華根霉蛋白質組雙向電泳體系的建立及優化
2015-01-06 09:44:56鄭禮月
食品與生物技術學報 2015年5期
鄭禮月, 王 棟*, 徐 巖
(1.江南大學 工業生物技術教育部重點實驗室,江蘇 無錫 214122;2.江南大學 生物工程學院,江蘇 無錫214122)
華根霉蛋白質組雙向電泳體系的建立及優化
鄭禮月1,2, 王 棟*1,2, 徐 巖1,2
(1.江南大學 工業生物技術教育部重點實驗室,江蘇 無錫 214122;2.江南大學 生物工程學院,江蘇 無錫214122)
華根霉(Rhizopus chinensis CCTCC M201021)是從我國傳統微生物載體——大曲中分離篩選得到的一株絲狀真菌,具有重要的工業應用前景。為建立一種適于華根霉胞內蛋白質的雙向電泳體系以進行蛋白質組學研究,作者對華根霉胞內蛋白質的提取方法及雙向電泳流程相關參數進行了考察和優化.確定采用液氮研磨與高速珠磨相結合的方法對華根霉進行細胞破碎,用TCA/丙酮對提取的蛋白質進行純化,采用被動水化的上樣方式上樣,24 cm、pH 4~7的線性IPG膠條,上樣量為1 200 μg蛋白質,考馬斯亮藍G-250膠體染色法,最終得到了背景清晰、分辨率高的雙向電泳圖譜。
華根霉;全蛋白;雙向電泳
絲狀真菌是一類重要的工業微生物,具有復雜的生理生化和代謝特性,多年以來其發酵產品已廣泛應用于食品、醫藥、農業、能源、化工等眾多領域.華根霉(CCTCC M201021)是從我國傳統釀造微生物載體——白酒大曲中分離篩選得到的一株很有特點的絲狀真菌,在分類學上屬于毛霉目根霉屬。華根霉可發酵產生多種酶和代謝產物[1],具有較為復雜的代謝調節機制,其產生的脂肪酶能在非水相中催化酯合成、酯交換、酸解、醇解和氨解反應,具有重要的工業應用價值[2]。華根霉作為一種絲狀真菌,液態發酵中會形成不同的菌體形態,進而影響合成活性脂肪酶的發酵水平[3]。目前針對絲狀真菌的發酵研究主要停留在發酵條件[4]、培養基成分優化[5]和蛋白質的異源表達[6]等方面,其生理代謝特點、菌體形態與其代謝產物間分子層面的生物學機制目前尚不清楚。
隨著后基因組時代的來臨,以基因組學、轉錄組學、蛋白質組學等為代表的系統生物學技術和方法為理解絲狀真菌的形態問題提供了強有力的手段。近年來,隨著曲霉、根霉等微生物全基因組的解析[7],對于絲狀真菌的的認識除了基本的代謝和細胞功能外,諸如細胞壁分化、蛋白質分泌機制等生理現象和問題也得到了更深入的認識和理解。相比于基因組學,蛋白質組學能直接從蛋白質水平上反映細胞內蛋白質表達量的變化,因而蛋白質組學正被廣泛的應用于微生物領域的研究。雙向電泳技術是蛋白質組學中最常見一種實驗技術,因其高靈敏度,高分辨率和高通量的優勢[8]被廣泛應用。然而,因雙向電泳實驗步驟繁雜及參數不確定性等原因,很難找到一種通用于所有生物的雙向電泳條件。
目前,針對絲狀真菌的蛋白質組學研究主要集中在曲霉上,根霉由于其細胞破碎困難等原因而鮮有報道。因而本研究的目的是創建和優化華根霉的胞內蛋白質雙向電泳體系,為研究菌體形態與代謝產生的分子機制提供技術支持,同時也為其他霉菌的蛋白質組學研究提供參考。
1 材料與方法
1.1 實驗材料
1.1.1 菌種與培養條件
1)菌株:華根霉 (Rhizopus chinensis CCTCC M201021)由江南大學釀造微生物與應用酶學研究室篩選保藏。
2)斜面培養基:馬鈴薯—葡萄糖瓊脂培養基;發酵培養基:麥芽糖10 g/L,蛋白胨40 g/L,ΜgSO4· 7H2O 0.5 g/L,K2HPO4·3H2O 2 g/L,橄欖油20 g/L;初始pH 5.5。
3)培養條件:種子培養,37℃靜置培養72 h;發酵培養,30℃ 旋轉式搖床200 r/min培養72 h,每升培養基接種5×107個孢子。
1.1.2 實驗試劑IPG膠條 (pH 3~10/4~7,24 cm,線性)、兩性電解質IPG buffer、非干擾性蛋白質濃度測定試劑盒2-D Quant Kit:美國GE公司產品;礦物油:Bio-Rad公司產品;二硫蘇糖醇(DTT)、溴酚藍、N,N,N’,N’-四甲基乙二胺(TEMED)、硫脲、碳酸氫銨 (NH4HCO、α-氰基-4-羥基肉桂酸(HCCA)、胰蛋白酶、乙腈(ACN):Sigma公司;低熔點瓊脂糖、尿素、三氟乙酸(TFA)、三氯乙酸(TCA)、蛋白酶抑制劑、甲叉雙丙烯酰胺、過硫酸銨(APS)、十二烷基硫酸鈉(SDS)、丙烯酰胺、3-[(3-膽固醇氨丙基)二甲氨基]-1-丙磺酸 (CHAPS)、碘乙酰胺(IAA)、考馬斯亮藍 G-250:上海生工產品;其它試劑均購自國藥。
1.1.3 實驗儀器EttanTMIPGphor3等電聚焦儀:瑞典GE公司;EttanTMDALTSix垂直電泳系統:瑞典GE公司;ImageScanner掃描儀:瑞典GE公司;蛋白質電泳系統(Mini-Protein3 Cell):美國Bio-Rad公司;臺式35K高速冷凍離心機:美國Sigma公司;Ultraflextreme飛行時間質譜儀:德國Bruker公司。
1.2胞內蛋白質提取
1.2.1 細胞破碎方法
1)液氮研磨法:收集發酵72 h的菌體,超純水清洗菌體,5 000 r/min離心5 min,重復洗3次。稱濕菌體質量,將濕菌體與玻璃珠以1∶3質量比混合后加入液氮,研磨至粉狀。取裂解液1 mL,每毫升裂解液中加入10 μL蛋白酶抑制劑 (Protein Cocktail for fungi)、10 μL 20%IPG buffer和1.2 g玻璃珠與菌體混合物,于4℃下放置12 h,期間每2小時拿出渦旋2 min。
2)液氮研磨+Mini beads beater破碎法:取裂解液1 mL,每毫升裂解液中加入10 μL蛋白酶抑制劑(Protein Cocktail for fungi)、10 μL 20%IPG buffer和1.2 g玻璃珠與菌體混合物,渦旋混勻后置于mini beads beater破碎儀破碎。破碎后固液混合物于4℃放置12 h,期間每2小時拿出渦旋2 min。
3)Mini beads beater破碎法 (高速珠磨法):將(2)中1.2 g玻璃珠與菌體混合物替換為0.3 g濕菌體和0.9 g玻璃珠,其余全部與(2)相同。
將上述三種方法得到的固液混合物在4℃、12 000 r/min離心30 min,上清液即為胞內全部蛋白質溶液,蛋白質定量使用美國GE公司的2-D quant kit完成。
1.2.2 蛋白沉淀方法
1)丙酮沉淀法:將細胞破碎后上清液轉移到50 mL離心管中,加入10倍體積丙酮溶液沉淀蛋白,于-20℃放置過夜,14 000 r/min離心30 min得蛋白質沉淀。
2)TCA-丙酮沉淀:蛋白溶液中加入10倍體積10%TCA/丙酮溶液,-20℃沉淀蛋白質 24 h,12 000 r/min、4℃離心30 min。去除上清液,加入與10%TCA/丙酮相同體積的丙酮(-20℃預冷)洗滌沉淀3次,以除去沉淀顆粒中殘留的TCA。將得到的沉淀敞口置于室溫下,使沉淀中殘留的丙酮揮發干凈。
向上述兩種方法收集的沉淀中加入水化液渦旋至蛋白質完全溶解,溶解后在4℃、12 000 r/min離心30 min,收集上清液,使用GE 2-D quant kit對2.1中所得蛋白質濃度進行定量。
1.2.3 雙向電泳雙向電泳實驗參照GE公司的《雙向電泳操作手冊》進行。向定量后的蛋白質溶液中加入體積分數0.2%的IPG緩沖液后開始上樣,24 cm IPG膠條的蛋白質上樣量與上樣體積見表1。IPG膠條20℃水化12 h后進行第一向等電聚焦。
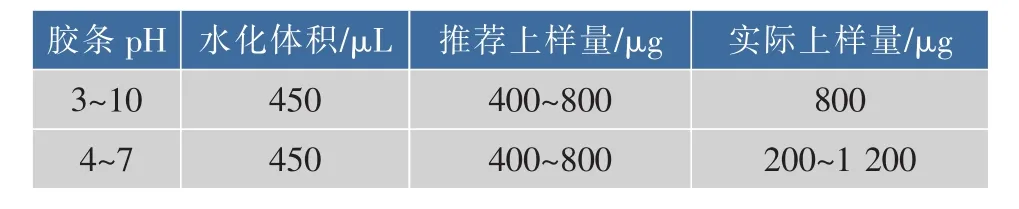
表1 不同IPG膠條的上樣量Table 1 Sampling amount of different IPG strips
將等電聚焦后的膠條在平衡液Ⅰ平衡15 min,再在平衡液Ⅱ中平衡15 min,將平衡后的IPG膠條轉移到SDS-PAGE凝膠上,用低熔點瓊脂糖封膠液封膠后進行第二向SDS-PAGE。此過程采用恒功率進行,首先為2 W/膠,進行一段時間后改為14 W/膠。凝膠染色采用改進后的膠體考馬斯亮藍染色法[9],雙向電泳圖譜分析采用Pdquest8.0.1軟件。
2 結果與討論
2.1 不同細胞破碎方法對胞內蛋白質提取效果比較
華根霉因其堅實的細胞壁導致其胞內的微生物很難被釋放出來,因而想要獲得華根霉胞內全蛋白質,選取一種合適的方法對華根霉進行破壁是進行蛋白質組學研究的先決條件。本研究的目的是得到華根霉的胞內全蛋白質,以期探索出適合華根霉胞內蛋白質提取的最佳方法,為華根霉下一步蛋白質組學研究提供基礎。
關于真菌胞內蛋白質提取的方法,國內外已有很多報道[10-11],但由于真菌種類、菌絲類型、細胞壁結構及次生代謝產物等特點各不相同,很難有通用的蛋白質提取方法。作者比較了液氮研磨法、高速珠磨法、液氮研磨與高速珠磨結合法等三種的細胞破碎方法,由圖1可以看出,液氮研磨法和高速珠磨法法對華根霉細胞壁的破碎都有一定作用,能釋放華根霉部分胞內蛋白質。從蛋白質總量來看,高速珠磨法的效果要好于液氮研磨法。但是將兩種方法結合起來發現,相同質量的細胞菌體,提取到的蛋白質的總量遠遠大于單獨使用兩種破碎方法。說明這兩種方法對華根霉有一定的破壁作用,但破碎并不完全,仍有一部分華根霉并沒有被破碎,導致了其胞內蛋白質沒有被完全釋放,提取到的蛋白質僅為胞內部分蛋白質,為了從整體上對胞內全蛋白進行考察分析,避免胞內蛋白質提取不充分對后續的雙向電泳圖譜及蛋白質組的分析的影響,作者采取液氮研磨法與高速珠磨法相結合的破壁方法對華根霉進行細胞破碎。
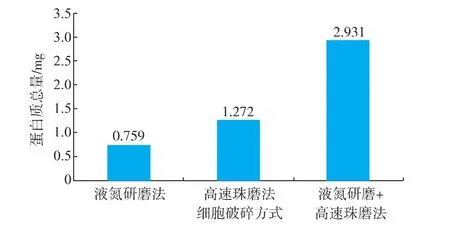
圖1 不同破碎方法得到的蛋白質總量Fig.1 Protein amount obtained by different cell disruption methods
2.2 細胞破碎時間對胞內蛋白質提取效果的影響
早期研究發現菌體與石英砂的比例、菌懸液的濃度、破碎時間等都會影響細胞的破碎率[11]。作者采取固定菌體與石英砂的比例和菌懸液的濃度兩個因素來優化破碎時間,提高破碎效果。另外,在細胞破碎過程中,細胞自有的蛋白酶也會被釋放出來,水解樣本中的蛋白質,極大地增加雙向電泳結果分析的難度。因此,在細胞破碎和隨后的制備過程中,應防止樣本中的蛋白質被水解[12],因而需加入蛋白酶抑制劑以抑制蛋白酶活性。作者在優化破碎時間時,考察了兩種濃度的蛋白酶抑制劑對最終蛋白質提取效果的影響。由圖2可知,在最開始蛋白質質量濃度隨破碎時間的增加而不斷增大,當破碎時間到達4 min后,破碎時間的延長對蛋白質質量濃度的增加的影響變小,說明4 min時細胞的破碎率達到了最大。另外,從圖2可以看出,蛋白酶抑制劑的量對胞內蛋白質質量濃度的影響較小,因此確定蛋白酶抑制劑的添加量為10 μL/mL,破碎時間為4 min。
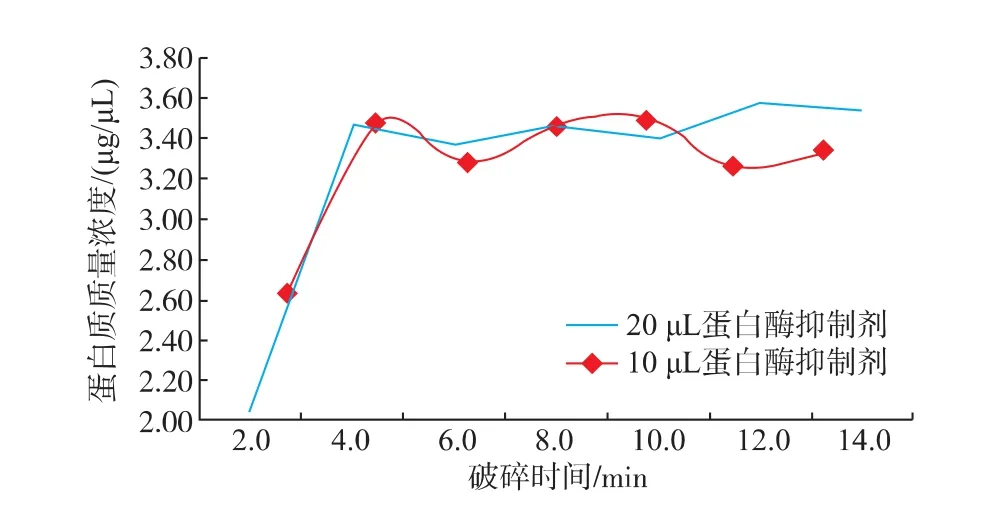
圖2 蛋白質質量濃度與破碎時間的關系Fig.2 Protein concentration varies along with broken time
2.3 不同凝膠參數對雙向電泳圖譜的影響
預制IPG膠條pH范圍的選擇能顯著影響雙向電泳圖譜,寬pH范圍的膠條對蛋白質的覆蓋率大,但是分辨率較低,窄pH范圍的膠條則恰恰相反[13]。鑒于此,作者先用寬范圍即24 cm、pH 3~10的IPG膠條做預實驗,發現華根霉的胞內蛋白質多分布于酸性范圍,堿性范圍內的蛋白質點很少,見圖3。采用pH 4~7的預制膠條進行實驗后發現,蛋白質點多且分布均勻,見圖4,為提高圖譜的分辨率,后續實驗換用pH 4~7的IPG膠條。
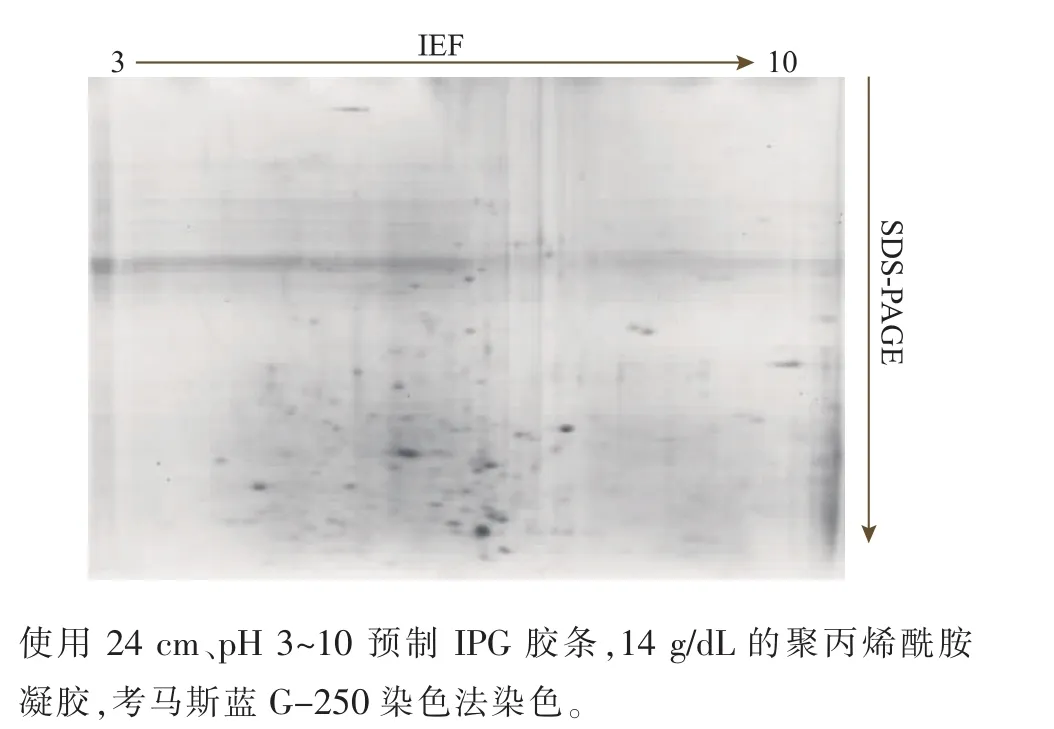
圖3 華根霉雙向電泳圖譜Fig.3 2-DE map of R.chinensis
2.4 不同蛋白質純化方法對雙向電泳圖譜的影響
在提取胞內蛋白質時,細胞內的脂類、核酸、多聚糖等也會一并被提取出來,這些污染物會使雙向電泳圖譜產生明顯的橫、縱條紋,影響結果的可靠性,因而對蛋白質的純化、除雜就顯得尤為重要[14]。在全細胞裂解液中,各種蛋白質的質量濃度范圍差異較大,且呈動態變化,需要根據實驗目的來決定是要高豐度蛋白質還是低豐度蛋白質,進而決定對收集到的蛋白質溶液的后續處理。蛋白質沉淀是一種常見的去除胞內干擾物質的方法,通過這種處理,可以有效除去污染物,使雙向電泳圖像清晰、分辨率高、無明顯條紋,從而保證實驗結果真實可信,揭示潛在的客觀規律。
蛋白質組學中常見的蛋白質沉淀方法有丙酮沉淀法、TCA/丙酮沉淀法,還有Clean-up等試劑盒法,因Clean-up試劑盒的處理量小,處理成本高,而本研究中樣品需求量大,故不采用此方法。作者對比了兩種蛋白質純化方法:丙酮沉淀法和TCA/丙酮沉淀法,見圖4。從雙向電泳圖譜質量來看,利用丙酮對蛋白進行沉淀時,見圖4a。圖譜的酸性端會產生明顯的橫條紋,使得酸性端相當一部分蛋白質不能夠被識別、分析;從蛋白質回收率來看,丙酮沉淀法的回收率為26.34%,而TCA/丙酮沉淀法的回收率為38.19%,丙酮沉淀法的回收效果差,蛋白質丟失率較高,因而本研究選取TCA/丙酮沉淀法作為蛋白質的純化方法。
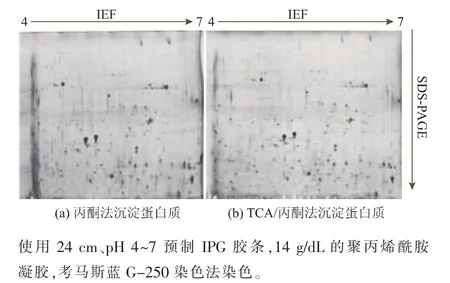
圖4 不同蛋白質沉淀方法所得華根霉雙向電泳圖譜Fig.4 2-D mapsofR.chinensiswith different precipitation methods
2.5 不同蛋白質上樣方式對雙向電泳圖譜的影響
除樣品制備外,蛋白質上樣方式對雙向電泳圖譜的清晰度、分辨率等有著直接的影響[13]。常見的上樣方式有主動水化上樣和被動水化上樣等。作者比較了主動水化上樣和被動水化上樣兩種方式,以期獲得蛋白質點數較多、分辨率較高的雙向電泳圖譜。從圖譜質量看,主動水化上樣后進行等電聚焦時酸性端高相對分子質量區域會有橫條紋出現,見圖5a,而被動水化得到的雙向電泳圖譜酸性區域沒有明顯條紋出現,見圖5b;從蛋白質點數目來看,主動水化的上樣方式得到的蛋白質點數明顯少于被動水化。可能是由于主動水化槽較被動水化槽寬(底面面積較大),過多的蛋白質樣品吸附于水化槽中,使得進入IPG預制膠條的蛋白質量減少,從而導致圖譜上蛋白質點數目過少,確定華根霉胞內蛋白質的上樣方式為被動水化上樣。
2.6 不同蛋白質上樣量對雙向電泳圖譜的影響
分離到盡可能多而且清晰的蛋白質點是進行蛋白質組學的首要條件[15],單就上樣環節而言,在不降低蛋白質分離質量的前提下,大的蛋白質上樣量可以得到數目較多的蛋白質點。一般而言,對于窄pH范圍的IPG膠條,需要比寬pH范圍的IPG膠條上更多的樣品,這是因為pI值不在此范圍內的蛋白質在等電聚焦的過程中會走出膠條。單pH范圍IPG膠條的上樣量往往較大,這樣就可以很好的檢測低豐度的蛋白質。作者采用了400 μg和1 200 μg兩種上樣量來考察上樣量對華根霉雙向電泳圖譜的影響,見圖6。本研究推薦采用大上樣量,其目的在于,如果在大的上樣量的基礎上能夠提高雙向電泳圖譜的質量,可以大大提高蛋白質點的數目并為下游蛋白質點的鑒定奠定基礎。使用pH 4~7范圍的IPG膠條對華根霉胞內蛋白質進行雙向電泳,見圖6。增大了上樣量后,膠圖上沒有明顯條紋及拖丟等現象,可見的蛋白質點數明顯增加,高豐度蛋白質點更加清晰、明顯,低豐度的蛋白點開始被識別,因而在后續的實驗中均采用1 200 μg的蛋白質上樣量。

圖5 不同上樣方式所得華根霉雙向電泳圖譜Fig.5 2-D maps of R.chinensis with different protein loading methods
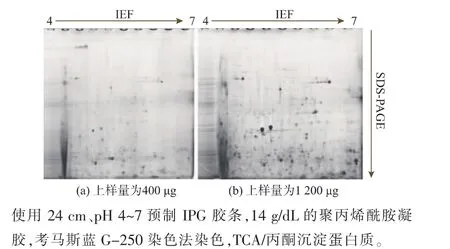
圖6 不同上樣量所得華根霉雙向電泳圖譜Fig.6 2-DE map of protein sample by different loading quantity
2.7重復性實驗
重復性是分離蛋白質組所有蛋白質的關鍵參數之一,雙向電泳步驟繁瑣,實驗周期長,實驗結果往往具有不穩定性。為確保從蛋白質雙向電泳圖譜中獲得可靠的生物學信息,良好的重復性是十分必要的[16]。作者利用已創建條件,對樣品進行了生物學重復,以驗證樣品提取及雙向電泳技術的重復性。由圖7可知,兩個重復樣品的電泳圖譜無明顯橫縱條紋,分辨率較高,蛋白質點的分離效果較好。利用PDQuest8.0.1軟件對雙向電泳圖譜進行分析后發現,圖譜中蛋白質點的匹配率可以達到85.7%(每張膠圖上約800個蛋白質點),證明利用上述已確立的雙向電泳條件不僅可以得到較高分辨率的雙向電泳圖譜,同時具有較好的重復性,所得數據具有統計學意義,能夠真實反映華根霉胞內蛋白質表達水平。
3 結語

圖7 華根霉胞內蛋白雙向電泳圖譜的重復性Fig.7 Repeatability of 2-DE maps with intracellular proteins separated from Rhizopus.chinensis
為了獲得高分辨率、高重復度、高穩定性、無明顯橫縱條紋的雙向電泳圖譜,作者探究了不同的細胞破碎方法、蛋白質提取純化方法、不同凝膠參數、不同上樣量及上樣方式等對華根霉雙向電泳圖譜的影響,創建并優化了適用華根霉的雙向電泳體系。經過對各個條件進行實驗比較,確定華根霉的雙向電泳條件如下:液氮研磨和高速珠磨相結合的方法對華根霉進行細胞破碎,破碎過程中添加真菌專用蛋白酶抑制劑,然后用TCA/丙酮沉淀法對蛋白質樣品進行純化,選擇24 cm、pH 4~7的IPG預制膠條,采取被動上樣方式進行上樣,上樣量為1 200 μg,使用質量濃度為14 g/dL的丙烯酰胺凝膠進行雙向電泳實驗,然后選取改進后的膠體考馬斯藍G-250染色法對凝膠進行染色。所得圖譜經PDQuest軟件分析,每張膠約能分離到800個蛋白質點,該圖譜的創建為后續華根霉蛋白質組學的研究打下了基礎,可為其他霉菌的細胞破碎、蛋白質提取純化方法研究提供參考。
[1]徐巖,趙光鰲,高克亮,等.華根霉產脂肪酶發酵培養基的研究[J].工業微生物,2000,30(3):8-10.
XU Yan,ZHAO Guangao,GAO Keliang,et al.Research of Rhizopus chinensis of lipase production fermentation medium[J]. Industrial Microbiology,2000,30(3):8-10.(in Chinese)
[2]王棟.華根霉(Rhizopus chinensis)非水相合成活性脂肪酶及其酶學特性的研究[D].無錫,江南大學,2008.
[3]Teng Yun,Xu Yan,Wang Dong.Changes in morphology of Rhizopus chinensis in submerged fermentation and their effect on production of mycelium-bound lipase[J].Bioprocess and Biosystems Engineering,2009,32(3):397-405.
[4]朱增亮,王棟,徐巖.發酵罐裝置改進提高華根霉膜結合脂肪酶液態發酵水平[J].工業微生物,2013,43(2):1001-6678.
ZHU Zhengliang,WANG Dong,XU Yan.Fermentation level of membrane-bound lipase produced by Rhizopus chinensis[J]. Industrial Microbiology,2013,43(2):1001-6678.(in chinese)
[5]孫舒揚,王棟,徐巖.固液態發酵中橄欖油對Rhizopus chinensis全細胞脂肪酶的影響[J].微生物學通報,2006,33(4):10-14.
SUN Suyang,WANG Dong,XU Yan.Effects of olive oil on whole-cell lipase production of solid-state and submerged fermentation by Rhizopus chinensis[J].Microbiology China,2006,33(4):10-14.(in chinese)
[6]王睿,喻曉蔚,沙沖,等.定向進化-易錯PCR方法提高華根霉Rhizopus chinensis CCTCC M201021脂肪酶的活力[J].生物工程學報,2009,25(12):1892-1899.
WANG Rui,YU Xiaowei,SHA Chong.Increasing activity of Rhizopus chinensis CCTCC M201021 lipase by directed evolution-error prone PCR[J].Chinese Journal of Biotechnology,2009,25(12):1892-1899.(in Chinese)
[7]Ma,Li-Jun,et al.Genomic analysis of the basal lineage fungus Rhizopus oryzae reveals a whole-genome duplication[J].PLoS Genetics,2009,5(7):e1000549.
[8]廖翔,應天翼,黃留玉,等.蛋白質組學研究中的雙向電泳技術[J].生物技術通訊,2003,14(6):522-524.
LIAO Xiang,YING Tianyi,HUANG Liuyu.Two-dimensional electrophoresis technology in proteomics[J].Letters in Biotechnology,2003,14(6):522-524.(in Chinese)
[9]Candiano,Giovanni.Blue silver:a very sensitive colloidal Coomassie G-250 staining for proteome analysis[J].Electrophoresis,2004,25(9):1327-1333.
[10]Nandakumar.Mark R.Marten Comparison of lysis methods and preparation protocols for one-and two-dimensional electrophoresis of Aspergillus oryzae intracellular[J].Electrophoresis,2002(23):2216-2222.
[11]胡彬彬,林連兵,魏云林,等.一種高效的真菌總蛋白質提取方法[J].中國生物工程雜志,2013,33(9):53-58.
HU Binin,LIN Lianbin,WEI Yunlin.An efficient method of fungus protein extraction[J].China Biotechnology,2013,33(9):53-58.(in Chinese)
[12]Urquhart,Brooke L,et al.'Proteomic contigs'of Mycobacterium tuberculosis and Mycobacterium bovis(BCG)using novel immobilised pH gradients[J].Electrophoresis,1997,18(8):1384-1392.
[13]張麗,應天翼,黃留玉,等.桃果實總蛋白質雙向電泳優化體系的建立[J].食品與生物技術學報,2013,32(3):250-257.
ZHANG Li,YING Tianyi,HUANG Liuyu.Optimization of two-dimensional electrophoresis conditions for proteome analysis of peach fruit[J].Journal of Food Science and Biotechnology,2013,32(3):250-257.(in Chinese)
[14]張國慶,廖杰,于力方.雙向電泳技術在蛋白質組研究中的應用[J].標記免疫分析與臨床,2003,10(3):171-173.
ZHANG Guoqing,LIAO Jie,YU Lifang.Application of two-dimensional electrophoresis in proteomics research[J].Labeled Immunoassay and Clinical,2003,10(3):171-173.(in Chinese)
[15]嚴廣斌.蛋白質組學[J].中華關節外科雜志,2011,5(4):27-27.
YAN Guangbin.Proteomics[J].Chinese Journal Joint Surgeon,2011,5(4):27-27.(in Chinese)
[16]張衛東,郗園林.蛋白質雙向電泳技術分析志賀菌誘導耐多藥相關蛋白[J].河南醫學研究,2010,19(1):1-3.
ZHANG Weidong,XI Yuanlin.Analysis of proteins related to induced multi drug resistance of shigella flexneri by two -mensional electrophoresis[J].Henan Medical Research,2010,19(1):1-3.(in Chinese)
Construction and Optimization of Two-Dimensional Electrophoresis for Proteome Analysis of Rhizopus chinensis
ZHENG Liyue1,2, WANG Dong*1,2, XU Yan1,2
(1.Key Laboratory of Industrial Biotechnology,Ministry of Education,Jiangnan University,Wuxi 214122,China; 2.School of Biotechnology,Jiangnan University,Wuxi 214122,China)
Rhizopus chinensis CCTCCM201021 is an important filamentous fungus,which was isolated from Daqu,a traditional leaven in the production of Chinese liquor.In order to understand this fungal deeply,the conditions of two-dimensional electrophoresis(2-DE)for proteome analysis of R.chinensis were investigated and optimized.Intracellular proteins of Rhizopus chinensis was extracted and separated for 2-DE.Different sample preparation methods,parameters of the gel,sample loading quantity and methods were investigated to optimize 2-DE maps of R.chinensis. Ultimately disrupt cell in liquid nitrogen and mechanical agitation,precipitate proteins with TCA/acetone,passively rehydrate proteins with 24 cm pH 4~7 linear strips were selected.Thesample loading quantity for each strip was 1 200 μg,and modified colloidal coomassie brilliant blue G-250 was applied to stain gels.Finally,2D-gel map with no obvious vertical and horizontal stripes,low background,high resolution and good reproducibility was obtained.Approximately 800 protein spots were isolated from the 2-DE protein map after analyzed by PD-Quest software.
Rhizopus.chinensis,intracellular proteins,two-dimensional electrophoresis
Q 93
A
1673—1689(2015)05—0487—07
2014-01-25
國家973計劃項目(2011CB710800);國家自然科學基金項目(31271920);國家863計劃項目(2012AA022207,2013AA102108);江蘇省高校優勢學科建設工程資助項目。
*通信作者:王 棟(1971—),男,浙江紹興人,工學博士,副教授,碩士研究生導師,主要從事釀酒科學與發酵工程方面的研究。
E-mail:dwang@jiangnan.edu.cn
