4 種禽源病毒感染CEF 后β -catenin 基因的動態表達
2015-03-18 07:48:20傅安靜黃名英
中國獸醫雜志 2015年5期
傅安靜,黃名英,高 艷,文 紅
(1.成都農業科技職業學院,四川 成都611130 ;2.西南民族大學,四川 成都610041 ;3.成都溫江區農法局動物檢疫疾病控制中心,四川 成都 611130)
新城疫病毒(NDV)、高致病性禽流感病毒(AIV)、禽白血病病毒(ALV)、網狀內皮增生癥病毒(REV)嚴重危害養禽業。利用雞胚成纖維細胞(CEF)探討病毒與宿主相互作用的機制,對研究病毒感染及宿主抗病機理有重要意義。β連環蛋白(β-catenin)是Wnt/β-catenin 信號通路的一個關鍵參與者,Wnt/βcatenin 信號通路在細胞增殖分化、人類惡性腫瘤的發生發展中都有重要作用[1]。而有關病毒感染與Wnt/β-catenin 信號之間關系的研究多集中于丙型肝炎病毒(HCV)、人類皰疹病毒4 型(EBV)等致瘤性慢病毒,通過多種機制增加β-catenin 積累,激活其信號傳導,從而促進細胞惡性轉變及病毒感染伴隨的癌癥的發生[2-4]。ALV、REV感染后對β-catenin 表達的影響是否與人致瘤病毒相似;NDV、AIV感染后對βcatenin 表達的影響又如何,都未見文獻報道。因此,本試驗對這4 種病毒感染CEF 后β-catenin mRNA的動態表達進行檢測,為探討這幾種病毒感染與機體的相互作用提供參考。
1 材料與方法
1.1 毒株和雞胚 新城疫強毒株F48E9(NDVF48E9),購自中國獸醫藥品監察所;H5N1 亞型禽流感病毒(AIVH5N1)A/Duck/Guangdong/8/2001、禽網狀內皮增殖癥病毒(REV)SD9901 和禽白血病病毒(ALV)NX0101,均由中國動物衛生及流行病學中心提供。SPF 雞胚,購自青島易邦生物公司,由本實驗室孵育至10 日齡后使用。
1.2 主要儀器 實時熒光定量PCR儀ABI 7300(ABI公司,美國);凝膠成像系統Doc2000(Bio-Rad 公司,美國);紫外分光光度計DU800(Beckman公司,美國);高速冷凍離心(Eppendorf 公司,德國)。
1.3 主要試劑 SYBR?Premix ExTaqTM,購自日本TaKaRa 公司;TRIZol Reagent,購自Invitrogen 公司;RT-PCR 試劑盒,購自博瑞克公司;DMEM/high 培養基、0.25%胰酶、小牛血清,均購自Hyclone 公司。
1.4 病毒感染CEF和采樣 按文獻[5]方法進行CEF的培養,待細胞長成單層后,繼一代至24 孔板上,每孔接5×105個細胞。待細胞在24孔板上長成單層后,分別接100 個TCID50(組織細胞半數感染量)的病毒NDVF48E9、AIVH5N1、ALV和REV,同時設對照組,對照組加同樣體積的Hank′s 液。NDVF48E9 和AIVH5N1 37 ℃感作30 min,ALV和REV感作60 min,添加含2%小牛血清的DMEM維持液,繼續培養。NDVF48E9 和AIVH5N1 在接毒后3、6、9、12、24 h,ALV和REV在接毒后24 h、3、5、7 d 和8 d,每個時間點接毒組和對照組都設3 個復孔,加TRIZol 收集細胞,細胞于-80 ℃保存。
1.5 RNA的提取及質量檢測與cDNA的合成 按說明書進行RNA的提取,電泳檢測其完整性以及通過核酸蛋白檢測儀測OD260/280的比值和RNA的濃度。cDNA的合成按照反轉錄試劑說明書進行。
1.6 β-catenin mRNA表達水平的檢測及數據統計和分析 參考本實驗室優化篩選的內參基因,以核糖體蛋白亞基4(RPL4)作為內參基因。用Rrt-PCR方法對樣本中β-catenin mRNA表達量進行檢測,引物序列為PF:5′-TGCTACAGCCGTTTCTAAT-3′,PR:5′-GCTACAATAACTTTGGGATAA-3′,反應體系為:SYBR Green I 10 μL,ROX 0.4 μL,PF、PR 各0.5 μL,cDNA2 μL,去離子水6.6 μL,共20 μL 反應體系。反應條件為:95 ℃5 min;95 ℃30 s,52 ℃30 s,72 ℃30 s,共40 個循環。對細胞樣品中β-catenin mRNA的表達量進行相對定量[6],同時用t檢驗對數據進行統計學處理。以對照組細胞的β-catenin mRNA表達量為“1”,接毒組是對照組的倍數。
2 結果
2.1 病毒感染后細胞病變情況及病毒的Rrt-PCR檢測結果 NDVF48E9 接毒組細胞9 h 開始出現細胞病變,24 h 時細胞病變達到60%,30 h 時細胞病變達到90%;AIVH5N1 接毒組細胞12 h 時開始出現細胞病變,24 h 時細胞病變達到40%,30 h時細胞病變達到80%;ALV和REV接毒組細胞無細胞病變和死亡,但是ALV和REV都是在感染CEF 后7 d 檢測到病毒;對照組無細胞病變和死亡,說明4 種病毒成功感染CEF。接毒后的細胞形態見圖1。
2.2 RNA質量檢測結果 RNA樣本的電泳檢測見圖2,結果顯示,3 條帶分別為5 SrRNA、18 SrRNA、28 SrRNA;所有RNA樣本的核酸蛋白檢測儀檢測OD260/280比值都在1.8~2.0 之間。說明所提取的RNA符合要求。
2.3 4 種禽源病毒感染后β-catenin mRNA的動態表達
2.3.1 AIVH5N1 感染后β-catenin mRNA的動態表達 AIVH5N1 感染CEF 后β-catenin mRNA的表達水平如圖3 所示。結果顯示,AIVH5N1 感染后3h 誘導β-catenin mRNA的表達,接毒組是對照組的3.14 倍,差異極顯著(P<0.01);接毒后6 h 時略高于對照組,是對照組的1.12 倍,無統計學意義;隨后β-catenin mRNA的表達水平降低,接毒后9、12、24 h 時接毒組分別是對照組的0.5、0.03、0.3 倍,其中12 h 時差異極顯著(P<0.01)。

圖1 接毒后各組細胞形態
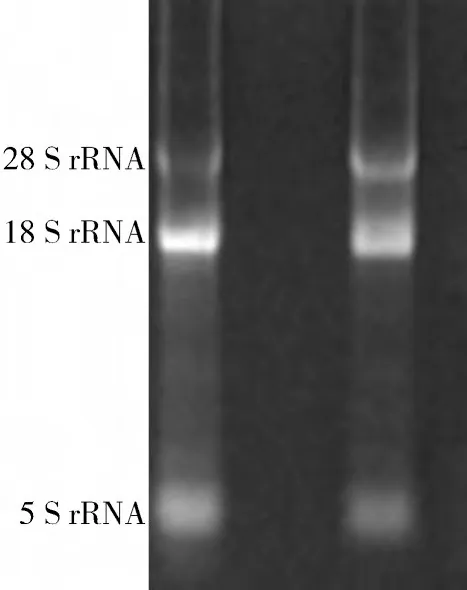
圖2 R NA電泳結果
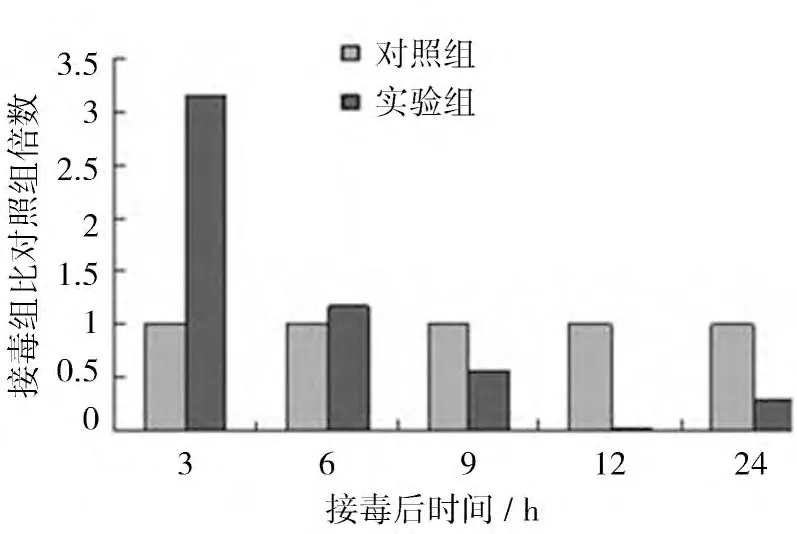
圖3 AIV感染CEF 后β-cateninmR NA的表達量比對照組
2.3.2 NDVF48E9 感染后β-catenin mRNA的動態表達 NDVF48E9 感染CEF 后β-catenin mRNA的表達水平如圖4 所示。結果顯示,NDVF48E9 感染后3 h 誘導β-catenin mRNA的表達,接毒組是對照組的1.55 倍,無統計學意義;隨后,接毒組細胞β-catenin mRNA表達水平降低,感染后6、9、12、24 h 接毒組β-catenin mRNA表達水平分別是對照組的0.84、0.22、0.53、0.25 倍,其中接毒后9、24 h 時差異顯著(P<0.05)。
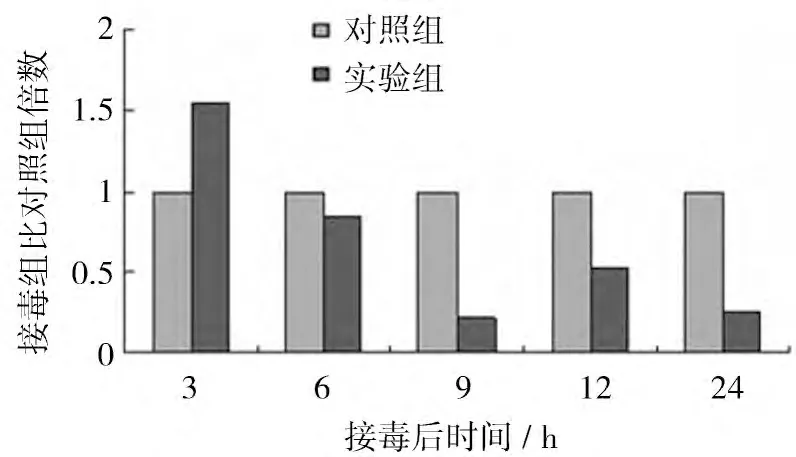
圖4 NDV感染CEF 后β-cateninmRNA的表達量比對照組
2.3.3 ALV感染后β-catenin mRNA的 動態 表達ALV感染CEF 后β-catenin mRNA的表達水平如圖5所示。結果顯示,ALV感染后24 h,接毒組β-catenin mRNA的表達水平略低于對照組,是對照組的0.82倍,無統計學意義;隨后誘導β-catenin mRNA的表達,接毒后3、5、7、8 d時,接毒組細胞β-catenin mRNA的表達水平分別是對照組的6.20、3.11、6.70、2.21倍,其中3、7 d 時差異極顯著(P<0.01),5 d 時差異顯著(P<0.05)。
2.3.4 REV感染后β-catenin mRNA的動態表達
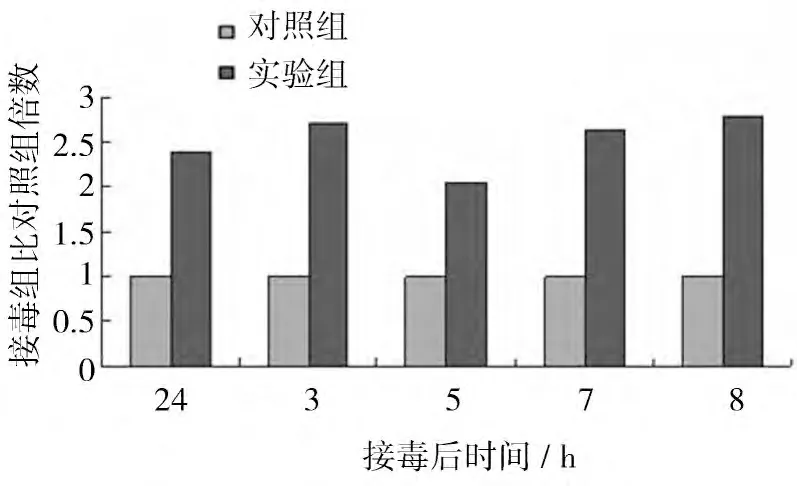
圖5 ALV感染CEF 后β-cateninmR NA的表達量比對照組
REV感染CEF 后β-catenin mRNA的表達水平如圖6 所示。結果顯示,REV感染后誘導β-catenin mRNA的表達,接毒后24 h、3、5、7、8 d 接毒組βcatenin mRNA的表達水平分別是對照組的2.38、2.71、2.05、2.64、2.80 倍,差異均顯著(P<0.05)。
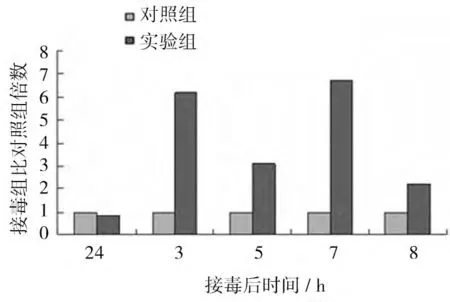
圖6 R EV感染CEF 后β-cateninmR NA的表達量比對照組
3 討論與小結
4 種禽源病毒感染后對CEF 內β-catenin mRNA表達的影響 本研究顯示,β-catenin 參與NDV、AIV、REV和ALV的感染應答,但表達模式不同:NDV、AIV感染初期都誘導β-catenin mRNA的表達,感染中、后期抑制β-catenin mRNA的表達;REV和ALV感染后均誘導β-catenin mRNA的表達。
一些病毒的體內或體外感染研究中,感染后抑制機體β-catenin 表達的報道很少見。GSK-3β是β-catenin 降解復合體的重要成員之一,是βcatenin 的負調節物,該酶激活會抑制β-catenin 的積累[7]。Wnt/β-catenin 信號傳導途徑下游的效應物TCF-4 可能與HIV形成多蛋白復合體,導致HIV轉錄的抑制,而通過轉染顯性負相突變體阻斷TCF-4 或β-catenin 的活性,則又增強HIV的復制[8]。這些有關HIV的資料表明,病毒感染后有可能通過一些機制抑制Wnt/β-catenin 信號傳導而致病,或誘導細胞凋亡,或增強病毒復制。NDV感染后能引起宿主細胞凋亡,而研究表明,NDV引起的細胞病變效應是由凋亡引起的[9-10]。AIVH5N1 感染也會誘導細胞凋亡,包括鴨胚成纖維細胞(DEF)、人氣管上皮細胞、人肺癌細胞系A549、MDCK 細胞等,研究表明,H5N1 亞型AIV導致哺乳動物氣管上皮細胞死亡是由于細胞凋亡的誘導[11-13]。Wnt/β-catenin 信號通路參與調控細胞凋亡[14],這就提示了Wnt/β-catenin 信號通路在NDV、AIV感染誘導細胞凋亡,引發細胞病變死亡中發揮作用的可能性。
REV和ALV感染CEF 后持續誘導β-catenin mRNA的表達,這與一些人類致瘤性病毒感染后βcatenin 的表達模式相似。丙型肝炎病毒(HCV)的非結構蛋白5A(NS5A)可通過激活PI3K 和Akt 激酶而刺激GSK-3β的磷酸化,使其失活,導致β-catenin的積累和β-catenin 依賴性轉錄的增強;或者可能單獨與p85形成復合體,與β-catenin 直接作用,激活βcatenin 信號[8-9]。EBV的潛伏膜蛋白(LMP)同樣可通過一些途徑激活β-catenin 信號,導致β-catenin 的高表達和積累,成為其致細胞惡性轉變和腫瘤產生的誘因[4]。REV和ALV都是致瘤性慢病毒,其感染后β-catenin mRNA的表達模式與人類致瘤病毒相似,由此可推測Wnt/β-catenin 信號通路可能也在REV和ALV的致病中發揮了重要作用。
這4 種病毒感染后通過何種蛋白成分、何種途徑影響Wnt/β-catenin 信號通路,作用如何都有待研究,但上述結果為進一步研究病毒感染致病與機體相互作用的分子機制提供了有價值的參考。
[1]Nusse R.Wnt signaling in disease and in development[J].Cell Res,2005,15(1):28-32 .
[2]Milward A,Mankouri J,Harris M.Hepatitis Cvirus NS5Aprotein interacts with beta-catenin and stimulates its transcriptional activity in a phosphoinositide-3 kinase-dependent fashion[J].JGen Virol,2010,91(2):373-381.
[3]Park CY,Choi SH,Kang SM,et al.Nonstructural 5Aprotein activates beta-catenin signaling cascades:implication of hepatitis Cvirus-induced liver pathogenesis[J].JHepatol,2009,51(5):853-864.
[4]Tomita M,Dewan MZ,Yamamoto N,et al.Epstein-Barr virusencoded latent membrane protein 1 activates beta-catenin signaling in Blymphocytes[J].2009,100(5):807-812
[5]殷震,劉景華.動物病毒學[M].2 版.北京:科學出版社,1997:223-224.
[6]Vandesompele J,De Preter K,Pattyn F,et al.Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes[J].Genome Biol,2002,3(03):43.
[7]吳桐,龐小穎,張浩.Wnt 信號途徑的分子機制[J].食品與藥品,2006,8(7):5-8.
[8]Kumar A,Zloza A,Moon R T,et al.Active beta-catenin signaling is an inhibitory pathway for human immunodeficiency virus replication in peripheral blood mononuclear cells[J].JVirol,2008,82(6):2813-2820.
[9]Ravindra P V,Tiwari AK,Sharma B,et al.HN protein of Newcastle disease virus causes apoptosis in chicken embryo fibroblast cells[J],Arch Virol,2008,153:749-754.
[10]Ravindra P V,Tiwari AK,Ratta B,et al.Newcastle disease virus-induced cytopathic effect in infected cells is caused by apoptosis[J].Virus Research,2009,141:13-20.
[11]Ueda M,Daidoji T,Du A,et al.Highly pathogenic H5N1 avian influenza virus induces extracellular Ca2+influx,leading to apoptosis in avian cells[J].JVirol,2010,84(6):3068-3078.
[12]Lam WY,Tang JW,Yeung AC,et al.Avian influenza virus A/HK/483/97(H5N1)NS1 protein induces apoptosis in human air-way epithelial cells[J].JVirol,2008,82(6):2741-2751.
[13]Yng W,Qu S,Liu Q,et al.Avian influenza virus A/chicken/Hubei/489/2004(H5N1)induces caspase-dependent apoptosis in a cell-specific manner[J].Mol Cell Biochem,2009,332(1-2):233-241.
[14]韓琳,馮新港.Wnt 信號通路及其在動物生長發育過程中的作用[J].中國獸醫寄生蟲病,2008,16(3):47-52.
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
人大建設(2019年12期)2019-05-21 02:55:32
電子制作(2018年11期)2018-08-04 03:25:42
海峽科技與產業(2016年3期)2016-05-17 04:32:12
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
