KDR等6個基因多態性與雞生長和體組成性狀的相關研究
2012-08-08 12:23:30吳憲文金鵬程王守志
東北農業大學學報 2012年12期
吳憲文,金鵬程,王守志,張 慧,李 輝
(農業部雞遺傳育種重點實驗室,東北農業大學動物科技學院,哈爾濱 150030)
試驗前期通過雞腹脂性狀的全基因組關聯分析(GWAS)鑒定出多個與雞腹脂性狀顯著相關的多態性位點,其中6個位點分別為rs14483313(命名來源于60 K SNP芯片,下同)位于Ⅲ型酪氨酸激酶受體(a type III receptor tyrosine kinase,KDR/FlkⅠ/VEGF-2)上;rs13548811位于雞腫瘤候選抑癌基因3(Tumor suppressor candidate 3,TUSC3)上 ;GGaluGA262999位于磷酸核糖焦磷酸轉氨酶(Phosphoribosyl pyrophosphate amidotransferase,PPAT)上;GGaluGA263078位于胞外復合體成分1(Exocyst complex component 1,EXOC1)上;GGaluGA176566位于v-myb鳥類成髓細胞血癥病毒致癌基因同系物2(v-myb myeloblastosis viral oncogene homolog(avian)-like 2,MYBL2)上;rs14482748位于 KIAA 1211上;這些位點的多態性與肉雞高、低腹脂雙向選擇品系第11世代的腹脂重和腹脂率顯著相關。
本研究對6個多態位點在高、低脂肉雞雙向選擇品系第14世代群體和AA隨機群體中進行SNPs檢測和關聯分析,旨在研究這6個基因的多態位點與雞生長和體組成性狀的相關性,從而為后續開展相關基因的功能研究和尋找可用于肉雞育種的分子標記奠定基礎。
1 材料與方法
1.1 雞群和性狀的測定
以東北農業大學動物科技學院選育的高、低脂肉雞雙向選擇品系第14世代群體(NEAUHLFG14)612個體(高脂系320個體,低脂系292個體)和AA肉雞商業隨機群體386個體為試驗材料。試驗雞群按常規方法進行飼養管理,7周齡時翅靜脈采血,EDTA-Na2抗凝,酚∶氯仿抽提DNA之后,TE溶解,-20℃保存。測定1、3、5、7周齡體重(Body weight,BW)、屠體重(Carcass weight,CW)、腹脂重(Abdominal fat weight,AFW)、腹脂率(Abdominal fat percent,AFP)(腹脂重/體重)、肝臟重(Liver weight,LW)、心臟重(Heart weight,HW)、脾臟重(Spleen weight,SpW)、睪丸重(Testicle weight,TeW)、極低密度脂蛋白濃度(Very low density lipoproteins,VLDL)。
1.2 引物設計
結果見圖1。

表1 引物序列Table 1 Primer sequences
根據本實驗室雞腹脂性狀的全基因組關聯分析提供的多態性位點數據和雞基因組測序結果(www.genome.ucsc.edu),使用Primer 5.0軟件設計6對引物用于多態性檢測,引物由北京英駿生物公司合成,如表1所示。
1.3 基因分型
根據六個多態位點,采用PCR-RFLP方法對高、低脂肉雞雙向選擇品系第14世代群體和AA肉雞商業隨機群體個體進行基因型分析。PCR反應體系(25 μL):50 ng·μg-1基因組DNA 1 μL、10×PCR Buffer 2.5 μL、10 mmol·L-1dNTP 2 μL、10 μmol·L-1上下游引物各0.5 μL、5 U·μL-1TaqDNA聚合酶0.25 μL、去離子水18.25 μL。PCR 反應條件:預變性條件均為94℃,5 min;變性條件均為94℃,30 s;延伸條件均為72℃,20 s;終極延伸條件均為72℃,5 min。退火條件、循環數及酶切條件如表2所示。內切酶由NEB公司提供,酶切反應體系(20 μL),按照使用說明進行配置。
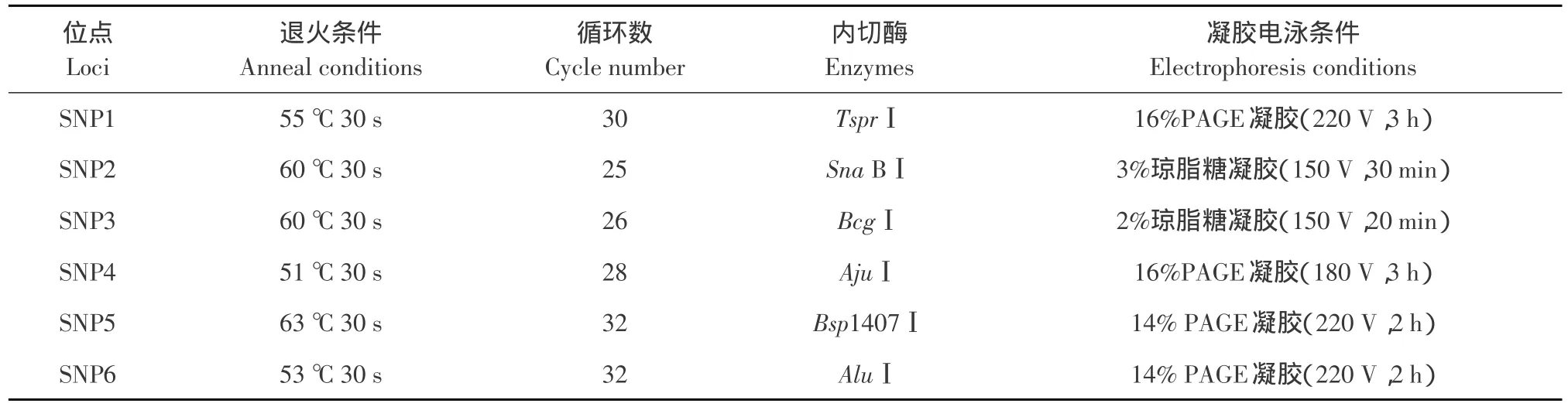
表2 各位點PCR及酶切條件Table 2 Conditions of PCR,PCR-RFLP
1.4 統計分析
根據高、低肉雞腹脂雙向選擇品系群體的特點,構建基因型效應統計模型①,根據AA肉雞商業隨機群體的特點,構建基因型效應統計模型②:

其中Y為性狀觀測值,μ為群體均值,G為基因型固定效應,L為品系固定效應,G*L為基因型和品系互作效應,F(L)為品系內的家系效應,D(F,L)為家系內與品系內母雞的隨機效應,S為性別固定效應,G*S為基因型和性別互作效應,F為家系的隨機效應,D(F)為家系內母雞的隨機效應,BW7為7周齡體重做為屠體組成性狀的校正因子,e為剩余值效應。
運用統計軟件JMP4.0(SAS公司,2000)進行相關分析;采用SAS9.1.3軟件對高、低肉雞腹脂雙向選擇品系間基因型頻率和等位基因頻率差異進行χ2獨立性檢驗。
2 結果與分析
2.1 PCR-RFLP檢測
結果見圖1。

圖1 PCR-RFLP基因分型Fig.1 PCR-RFLP patterns
針對以上六個位點用相應的引物對高、低肉雞腹脂雙向選擇第14世代群體和AA肉雞商業隨機群體基因組DNA進行PCR擴增,擴增片段與目的片段大小一致且特異性較好。采用RFLP的方法進行個體基因型分析。PCR產物經過酶切后均產生3種基因型,分型及命名見圖1。圖1中A-E分別對應為SNP1-6的基因分型圖片。
2.2 群體遺傳學分析
對六個突變位點在NEAUHLF第14世代中進行群體遺傳學分析,檢驗結果表明基因型頻率、等位基因頻率分布在高、低腹脂雙向選擇品系間均存在極顯著差異(P<0.01)。SNP1和SNP3位點TT基因型和T等位基因在低脂系中的頻率極顯著高于在高脂系中的頻率,CC基因型和C等位基因在高脂系中的頻率極顯著高于在低脂系中的頻率。SNP2、SNP4和SNP5位點TT基因型和T等位基因在高脂系中的頻率極顯著高于在低脂系中的頻率,CC基因型和C等位基因在低脂系中的頻率極顯著高于在高脂系中的頻率。SNP6位點AA基因型和A等位基因在高脂系中的頻率極顯著高于在低脂系中的頻率,GG基因型和G等位基因在低脂系中的頻率極顯著高于在高脂系中的頻率,如表3所示。
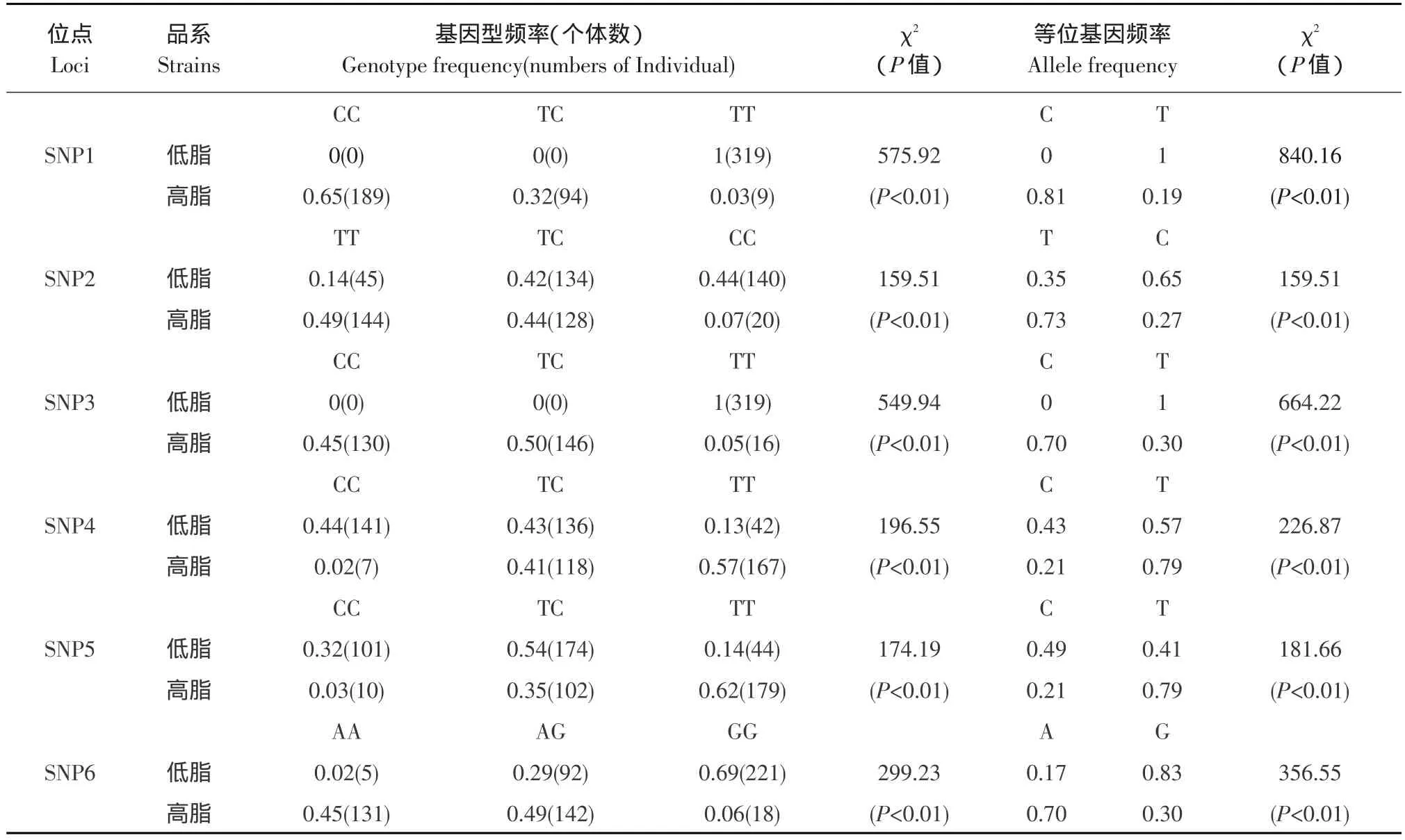
表3 六個位點不同基因型、等位基因頻率在高、低脂系兩系間的分析Table 3 Comparisons of frequencies of genotype and allele in two lines of chicken divergently selected for abdominal fat content
2.3 6個位點多態性與雞生長和體組成性狀的相關分析
在高、低肉雞腹脂雙向選擇品系第14世代群體和AA肉雞商業隨機群體中進行基因型與生長和體組成性狀的相關分析,各位點對性狀的影響(P值)見表4,基因型效應間的多重比較結果見表5。結果表明SNP1位點的多態性對NEAUHLFG14的AFW、AFP、CW和LW 有顯著影響(P<0.05),并且這四種性狀的CC和CT基因型個體最小二乘均值(以下簡稱為均值)均顯著高于TT基因型個體(P<0.05)。
SNP2位點的多態性對NEAUHLFG14的BW7、HW和VLDL有顯著影響(P<0.05),BW7和VLDL的CC和TC基因型個體均值顯著高于TT型個體(P<0.05),HW的TT和TC型個體均值顯著高于TT型個體(P<0.05);該位點的多態性對AA群體的SpW有顯著影響(P<0.05),且CC型個體均值顯著高于TC和TT型個體(P<0.05)。
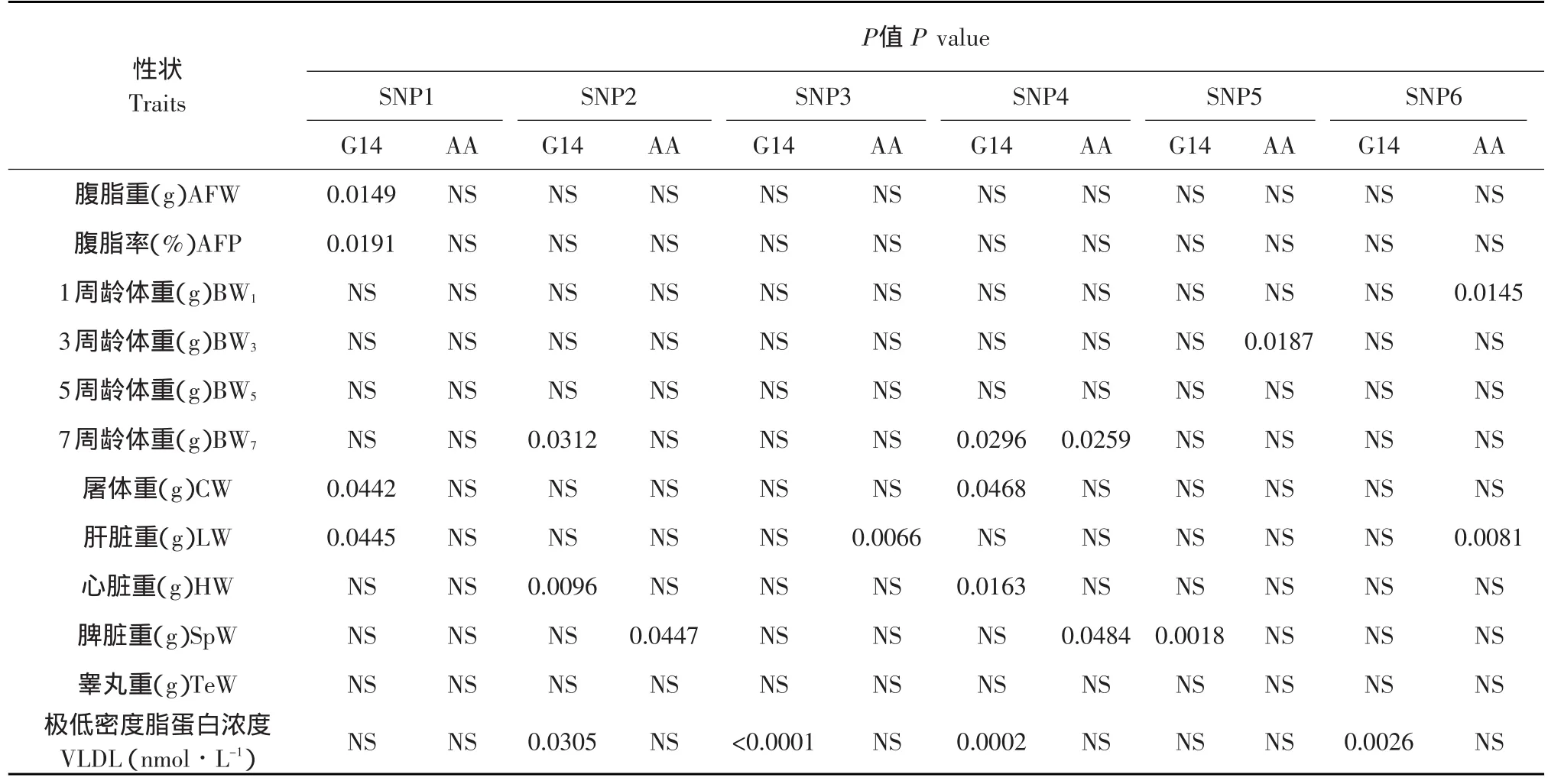
表4 六個多態位點對高、低脂雙向選擇品系14世代和AA肉雞商業隨機群體生長和體組成性狀的影響Table 4 Effects of six polymorphism on growth and body composition traits in NEAUHLFG14 and Arbor Acres(AA)commercial random population
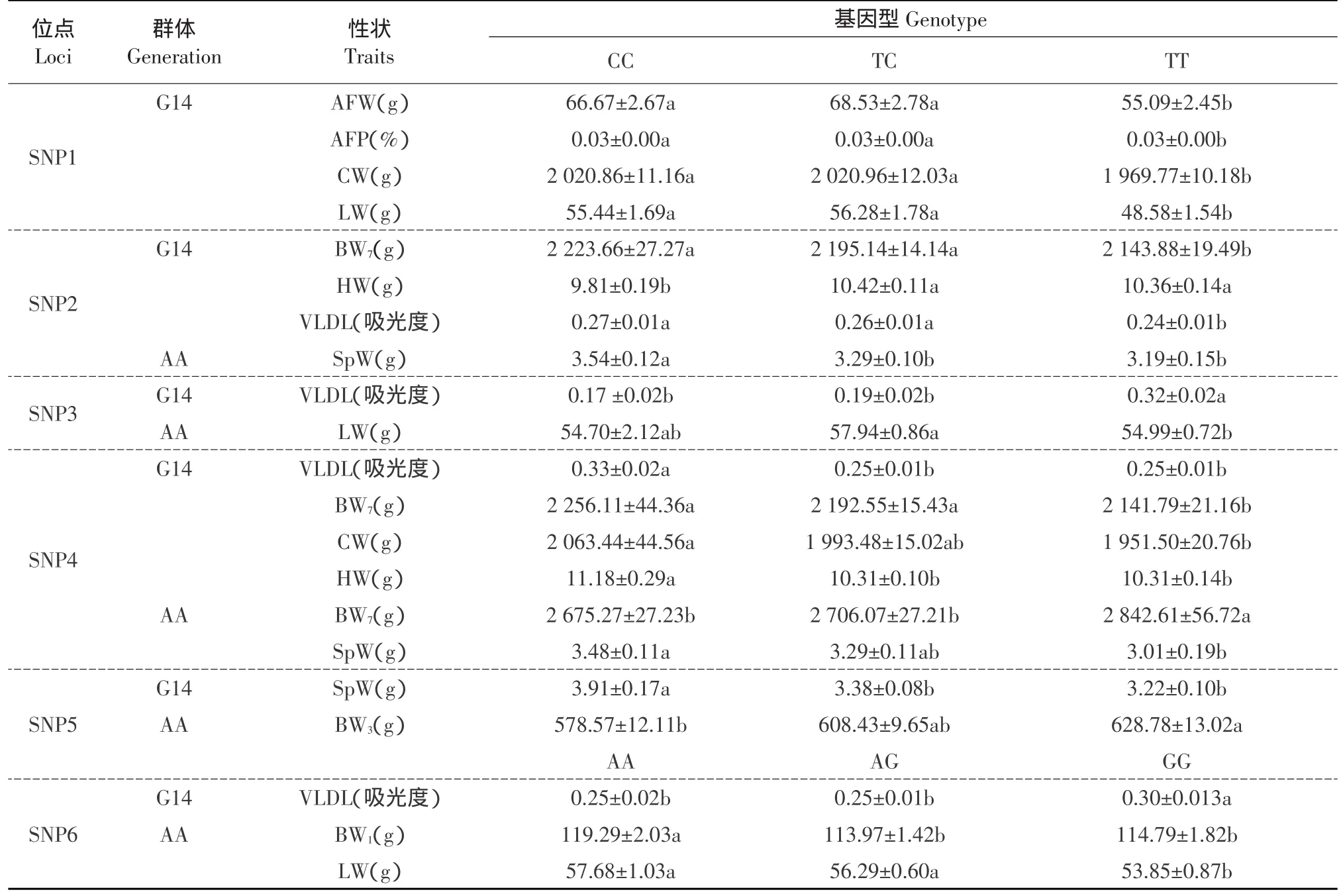
表5 高、低脂雙向選擇品系第14世代中各位點基因型間多重比較Table 5 Multiple comparisons among least-squares means with different genotypes in NEAUHLFG14
SNP3位點的多態性對NEAUHLFG14的VLDL有顯著影響(P<0.05),且TT型個體均值顯著高于TC和CC型個體(P<0.05);該位點的多態性對AA群體的LW有顯著影響(P<0.05),且TC型個體均值顯著高于TT型個體(P<0.05)。
SNP4位點多態性對NEAUHLFG14的BW7、CW、HW和VLDL有顯著影響(P<0.05),CC型個體的VLDL和HW顯著高于TC和TT型個體(P<0.05),CC和TC基因型個體的BW7顯著高于TT型個體(P<0.05),CC型個體的CW顯著高于TT型個體(P<0.05);該位點的多態性對AA群體的BW7和SpW有顯著影響(P<0.05),且TT型個體的BW7顯著高于TC和CC型個體(P<0.05),CC型個體的SpW顯著高于TT型個體(P<0.05)。
SNP5位點多態性對NEAUHLFG14的SpW有顯著影響(P<0.05),且CC型個體均值顯著高于TC和TT型個體(P<0.05);該位點的多態性對AA群體的BW3有顯著影響(P<0.05),TT型個體均值顯著高于CC型個體(P<0.05)。
SNP6位點多態性對NEAUHLFG14的VLDL有顯著影響(P<0.05),且GG型個體均值顯著高于AA和AG型個體(P<0.05);該位點的多態性對AA群體的BW1和LW有顯著影響(P<0.05),且AA個體BW1顯著高于AG和GG型個體(P<0.05),AA和AG型個體的LW顯著高于GG型個體(P<0.05)。
3 討論與結論
rs14483313突變位于KDR基因上,KDR基因也叫做FlkⅠ、VEGF-2,血管內皮生長因子2(Vascular endothelial growth factor-2,VEGF-2)是血管內皮生長因子家族成員[1],它與血管的生成及造血功能有關[2]。有研究表明血管生成與脂肪生成在時間和空間上有很大的聯系。在時間上關于脂肪組織發育研究指出,胚胎期血管的生成早于脂肪的生成[3]。在空間上新的血管生成總是伴隨著脂肪細胞的分化,在這個過程中VEGF是關鍵因子[4]。肥胖與骨骼肌毛細血管密度降低有關[5],抑制血管生成可以減少脂肪聚集[6-9]。可以看出血管生成與脂肪生成之間存在密切關系。
本研究結果顯示,rs14483313(SNP1)突變對AA肉雞商業隨機群體腹脂性狀的影響未達到顯著水平。但對高、低脂肉雞雙向選擇品系第14世代AFW和AFP都有顯著影響(P<0.05),這與前期GWAS在高、低脂肉雞雙向選擇品系第11世代所得結果一致。該位點在NEAUHLFG14的低脂系中所有個體均為TT基因型,推測T等位基因有可能經過前幾個世代的選擇已固定。從基因型效應來看,TT基因型個體AFW、AFP顯著低于CC型個體(P<0.01),所以,T等位基因是降低雞腹脂沉積的等位基因,在育種中可以嘗試選擇TT基因型個體培育低脂肉雞品系。
研究發現,rs13548811(SNP2)突變對高、低脂肉雞雙向選擇品系第14世代群體后期體重性狀有影響,從3~7周齡逐漸增大,到3、5周齡時已接近顯著(P值分別為0.0736,0.0637,表4中未給出),7周齡時達到顯著水平(P<0.05)。CC型個體的7周齡體重顯著高于TT型個體(P<0.05),CC基因型和C等位基因在低脂系中的頻率顯著高于在高脂系中的頻率,以此推斷CC基因型為高、低脂系中體重性狀的有利基因型,可以嘗試在育種中通過選擇此基因型個體提高雞的體重性狀。
GGaluGA263078(SNP4)突變對雞7周齡體重有顯著影響,尤其在NEAUHLFG14和AA群體遺傳背景不同的群體中該位點對BW7的影響一致(P<0.05),因此推測該位點很可能是影響雞體重性狀的致因突變位點,或與致因突變位點緊密連鎖。
本文所研究的6個突變位點在高、低脂肉雞雙向選擇品系第11世代中都與腹脂性狀顯著相關,但在NEAUHLFG14和AA群體中卻無一致結果。群體的遺傳背景不同是造成群體間相關分析不一致的重要原因之一。高、低脂雙向選擇品系是一個對腹脂性狀經過多世代選擇的群體,隨著從第11世代到第14世代的選擇,該群體基因組位點間連鎖不平衡程度發生了變化;與高、低脂雙向選擇品系群體相比,AA隨機群體未經過特定性狀選擇群體,其遺傳背景(基因的等位基因頻率、基因位點連鎖不平衡程度等)不同的。這時突變位點與影響雞腹脂性狀的致因突變緊密連鎖。
在這6個突變位點中有4個(SNP2、SNP3、SNP4和SNP6)都與NEAUHLFG14的VLDL濃度顯著相關(P<0.05)。有研究表明,VLDL濃度是間接度量雞體肥度的最好指標,血漿VLDL濃度與腹脂重和腹脂率呈顯著的正表型相關,相關系數為0.4168和0.3576[10],對血漿VLDL濃度的低向選擇會使腹脂沉積減少[11]。結果表明,這些基因可能對雞的腹部脂肪組織生長和發育發揮重要作用。
綜上所述,這六個基因可能是影響雞腹脂性狀的重要基因,但本研究中的多態位點是否為功能性位點需進一步驗證。
[1] Hu J S,Hastings G A,Cherry S,et al.A novel regulatory function of proteolytically cleaved VEGF-2 for vascular endothelial and smooth muscle cells[J].FASEB J,1997,11(6):498-504.
[2] Shalaby F,Ho J,Stanford W L,et al.A requirement for Flk1 in primitive and definitive hematopoiesis and vasculogenesis[J].Cell,1997,89(6):981-990.
[3] Neels J G,Thinnes T,Loskutoff D J.Angiogenesis in anin vivomodel of adipose tissue development[J].FASEB J,2004,18(9):983-985.
[4] Nishimura S,Manabe I,Nagasaki M,et al.Adipogenesis in obesity requires close interplay between differentiating adipocytes,stromal cells,and blood vessels[J].Diabetes,2007,56(6):1517-1526.
[5] Gavin T P,Stallings H R,Zwetsloot K A,et al.Lower capillary density but no difference in VEGF expression inobesevs.lean young skeletal muscle in humans[J].J Appl Physiol,2005,98(1):315-321.
[6] Rupnick M A,Panigrahy D,Zhang C Y,et al.Adipose tissue mass can be regulated through the vasculature[J].Proc Natl Acad Sci,2002,99(16):10730-10735.
[7] Liu L,M M.Angiogenesis Inhibitors may regulate adiposity[J].Nutrition Reviews,2003,61(11):384.
[8] Fukumura D,Ushiyama A,Duda D G,et al.Paracrine regulation of angiogenesis and adipocyte differentiation duringin vivoadipogenesis[J].Circ Res,2003,93(9):88-97.
[9] Kolonin M G,Saha P K,Chan L,et al.Reversal of obesity by targeted ablation of adipose tissue[J].Nat Med,2004,10(6):625-632.
[10] 李輝,龔道清.肉雞血漿極低密度脂蛋白濃度與屠體肥度性狀的相關研究[J].黑龍江畜牧獸醫,1997(8):1-5.
[11] 李輝,王靜.蛋用型雞血漿極低密度脂蛋白濃度與蛋黃脂肪酸含量的相關研究[J].新疆農業科技,1996(1):54-57.
