基于酵母產孢的syntaxin家族蛋白SNARE區域功能分析
2015-01-06 09:45:12王麗萍邵侃凱高曉冬中西秀樹
食品與生物技術學報 2015年5期
王麗萍, 邵侃凱, 高曉冬, 中西秀樹
(江南大學 生物工程學院,江蘇 無錫214122)
基于酵母產孢的syntaxin家族蛋白SNARE區域功能分析
王麗萍, 邵侃凱, 高曉冬, 中西秀樹*
(江南大學 生物工程學院,江蘇 無錫214122)
真核細胞中大部分膜融合過程由SNARE蛋白介導,但其功能和調節機制尚未完全清楚。酵母孢子形成是研究囊泡融合機制包括SNARE蛋白的理想模型系統。在該過程中涉及t-SNARE蛋白Sso1,它是突觸囊泡融合所需蛋白syntaxin 1A的同源物,兩者SNARE區域有51%的同源性。盡管如此,將SSO1的SNARE區域完全替換成syntaxin 1A,而構建的嵌合體卻無法回補sso1△突變的產孢缺陷。為了確定哪些殘基為Sso1功能所必須,作者進行了嵌合體和突變分析,發現Sso1/syntaxin 1A嵌合體中syntaxin 1A的SNARE區域的220位丙氨酸換成谷氨酸后獲得產孢功能。另外,Sso1發生相應的突變-218位谷氨酸突變成丙氨酸后失去其功能。因此,218位谷氨酸殘基為Sso1產孢功能所必須。
SNARE;人源化;釀酒酵母;Sso1p;syntaxin1A
在真核生物中,囊泡融合機制是高度保守的,SNARE(soluble N-ethylmaleimide-sensitive factor attachment protein receptors;可溶性NSF附著蛋白受體)蛋白是融合蛋白中的重要成員[1-2]。SNARE蛋白是保守性很強的蛋白家族,經常被分為兩組:QSNARE蛋白和R-SNARE蛋白[3]。SNARE蛋白位于特定的膜上,只有當同源的Q-SNARE蛋白和RSNARE蛋白結合時,泡膜才能融合。
在神經元的突觸囊泡融合過程中,syntaxin-1[4]和SNAP-25扮演Q-SNARE角色,synaptobrevin則扮演著R-SNARE角色。當囊泡膜上的synaptobrevin與syntaxin-1和 SNAP-25結合在一起時,突觸囊泡中的神經遞質就會釋放出來,傳遞給突觸后膜[1,3,5-7]。SNARE蛋白不僅和神經遞質釋放有關,也和粘蛋白及血小板升高引起的人類疾病有關。研究表明,抑制SNARE蛋白功能可導致神經退行性疾病[8-9]。SNARE融合過程受到complexin[10]、Tomosyn[11-12]、Munc18/nSec-1[13]等多種蛋白質的調控,但調控機制尚不明確。
酵母細胞中同樣存在 SNARE融合系統,syntaxin-1在突觸囊泡融合過程中的作用機制可利用酵母的SNARE融合系統來研究。
酵母中有兩個 syntaxin蛋白——Sso1p和Sso2p,他們是哺乳動物syntaxin1的同源物且彼此高度相似。Sso1p和Sso2p在酵母營養細胞中共同執行基本功能——與SNARE蛋白Sec9p,Snc1p和Snc2p形成質膜復合體[14]。在孢子形成過程中則特異性地要求Sso1p存在[15-16]。在該過程中Spo20p代替Sec9p,與Sso1p、Snc1p和Snc2p形成復合體,促使紡錘體極點上prospore membrane(PrM)的擴展。Sso1p缺失的酵母因PrM無法擴展而不能形成孢子。
在SNAREs參與的孢子形成過程中,SSO1和SPO20是PrM形成所必需的基因[16]。Hideki Nakanishi等人已研發出監測PrM形成過程中囊泡融合的技術。這類現象也可通過熒光顯微鏡觀察到,并且在活細胞中可看到PrM的延伸[17-18]。因此,PrM的形成過程是研究囊泡融合機理的一個理想操作模型系統。SNARE蛋白基因在進化過程中比較保守,可以將 Sso1p的 SNARE區域替換成 syntaxin1的SNARE區域從而構建人源化的膜融合體系。根據改造得到的菌株不僅可用于在酵母細胞中研究人類突觸囊泡融合機理,還可用于醫療和工業用途,其中包括建立神經肉毒素的檢測系統。而且,它們也是篩選參與突觸囊泡融合新基因的非常有用的工具。
作者通過嵌合體的構建對Sso1p的SNARE區域的產孢功能區進行了定位,并成功構建了sso1△sso2△突變株,為篩選與syntaxin1A作用的蛋白或藥物提供了更為簡便的方法。也為后續人源化SNARE膜融合系統的構建及相關理論研究奠定了基礎。
1 材料與方法
1.1 材料與儀器
1.1.1 菌株和質粒釀酒酵母出發菌株 AN120、HJ3(sso1△spo20△)、質粒pFA6a-kanMX6、質粒pFA6a-his3MX6、質粒pRS314、質粒pRS304、質粒pRS316、質粒pRS426:由作者所在實驗室保存。
1.1.2 主要試劑和儀器KOD-Plus-Neo高保真DNA聚合酶:東洋紡公司;Taq DNA聚合酶、限制性內切酶及T4-DNA Ligase:TaKaRa公司;PCR產物純化試劑盒、DNA瓊脂糖凝膠回收試劑盒及質粒小提試劑盒:上海生工生物公司;其他試劑:均為進口分裝或國產分析純。
1.1.3 培養基
1)YPAD培養基:每升含酵母膏10 g,蛋白胨20 g,葡萄糖20 g,腺嘌呤30 mg;
2)YNB選擇培養基:每升含酵母基礎氮源6.7 g,葡萄糖20 g,補加適量必須元素;
3)YPAce培養基:每升含酵母膏10 g,蛋白胨20 g,乙酸鉀20 g,腺嘌呤30 mg;
4)產孢培養基:每升含乙酸鉀20 g;
5)5-FOA(5-Fluoroorotic Acid)[19]培養基:每升含酵母基礎氮源6.7 g,5-氟乳清酸1 g,葡萄糖20 g,補加適量必須元素;
6)LB-氨芐培養基:每升含酵母膏5 g,胰蛋白胨10 g,NaCl 10 g,氨芐100 mg;
7)G418培養基:每升含酵母膏10 g,蛋白胨20 g,葡萄糖20 g,腺嘌呤30 mg,G418 500 mg。固體培養基添加2 g/dL的瓊脂粉。
1.2 基因敲除
基因敲除方法見文獻 [20]。其中質粒pFA6akanMX6為敲除SSO1的模板,質粒pFA6a-his3MX為敲除SSO2的模板,PCR引物見表1。
表1PCR引物
Table 1 Primers

?
1.3 產孢分析
將含質粒的釀酒酵母在YNB選擇培養基于30℃培養過夜,取500 μL培養液轉接至4.5 mL YPAce培養基中培養20~24 h,收集細胞,無菌水洗后重懸于3 mL產孢培養基中繼續培養30 h,觀察記產孢率。每種質粒轉化兩次,每次選3個單菌落計數,細胞數不少于500。
1.4 PrM的觀察
將以pRS304為載體構建的質粒單酶切,回收所得線性質粒轉化sso1△缺陷株,色氨酸缺陷平板篩選陽性克隆。用pRS426-G20轉化所得菌株,色氨酸、尿嘧啶缺陷平板篩選陽性克隆,按產孢分析方法培養,產孢培養基中培養6、7、8 h時各取500 μL培養液收集細胞,水洗后重懸于50 μL無菌水,加450 μL甲醛,30℃搖床處理30 min。收集細胞,無菌水洗兩次后重懸于100 μL無菌水,取5 μL細胞懸液與5 μL DAPI溶液混合,制片觀察PrM。
1.5 營養細胞生長分析
以pRS316SSO1pr-SSO1轉化sso1△單倍體缺陷株,以篩選所得陽性克隆為出發菌株敲除SSO2,得sso1△sso2△雙缺陷型菌株。將待驗證質粒轉入sso1△sso2△,在YNB選擇性平板上篩選出陽性克隆,隨后劃線到5-FOA平板上觀察生長狀況。
2 結果與討論
2.1 Sso1p與syntaxin1A的嵌合體不能使酵母產孢
Syntaxin含有具三個短螺旋的 N端結構域、SNARE區域及帶有極短親水尾的C端跨膜結構域[3]。膜融合過程中,syntaxin的 SNARE區域與其他SNARE蛋白結合形成四螺旋束結構,將兩側膜拉近從而進行隨后的融合。Syntaxin的SNARE區域為自身N端結構及多種其他調控蛋白的作用部位,故在本研究中考慮將 Sso1p的 SNARE區域替換成syntaxin1A的SNARE區域。
以 pRS314為載體構建含syntaxin1A(sytx1A)與Sso1p的嵌合體Sso1p/sytx1A(SNARE區域)的質粒,轉化sso1△后觀察其能否產孢。在減數分裂Ⅱ期,含不同質粒的酵母細胞均能形成四個新的細胞核;含Sso1p的釀酒酵母可形成PrM,進而形成孢子,嵌合體則只在紡錘體處形成點狀結構而無法擴張,說明其缺乏Sso1p在孢子形成過程中的功能,意味著不能將 Sso1p的 SNARE區域全部替換成sytx1A的SNARE區域,見圖1。
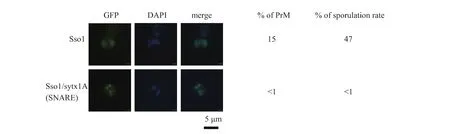
圖1 含重組質粒的sso1△在產孢培養基中培養7 h后PrM形態Fig.1 Shape of PrM of sso1△containing recombinant plasmid which cultured for 7 h in spore medium
2.2 A220為嵌合體無產孢功能的原因
Syntaxin1A與sso1p的SNARE區域有51%的同源性,將其替換sso1p的SNARE區域后卻使其失去產孢功能 (本研究中氨基酸編號為其原本在Sso1p與syntaxin1A中的編號),為何如此高的同源性功能卻有如此大的差異?故設計了一系列嵌合體對Sso1p產孢功能區進行定位,見圖2。產孢分析結果表明:保留Sso1p第214~221位氨基酸,其余SNARE區域全部替換成sytx1A,嵌合體仍能產孢,意味著 Sso1p的 214~221位氨基酸為其形成SNARE復合體所必須。
根據該結果,我們大膽假設當位于sytx1A與其他SNARE蛋白α螺旋的接觸表面上的與Sso1p有顯著性質差異的氨基酸發生突變后,可能使嵌合體具產孢功能。隨后在Sso1p/sytx1A(SNARE)基礎上設計了三個定點突變:I(209)M,HDM (213-215)TQL,AM(220,221)EE;結果sytx1A的SNARE區域發生AM(220,221)EE突變后能使sso1△產孢。進一步定點突變A(220)E、M(221)E,最終發現sytx1A發生A(220)E突變后具產孢功能,說明A220是Sso1p/sytx1A(SNARE)無產孢功能的原因。Sso1p/ sytx1A嵌合體僅將A220這一個氨基酸突變成E后便具有產孢活性,該嵌合體可用于與sytx1A相作用的蛋白質或藥物的篩選。

圖2 利用Sso1p與sytx1A的嵌合體定位Sso1p產孢功能區Fig.2 Mapping of the sporulation-specific functions in Sso1p using fusions with sytx1A
2.3 Sso1p功能的實現需要E218
由于Sso1p/sytx1A發生點突變A(220)E后具產孢功能,A220為Sso1p/sytx1A不能產孢的原因,在Sso1p中進行相應的突變E(218)A后則可以驗證Sso1p的產孢功能是否特異性需要E的存在。
將Sso1p的E218突變成A后觀察PrM狀態并進行產孢分析。結果含野生型Sso1p的菌株能形成4個PrM,突變體則只能形成點狀結構。結果表明,野生型的Sso1p發生該突變后無法形成PrM,見圖3。失去產孢功能,進一步證實該位點氨基酸E對產孢功能的重要性。
釀酒酵母中Sso1p為產孢所必須,其在營養細胞中功能與Sso2p相同[21]。在單倍體酵母中敲除SSO1和SSO2后,細胞將無法存活。于是構建了sso1△sso2△雙缺陷型單倍體菌株,可利用營養細胞實驗驗證嵌合體的功能。
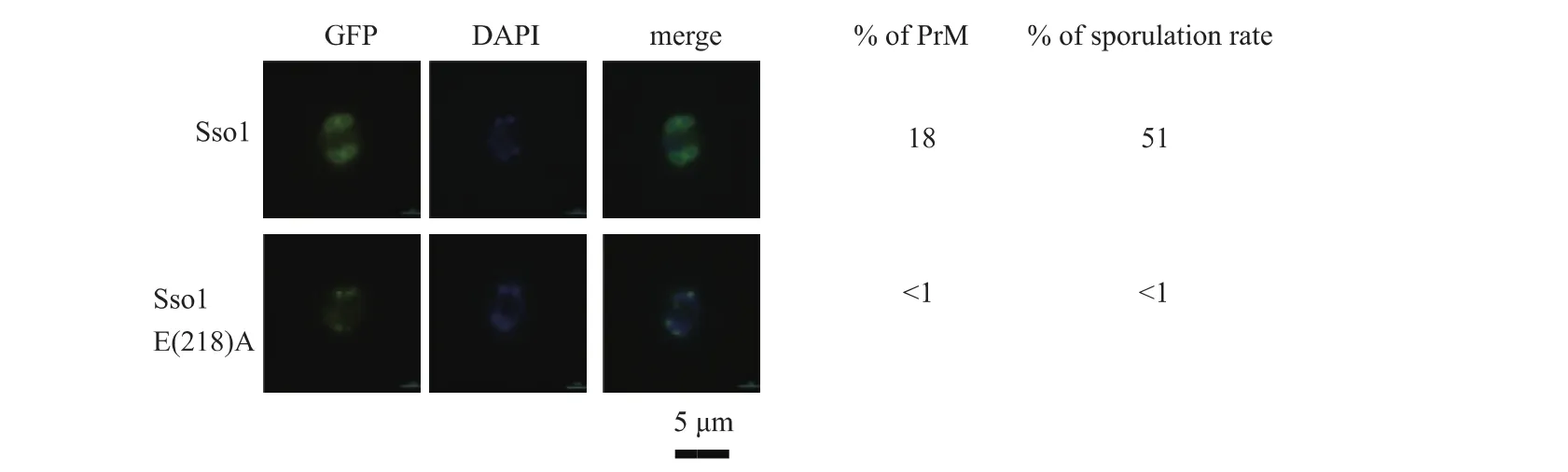
圖3 含重組質粒的sso1△在產孢培養基中培養7 h后PrM形態Fig.3 Shape of PrM of sso1△containing recombinant plasmid which cultured for 7 h in spore medium
將上述嵌合體轉入sso1△sso2△突變株,所得轉化株轉移至含5-FOA的平板上使其丟失含野生型SSO1的質粒,嵌合體將成為細胞中惟一的Sso蛋白[19];觀察轉化株生長狀況。在不含5-FOA的YNB選擇性(缺乏色氨酸、尿嘧啶)平板上,各轉化株生長狀況無明顯差別;轉接到含5-FOA的YNB選擇性(缺乏色氨酸)平板上后Sso1p/sytx1A(SNARE)-A(220)E能在 5-FOA平板上生長,Sso1p/sytx1A(SNARE)、Sso1p E(218)A則不能生長,見圖4。當Sso1p的E218突變成非極性氨基酸A后無法進行膜融合,因而不能存活。與上述結果一致,Sso1p的E218為其形成SNARE復合體所必須。利用生長分析所得結果和產孢分析一致,該系統可用于隨后的高通量篩選與sytx1A作用的基因或藥物,大大簡化篩選工作。
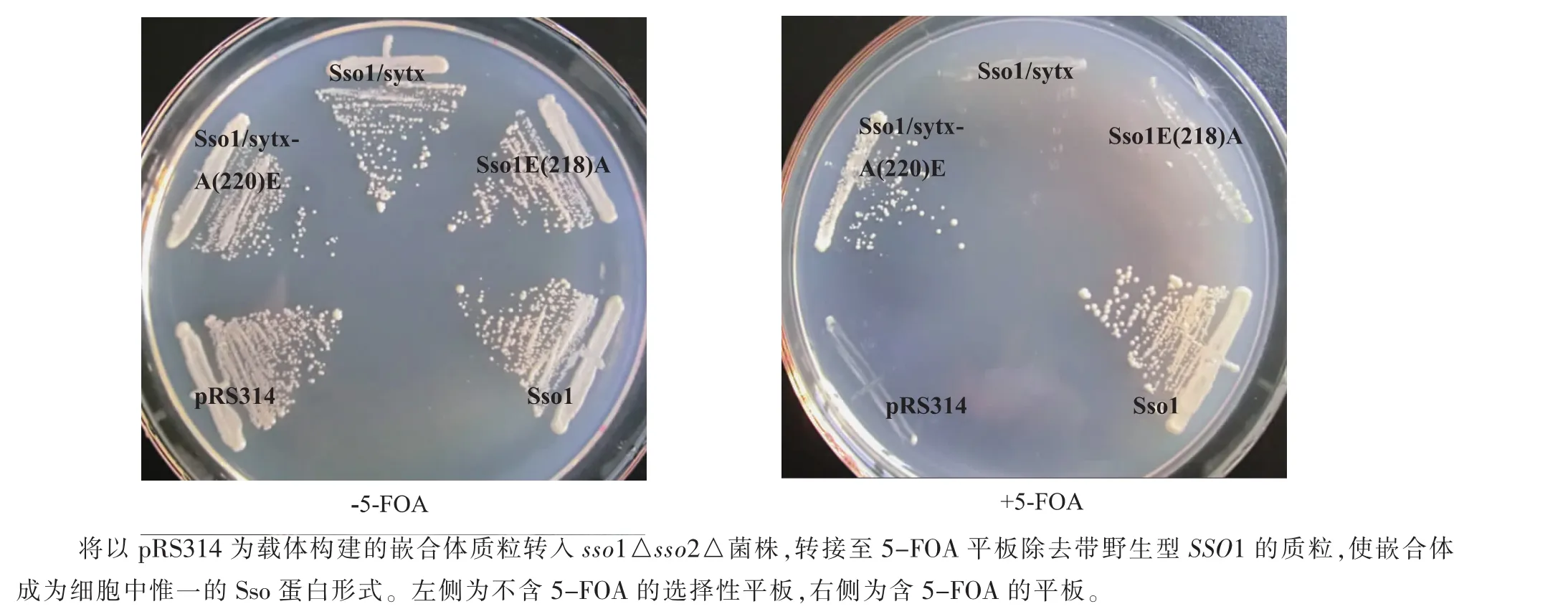
圖4 營養細胞實驗Fig.4 Vegetative cell experiments
3 結語
作者發現將酵母中Sso1p的SNARE區域替換成Sytx1A的SNARE區域而構建的嵌合體Sso1p/ sytx1A無法使釀酒酵母產生孢子,將嵌合體的A220突變成E后則具有產孢活性;而將Sso1p的E218突變成A后則失去了產孢活性,因此E218為Sso1p功能所必須。真核細胞中存在多種與syntaxin家族蛋白作用的蛋白質。Complexin與Tomosyn只存在于哺乳動物神經元細胞中,通過與syntaxin1A的SNARE區域結合調控神經遞質的釋放,具體的調控機制尚不明確。實驗構建的嵌合體Sso1p/sytx1A-A(220)E為利用酵母細胞研究這些蛋白質與sytx1A的作用機制提供了新的思路。Munc18/Sec-1也存在于釀酒酵母細胞中,與SNARE復合體結合從而調節膜融合過程。由于Munc18/Sec-1與SNARE復合體結合時,E218處于結合位點上,因此該氨基酸發生突變后可能影響這兩種蛋白質的結合。此外,形成SNARE核心復合體時,來自于4個螺旋的側鏈相接觸形成16個片層,位于片層上的氨基酸直接控制SNARE復合物的形成。而E218位于片層之外,發生點突變后使Sso1p失去活性,因此E218有可能調控SNARE復合體的形成,為釀酒酵母中膜融合機制的研究提供了參考。
[1]Hughson F M.Chaperones that SNARE neurotransmitter release[J].Science,2013,339(6118):406-407.
[2]Shi L,Shen Q-T,Kiel A,et al.SNARE proteins:one to fuse and three to keep the nascent fusion pore open[J].Science,2012,335(6074):1355-1359.
[3]Chen Y A,Scheller R H.SNARE-mediated membrane fusion[J].Nature Reviews Molecular Cell Biology,2001,2(2):98-106.
[4]Zhou P,Pang Z P,Yang X,et al.Syntaxin-1 N-peptide and Habc-domain perform distinct essential functions in synaptic vesicle fusion[J].The EMBO Journal,2013,32(1):159-171.
[5]Stein A,Weber G,Wahl M C,et al.Helical extension of the neuronal SNARE complex into the membrane[J].Nature,2009,460(7254):525-528.
[6]Südhof T C,Rothman J E.Membrane fusion:grappling with SNARE and SM proteins[J].Science,2009,323(5913):474-477.
[7]Ungermann C,Langosch D.Functions of SNAREs in intracellular membrane fusion and lipid bilayer mixing[J].Journal of Cell Science,2005,118(17):3819-3828.
[8]Garcia-Reitbock P,Anichtchik O,Bellucci A,et al.SNARE protein redistribution and synaptic failure in a transgenic mouse model of Parkinson’s disease[J].Brain,2010,133(7):2032-2044.
[9]Rendó n W O,Martí nez-Alonso E,Tomá s M,et al.Golgi fragmentation is Rab and SNARE dependent in cellular models of Parkinson’s disease[J].Histochemistry and Cell Biology,2013,139(5):671-684.
[10]Maximov A,Tang J,Yang X,et al.Complexin controls the force transfer from SNARE complexes to membranes in fusion[J]. Science,2009,323(5913):516-521.
[11]Yamamoto Y,Fujikura K,Sakaue M,et al.The tail domain of tomosyn controls membrane fusion through tomosyn displacement by VAMP2[J].Biochemical and Biophysical Research Communications,2010,399(1):24-30.
[12]Carr C M,Rizo J.At the junction of SNARE and SM protein function[J].Current Opinion in Cell Biology,2010,22(4):488-495.
[13]Rathore S S,Bend E G,Yu H,et al.Syntaxin N-terminal peptide motif is an initiation factor for the assembly of the SNARE-Sec1/Munc18 membrane fusion complex[J].Proceedings of the National Academy of Sciences,2010,107(52):22399-22406.
[14]Jantti J,Aalto M K,Oyen M,et al.Characterization of temperature-sensitive mutations in the yeast syntaxin 1 homologues Sso1p and Sso2p,and evidence of a distinct function for Sso1p in sporulation[J].Journal of Cell Science,2002,115(2):409-420.
[15]Burri L,Lithgow T.A complete set of SNAREs in yeast[J].Traffic,2004,5(1):45-52.
[16]Neiman A M.Sporulation in the budding yeast Saccharomyces cerevisiae[J].Genetics,2011,189(3):737-765.
[17]Yang H J,Nakanishi H,Liu S,et al.Binding interactions control SNARE specificity in vivo[J].The Journal of Cell Biology,2008,183(6):1089-1100.
[18]Nakanishi H,Morishita M,Schwartz C L,et al.Phospholipase D and the SNARE Sso1p are necessary for vesicle fusion during sporulation in yeast[J].Journal of Cell Science,2006,119(7):1406-1415.
[19]Boeke J D,Trueheart J,Natsoulis G,et al.5-Fluoroorotic acid as a selective agent in yeast molecular genetics[J].Methods in Enzymology,1987,154:164-175.
[20]Longtine M S,McKenzie III A,Demarini D J,et al.Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae[J].Yeast,1998,14(10):953-961.
[21]Yuan Q,Jantti J.Functional analysis of phosphorylation on Saccharomyces cerevisiae syntaxin 1 homologues Sso1p and Sso2p[J]. PloS One,2010,5(10):e13323.
Functional Analysis of the SNARE Domain of Syntaxin Family Proteins Using Yeast Sporulation
WANG Liping, SHAO Kankai, GAO Xiaodong, NAKANISHI Hideki*
(School of Biotechnology,Jiangnan University,Wuxi 214122,China)
In eukaryotic cells,most of membrane fusion processes are mediated by SNARE proteins. However,its functional and regulatory mechanisms have not been fully understood.Previous study have shown that yeast sporulation was an ideal model system to manipulate and investigate vesicle fusion machinery including SNARE proteins.The t-SNARE protein Sso1 involved in this process is an orthologue of syntaxin 1A which is required for the synaptic vesicle fusion,both share 51% homology at the SNRE domains.However,although their functional and amino acid sequence are similar,Sso1/syntaxin 1A chimera cannot complement the sporulation deficiency of sso1△mutant when SNARE domain of Sso1 is replaced completely by syntaxin 1A.To determine which residues are critical for Sso1 function,chimera and mutational analyses are further conducted.The resultsshowed that the SSO1/syntaxin 1A chimera gain the functionality by replacing Ala220 residue to Glu in the syntaxin 1A SNARE domain.By contrast,Sso1 lose its function by mutating the corresponding amino acid residue,Glu218 into Ala.Thus,Glu218 residue is specifically required for Sso1 function.
SNARE,humanization,Saccharomyces cerevisiae,Sso1p,syntaxin1A
Q 784
A
1673—1689(2015)05—0530—06
2014-00-00
國家 “111計劃”項目 (111-2-06);教育部科學研究項目 (313027);高等學校博士學科點專項科研基金項目(20120093110001);江南大學自主科研項目(JUSRP311A02)。
*通信作者:中西秀樹(1973—),男,日本人,農學博士,教授,博士研究生導師,主要從事分子生物學、酵母遺傳學、細胞生物學和糖生物學方面的研究。E-mail:hideki@jiangnan.edu.cn
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
現代出版(2020年3期)2020-06-20 07:10:34
中國科技論壇(2017年7期)2017-07-25 08:49:53
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
電測與儀表(2015年5期)2015-04-09 11:30:52
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
