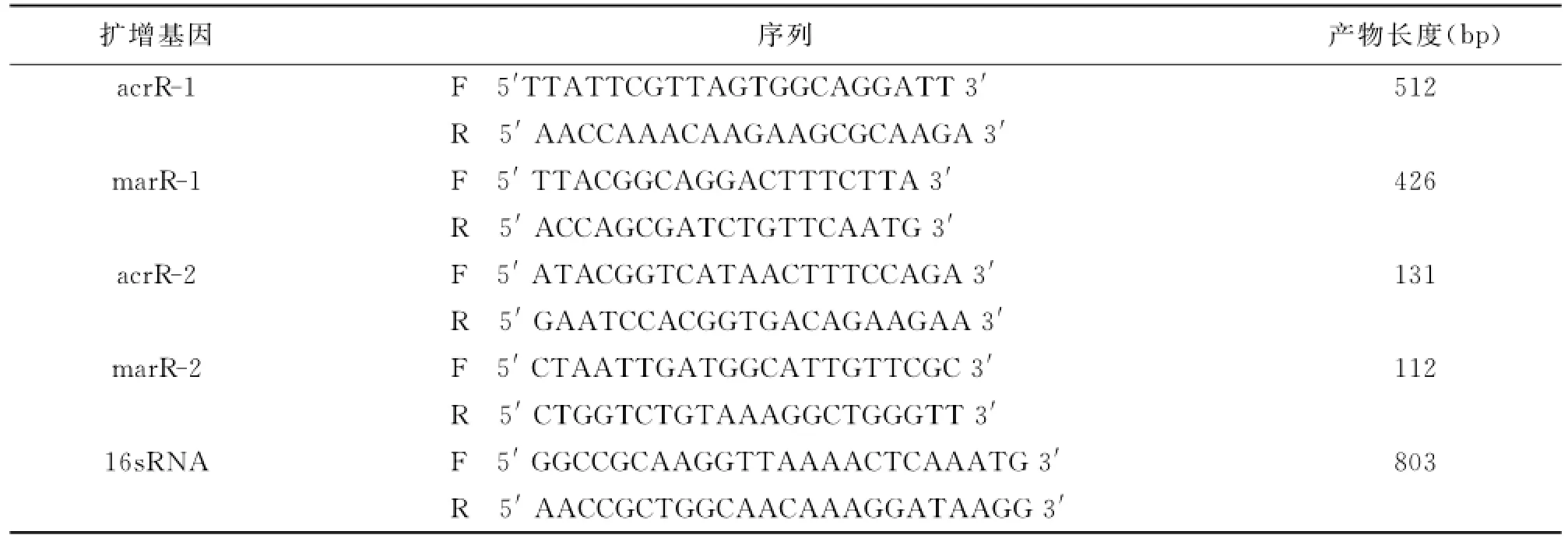
表1 PCR引物序列
2 結果
2.1 環丙沙星誘導結果
經過69代傳代誘導,環丙沙星對誘導菌株的MIC達64μg/mL。E-test法測定誘導后菌株的MIC,發現經過誘導的敏感菌株除了對環丙沙星產生耐藥外,還對與環丙沙星在結構和作用機制上都不同的其他幾種抗菌藥(氨芐西林、頭孢噻肟、慶大霉素、氯霉素、四環素、復方新諾明)也產生了耐藥,經過環丙沙星的誘導,原來的敏感菌株均逐步轉變為多重耐藥株。誘導前后各抗生素MIC值見表2。
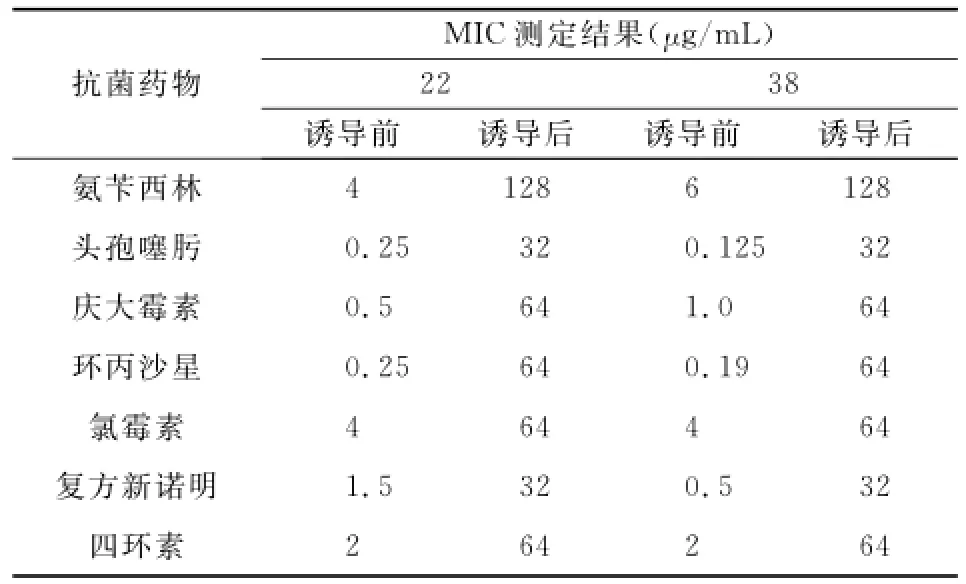
表2 兩株宋內志賀菌誘導前后的MIC結果
2.2 宋內志賀菌acrR和marR的PCR擴增產物及其測序結果
2株誘導株均擴增出大小分別為512bp和426 bp的片段,acrR和marR基因擴增產物,經測序與BLAST比對,均未發現有基因突變。
2.3 宋內志賀菌acrR、marR基因表達水平分析結果
讀取每個菌株誘導前后acrR、marR的Ct值與16sRNA的Ct值,實時熒光定量曲線圖見圖1。根據Ct值的比值,誘導耐藥株的acrR、marR表達水平明顯低于敏感株,見表3。這表明誘導耐藥后菌株的主動外排阻遏基因表達水平降低,相應的主動外排基因acrAB-tolC轉錄水平提高,證實了誘導劑環丙沙星激活了菌株的主動外排系統。
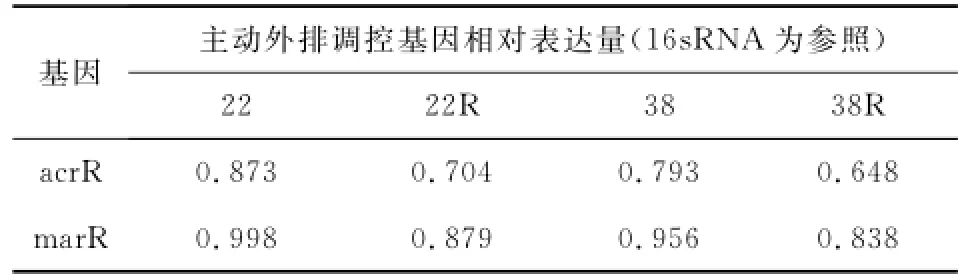
表3 誘導前后acrR和marR的相對表達量
3 討論
自Levy[7]等提出主動外排系統是細菌主要耐藥機制之一后,一個個新的主動外排系統不斷被發現,主動外排系統在臨床主要致病菌耐藥中的重要作用不斷被揭示,并為業界所認同,是目前細菌耐藥機制研究中的新熱點[8]。
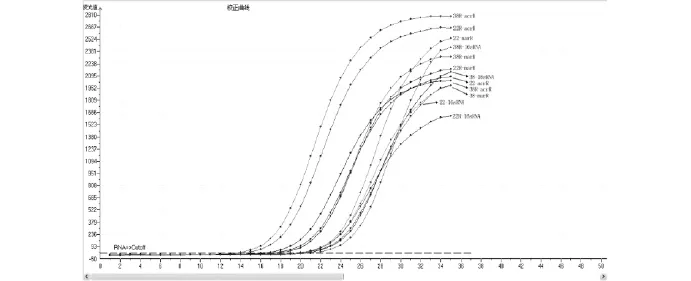
圖1 實時熒光定量PCR曲線圖
本實驗用環丙沙星對宋內志賀菌敏感菌株進行誘導耐藥,建立環丙沙星耐藥株,意外的是誘導后菌株除了對環丙沙星產生耐藥外,還對與環丙沙星在結構和作用機制上都不同的其他幾種抗菌藥(氨芐西林、頭孢噻肟、慶大霉素、氯霉素、四環素、復方新諾明)也產生了耐藥,而且這幾種藥物為主動外排泵AcrAB的作用底物。推測這可能是由于誘導劑環丙沙星激活了敏感株的主動外排系統,使敏感株變為多重耐藥菌株。
AcrAB的表達受多種調節因子的調節,其中阻遏子基因acrR和marRAB操縱子的作用最為重要。AcrR抑制子是由位于泵基因上游的阻遏子基因arcR編碼的,對acrAB轉錄起一定程度的抑制作用[3,4]。AcrR通常以二聚體形式結合于acrA基因和acrB基因啟動子附近的位點,抑制acrA和acrB基因的轉錄,防止其過量表達;marRAB操縱子包括啟動子marO、阻遏基因marR、轉錄調節因子marA和marB。該操縱子可編碼三種蛋白質,MarR(阻遏子)、MarA(轉錄激活物)和MarB(功能不明蛋白質),marRAB操縱子被激活后,可增強細菌多重耐藥性。MarR是由marR基因編碼的162個氨基酸大小的蛋白質,它可以結合到marRAB操縱子調控區域,抑制MarA的產生,從而抑制主動外排泵基因的轉錄[9]。有研究報道當acrR或marR基因突變后,不能有效阻遏acrAB的轉錄,導致acrAB表達增強,開啟主動外排系統,導致因藥物外排而產生的耐藥[3,4,10]。本項目中對兩株誘導株中的acrR和 marR基因進行測序,均未發現有基因突變發生,但通過定量分析,發現誘導耐藥株中acrR表達水平下降時,細菌對多種抗生素產生耐藥,推測可能是主動外排系統被激活。本實驗中未發現acrR和marR的基因突變,可能與實驗株有限相關。宋內志賀菌的主動外排泵AcrAB-TolC的影響因素很多,外排機制比較復雜。由于宋內志賀菌標本較少,耐藥宋內志賀菌標本更難獲得,本試驗僅就兩株誘導耐藥宋內志賀菌的主動外排泵相關的阻遏基因做了研究,將來需廣泛收集標本,擴大檢測例數作進一步研究。
[1]Blair J M,Piddock LJ.Structure,function and inhibition of RND efflux pumps in Gram-negative bacteria:an update[J].Curr Opin Microbiol,2009,12(5):512.
[2]王松泰,張繼瑜,魏小娟,等.福氏志賀菌主動外排基因acrA的克隆及原核表達[J].中國人獸共患病學報,2008,24(10):933.
[3]Pourahmad JR,Jazayeri N.Expression of acrA and acrB Genes in Esherichia coli Mutants with or without marR or acrR Mutations[J].Iran J Basic Med Sci,2013,16(12):1254.
[4]Ballesté-Delpierre C,SoléM,Domènech?,et al.Molecular study of quinolone resistance mechanisms and clonal relationship of Salmonella enterica clinical isolates[J].Int J Antimicrob Agents,2014,43(2):121.
[5]Kim JY,Kim SH,Jeon SM,et al.Resistance to fluoroquinolones by the combination of target site mutations and enhanced expression of genes for efflux pumps in Shigella flexneri and Shigella sonnei strains isolated in Korea[J].Clin Microbiol Infect,2008,14(8):760.
[6]Yamasaki S,Nagasawa S,Hayashi-Nishino M,et al.AcrA dependency of the AcrD efflux pump in Salmonella enterica serovar Typhimurium[J].J Antibiot(Tokyo),2011,64(6):433.
[7]Levy SB.Active efflux mechanisms for antimicrobial resistance[J].Antimicrob Agents Chemother,1992,36(4):695.
[8]張 永,唐英春.病原菌質子驅動型外排泵分子機制研究進展[J].國外醫藥(抗生素分冊),2004,25(1):6.
[9]Duval V,McMurry LM,Foster K,et al.Mutational analysis of the multiple-antibiotic resistance regulator MarR reveals a ligand binding pocket at the interface between the dimerization and DNA binding domains[J].J Bacteriol,2013,195(15):3341.
[10]Sato T,Yokota S,Uchida I,et al.Fluoroquinolone resistance mechanisms in an Escherichia coli isolate,HUE1,without quinolone resistance-determining region mutations[J].Front Microbiol,2013,24(4):125.
Changes of repressor gene acrR and marR induced resistant Sonnei Shigella
ObjectiveTo analyze mutation and expression of repressor gene acrR and marR for acrAB-tolC efflux system before and after induced resistant to Ciprofloxacin.Methods The resistance to Ciprofloxacin was induced step by step with 1/2MIC,The total DNA and RNA was extracted from the induced and the primary strains,and then detected the acrR and marR gene mutation and expression with sequencing and real time PCR.Results The results showed that the acrR and marR were not mutated,but declined in two induced sonnei shigellacompared with the wild strains.Conclusions Its indirectly means that the acrAB-tolC transcription declined and the active efflux was activated by Ciprofloxacin.
R378.2+5
A
2014-10-09)
1007-4287(2015)10-1625-05
GAO Pei-zeng,YAO Ming-xiao.
(ClinicalLaboratory of Ji’nan Fourth People’s Hospital,Ji’nan250031,China)
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
