戴氏綠僵菌變種拮抗耐甲氧西林金黃色葡萄球菌的作用
2013-07-06 02:03:00張孝彬陳玉燕楊艷紅
重慶理工大學學報(自然科學) 2013年4期
余 瑛,張孝彬,陳玉燕,楊艷紅
(重慶理工大學藥學與生物工程學院,重慶 400054)
蟲草是一類復型真菌,其無性型產生的次級代謝產物具有抗菌、抗病毒、抗癌、殺蟲、調節機體免疫、抗氧化[1]等作用,在醫藥領域有很大的應用潛力。戴氏綠僵菌變種Metarhizium taii var.chongqingensis nov(M.taii var)是一種低海拔地區蟲草的無性型菌株。本文報道了M.taii var發酵產物拮抗耐甲氧西林金黃色葡萄球菌(methicillin resistant Staphylococcus aureus,MRSA)的初步研究結果。
1 材料與方法
1.1 材料
1.1.1 菌種
供試菌:M.taii var,由重慶理工大學楊艷紅分離保存。
病原菌:MRSA標準菌株WHO2、臨床分離的金黃色葡萄球菌(Staphylococcus aureus,SA)共10株,由第三軍醫大學西南醫院惠贈。
1.1.2 培養基
1)PDA固體培養基:馬鈴薯200 g,葡萄糖20 g,瓊脂 15 g,pH=7.0,去離子水定容至 1 000 mL。
2)細菌培養基:胰蛋白10 g,酵母粉5 g,氯化鈉 5 g,Ph=7.0,去離子水定容至 1 000 mL。
3)抑菌平板培養基:胰蛋白10 g,酵母粉5 g、氯化鈉5 g,瓊脂30 g,去離子水定容至1 000 mL。
4)生長發酵培養基:NaNO30.5 g,KCl 0.5 g,蔗糖 30 g,蠶蛹粉 0.5 g,pH=6.5,去離子水定容至1 000 mL。
5)發酵培養基:葡萄糖10 g,蛋白胨2.5 g,酵母粉 5 g,pH 值為 6.0 ~7.0,去離子水定容至1 000 mL。
1.1.3 主要試劑
蛋白酶K,購自鼎國生物技術有限公司。
1.2 方法
1.2.1 供試菌種的活化
取-80℃冰箱保存的供試菌 M.taii var,在PDA平板上劃線接種,于28℃培養14 d至孢子發育成熟。
1.2.2 供試菌的發酵培養
孢子懸液的制備:從PDA平板上刮取孢子,用無菌水重懸成孢子懸液后以4層無菌擦鏡紙過濾孢子液,血球計數板計數孢子,并調整孢子懸液濃度為109cell/mL。
接種及發酵:取100 μL孢子懸液接種到100 mL生長培養基中。培養2 d后,按1∶10接種至發酵培養基中。發酵培養條件優化應用正交設計法,采用2組變量:發酵培養基 pH 值為5.5、6.0、6.5、7.0;培養溫度為 25℃、28℃、30℃。發酵培養3~9 d,每日取發酵上清2 mL,10 000 r/min離心5 min,取上清進行抗WHO2的活性測定。
1.2.3 病原菌的培養
挑取活化的WHO2或各臨床分離的SA單克隆分別接種于 100 mL細菌培養基,37℃,250 r/min過夜培養,備用。
1.2.4 抑菌活性測定
M.taii var發酵上清拮抗WHO2的活性測定采用瓊脂擴散法[2]。將過夜培養的 WHO2,用血球計數板計數調整濃度為104個/mL,用移液槍吸取100 μL菌液,均勻涂布到抑菌平板培養基上。用打孔器在培養基上打孔,孔中加入發酵上清液150 μL,于37℃恒溫培養箱中放置培養12 h,測定抑菌圈直徑大小。
臨床分離的SA對甲氧西林敏感性測定結果由西南醫院提供。SA對M.taii var發酵上清的敏感性采用瓊脂擴散法測定。
1.2.5 高溫及蛋白酶K對M.taii var發酵上清活性的影響
將發酵液上清分別在45℃、65℃、85℃、100℃各處理5 min、10 min、15 min,測定抑菌活性。用未經高溫處理的發酵液上清作對照。
分別在100 μL發酵液上清中加入質量體積分數為10 mg/mL蛋白酶 K 50μL,37℃各孵育30 min、60 min,測定抑菌活性。用未加入蛋白酶 K發酵液上清作對照。
2 結果與分析
2.1 發酵溫度及pH對M.taii var發酵上清抗菌活性成分表達的影響
在前期已對培養基進行篩選的基礎上,采用不同pH的發酵培養基和發酵溫度進行培養,提取發酵上清測定抑菌活性,結果(見表1)表明:30℃,pH值為5.5~6.0條件最適合抑菌成分的表達,該條件下發酵上清的抑菌活性最先達到峰值,即發酵7 d時抑菌圈直徑達到3.6 cm,其活性高于其他條件下的發酵上清。
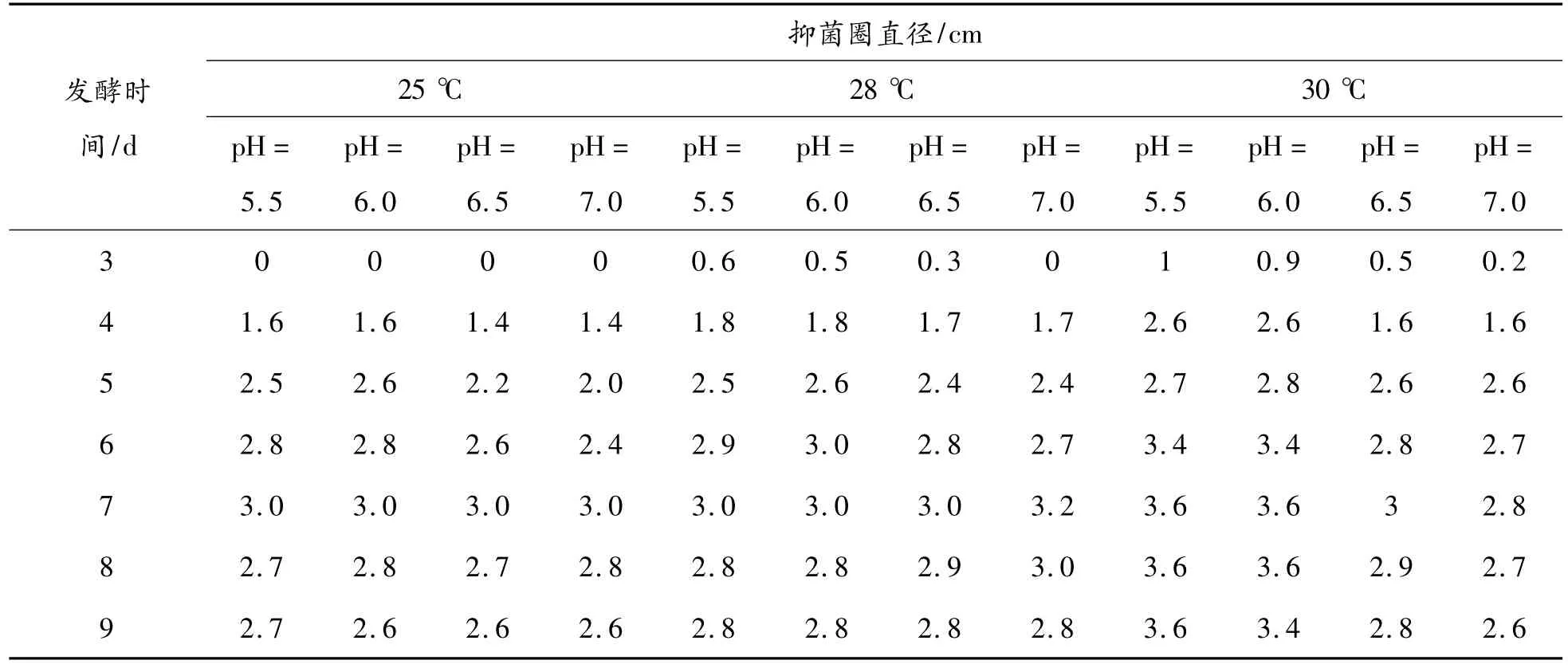
表1 不同培養溫度、pH條件下M.taii var發酵上清拮抗MRSA的活性
2.2 高溫對M.taii var發酵上清抗菌活性的影響
將M.taii var發酵上清分別用45℃、65℃、85℃、100℃處理后,測定其抑菌活性變化,結果顯示:不同溫度處理后 M.taii var發酵上清拮抗WHO2的抑菌圈直徑無顯著變化(見圖1),即M.taii var發酵上清中的抑菌成分在高溫條件下活性穩定。
2.3 蛋白酶K對M.taii var發酵上清抗菌活性的影響
在M.taii var發酵上清中加入蛋白酶K,37℃孵育30 min或60 min后測定其抑菌活性變化,結果顯示:處理后的M.taii var發酵上清拮抗WHO2的抑菌圈大小無顯著變化(見圖2),即蛋白酶K對M.taii var發酵上清中的抑菌成分活性無影響。
2.4 M.taii var發酵上清對各臨床分離SA的拮抗作用
測定M.taii var發酵上清對各臨床分離SA的抗菌活性,結果顯示該發酵上清對臨床分離的SA菌株均有拮抗作用,其抑菌圈直徑在3.4~3.6 cm(表2未顯示),這些被檢測的SA中有對甲氧西林耐受菌株即MRSA菌株,也有對甲氧西林敏感的SA菌株(見表2)。
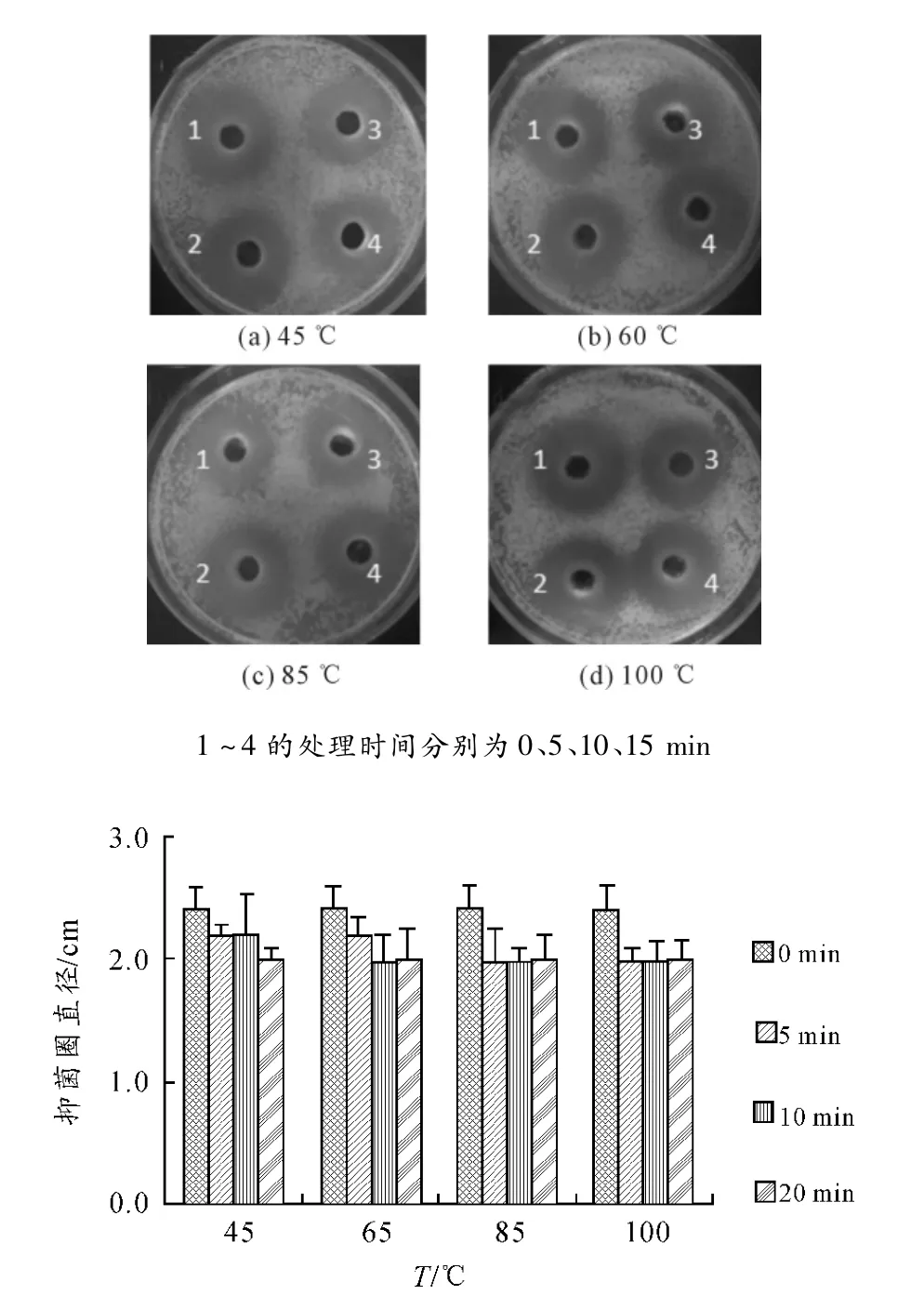
圖1 M.taii var抑菌成分在高溫處理后的穩定性
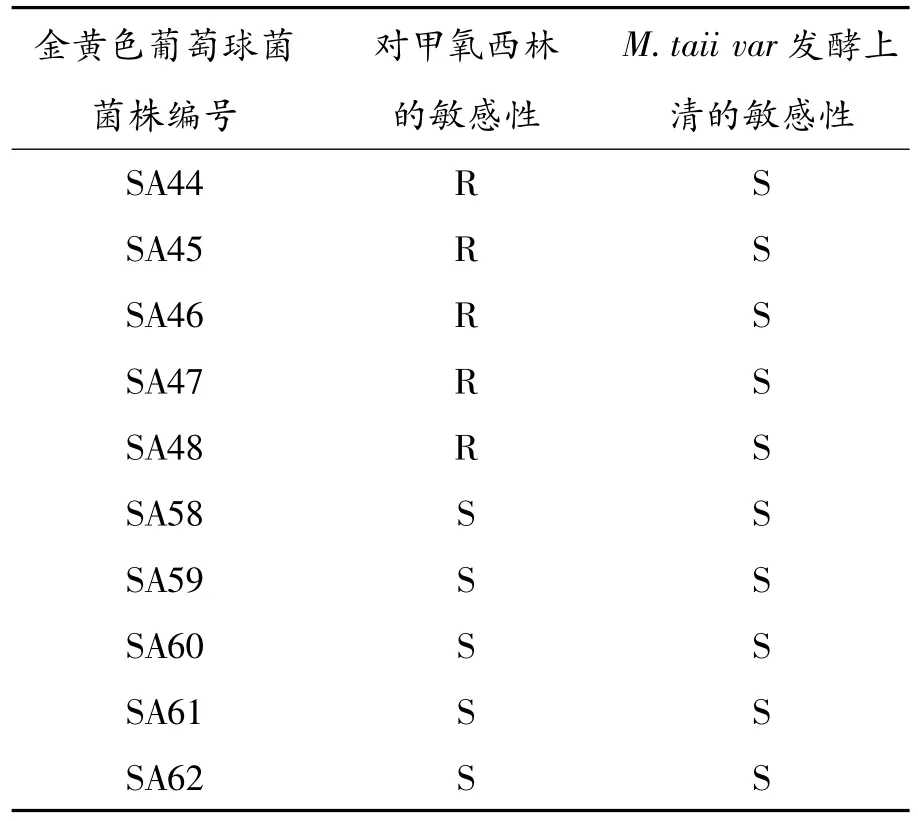
表2 M.taii var發酵上清對臨床分離SA的抗菌活性
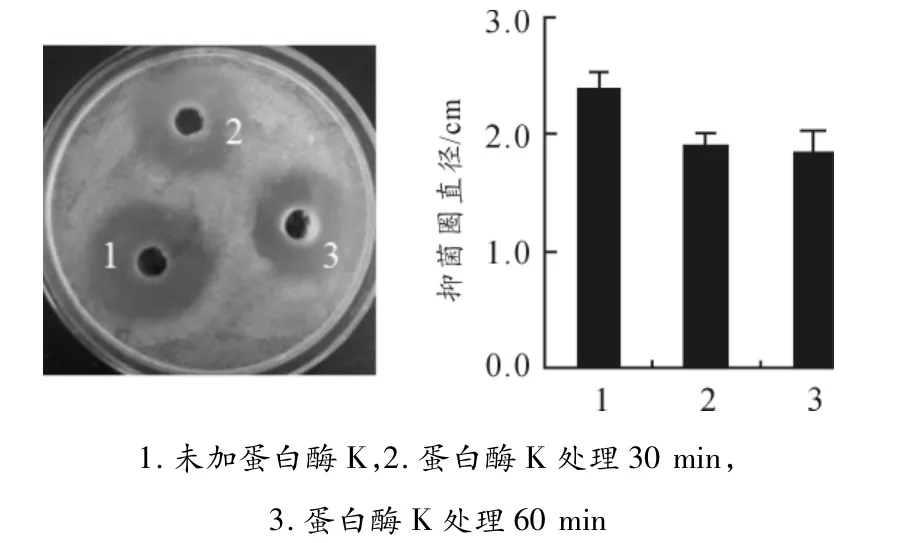
圖2 M.taii var發酵上清抑菌成分經蛋白酶K消化后的活性
3 討論
SA是引起細菌性食物中毒和化膿感染最常見的病原菌之一。自19世紀40年代青霉素問世以來,SA引起的感染性疾病受到較大的控制。但隨著青霉素的廣泛使用,有些SA產生了一種能水解β-內酰胺環的酶,可使青霉素無效,即產生了耐藥性。隨后推出的甲氧西林在臨床使用2年后就出現了耐甲氧西林的菌株MRSA[3]。目前,MRSA已成為院內感染的重要病原菌之一。耐藥菌株的不斷出現,使得MRSA治療藥物變得日益匱乏,尋找新的抗MRSA活性物質來源是進行新型抗MRSA藥物研發的工作基礎。本研究研究證實M.taii var發酵上清對臨床分離的不同MRSA菌株及普通SA菌株都有抗菌活性,表明M.taii var產生的抗菌成分與甲氧西林有不同的抗菌機制,該活性物該抗菌成分還有耐高溫和抗蛋白酶K水解的特性,在pH值為5.6~6.0,溫度為30℃時表達活性最高。已有的研究結果顯示綠僵菌、白僵菌等真菌的次生代謝產物中含有一種結構為環肽的綠僵菌素(destruxins,Dtxs)[4-5],具有抗菌、抗蟲、抗腫瘤、抗炎癥、抗骨質疏松、免疫抑制等多種活性[6],Dtxs結構均由一個α-羥酸和5個氨基酸殘基構成,不同菌株來源的Dtxs的α-羥酸類型、N-甲基化修飾、氨基酸側鏈基團有所不同,均有耐高溫、抗蛋白酶水解的特性[7]。M.taii var抗MRSA是否含有與先前報道的DTXs類似的結構仍需進一步研究。
[1]焦彥朝,梁宗琦,劉愛英.蟲草生物活性物質研究概況[J].貴州農業科學,1990(3):53-58.
[2]周長林.微生物學實驗與指導[M].北京:中國醫藥科技出版社,2008.
[3]Jensen J U S,Jensen E T,Larsen A R,et al.Control of a methicillin-resistant Staphylococcus aureus(MRSA)outbreak in a day-care institution[J].Journal of Hospital Infection,2006,63(1):84-92.
[4]Peng K C,Huang H S,Teng Y M,et al.Circular dichroism analysis of destruxins from Metarhizium anisopliae O-riginal Research Article[J].Journal of Biochemical and Biophysical Methods,2005,62(1):41-50.
[5]Morais-Urano R P,Chagas A C S,Berlinck R G S,et al.Acaricidal action of destruxins produced by a marine-derived Beauveria felina on the bovine tick Rhipicephalus(Boophilus)microplus[J].Experimental Parasitology,2012,132(3):362-366.
[6]Sarabia F,Chammaa S,Sánchez-Ruiz A,et al.Chemistry and biology of cyclic depsipeptides of medicinal and biological interest[J].Curr Med Chem,2004(11):1309.
[7]Asai T,Yamamoto T,Chung Y M,et al.Aromatic polyketide glycosides from an entomopathogenic fungus[J].Cordyceps indigotica Tetrahedron Letters,2012,53:277-280.
