斑點叉尾鮰hepcidin抗菌肽在大腸桿菌中的融合表達
2012-08-08 12:23:42趙冬梅
東北農(nóng)業(yè)大學學報 2012年12期
關鍵詞:融合
趙冬梅,陶 妍
(上海海洋大學食品學院,上海 201306)
Boman首先從惜古比天蠶蛹(Hyalophora cecropia)的免疫血淋巴細胞中分離到高效抗菌肽cecropin以來[1],至今已經(jīng)有約2 000多種抗菌肽被分離,主要來自于兩棲動物、甲殼動物、鳥類、魚類、哺乳動物和人類[2]。抗菌肽一般為富含帶正電荷氨基酸的堿性小分子,氨基端親水、羧基端疏水,具有兩性性質及廣譜抗菌性[3],被認為是抗生素的最佳替代品。此外,部分抗菌肽對胰蛋白酶或胃蛋白酶穩(wěn)定,并對某些真菌、原蟲、病毒和癌細胞具有殺傷作用[4]。
Hepcidin又稱LEAP-1(Liver-expressed antimicrobial peptide 1),是一種主要由肝細胞產(chǎn)生和分泌的小分子陽離子肽,具有較廣譜抗菌活性。它是由Kruase等[5]于2000年首次從人血清中分離出,次年又由Park等[6]從人尿液中分離,以后從哺乳動物、兩棲類動物和魚類中亦發(fā)現(xiàn)類似于hepcidin的同源基因[7]。hepcidin的成熟肽存在三種形式,分別為25個氨基酸(Hepc 25)、22個氨基酸(Hepc 22)和20個氨基酸(Hepc 20)的小肽,Hepc 25是主要存在形式。機體細胞中的hepcidin含量甚微,提取純化困難,而化學合成成本高,根本無法大量獲得。因此,基于重組DNA表達的基因工程技術無疑是制備hepcidin抗菌肽的最佳方法[8-10]。
近年來關于抗菌肽重組DNA表達方面的研究報道不少,但主要集中于哺乳類、昆蟲類和兩棲動物等[7]。相比之下,關于魚類抗菌肽基因克隆方面的研究報道較少,Bao等通過對斑點叉尾鮰(Ictalurus punctatus)進行細菌感染試驗,從其肝臟中分離和克隆了hepcidin的全長cDNA,并考察了該基因在不同組織中的表達水平[11]。重組DNA技術在魚類抗菌肽制備中的應用則甚少。據(jù)此,本文以斑點叉尾鮰肝臟組織為材料,以成熟肽為25個氨基酸的hepcidin為研究對象,通過RT-PCR對編碼其原前體肽(包括前體肽和成熟肽)的cDNA進行克隆;選擇pET32a(+)為表達質粒、E.coliBL21(DE3)為工程菌,構建原核表達系統(tǒng),在IPTG誘導下,實現(xiàn)hepcidin原前體肽在大腸桿菌中的融合表達。
1 材料與方法
1.1 材料
斑點叉尾鮰(體重800 g,體長35 cm)1尾購自上海市浦東果園農(nóng)貿(mào)市場。鮮活魚運至實驗室后,立即用鈍器擊殺取其肝臟分割成0.5 cm見方小塊,保存于-80℃冰箱待用。
1.2 總RNA提取和第一股cDNA合成
采用 Trizol試劑(Invitrogen,Carlsbad,CA,USA),按照說明書對斑點叉尾鮰的肝臟組織進行總RNA提取(100 mg∶1 mL),經(jīng)75%乙醇純化后的總RNA溶解于10 μL DEPC·H2O中。
第一股cDNA的合成根據(jù)M-MLV RTase cDNA Synthesis Kit(Takara,Otsu,Japan)說明書進行。2 μL總RNA、5 μL AP(Adapter Primer)通用引物、2 μL ddH2O混勻,70℃保溫10 min,冰浴1~2 min;依次 加 入 4 μL 5 × First strand buffer、 4 μL dNTP mixture(2.5 mmol·L-1)、 2 μL DTT(0.1 mol·L-1),42℃保溫2 min,再加入1 μL Reverse Transcriptase M-MLV(RNase H-)(200 U·L-1)保溫 70 min;70 ℃保溫15 min、冰上冷卻后得到第一股cDNA。
1.3 Hepcidin原前體肽基因的PCR擴增和測序
參考斑點叉尾鮰hepcidin的全長cDNA序列(GenBank:AY834209)設計一對引物,正向引物和反向引物分別為H-F(5'ATGAGGGCAATGAGC ATCGCG 3')和H-R(5'TTAGAACCTGCAGCAGAA CCC 3'),用于擴增hepcidin全長cDNA。以斑點叉尾鮰肝臟的第一股cDNA為模板,PCR反應體系和條件如下:2.5 μL 第一股 cDNA、各 4.0 μL 10 μmol·L-1的正向和反向引物、1.0 μLTaqplus DNA polymerase(5 U·μL-1)(TaKaRa,Otsu,Japan)、10 μLTaqplus Buffer、8.0 μL dNTP Mixture,用無菌水將反應液調至100 μL;94℃預變性3 min、94℃變性30 s、57℃退火30 s、72℃延伸1 min、最終72℃延伸5 min,反應共進行30個循環(huán)。將該DNA片段經(jīng)DNA純化試劑盒(天根生物科技公司,北京)純化后,作為第二次PCR的模板,設計一對分別含EcoR I和HindIII酶切位點的引物H-MQF(5'CGGAATTCTTACCATCTGAGGTACGGCTC 3')和HMQR(5'CCCAAGCTTTTAGAACCTGCAGCAGAAC CC 3'),用于擴增編碼hepcidin原前體肽的cDNA片段。PCR反應體系和條件同上,除了退火溫度改為58℃。目的片段“pIH”與pMD19-T simple質粒(TaKaRa,Otsu,Japan)按3:1比例、在T4DNA連接酶的作用下,16℃保溫2 h,轉化感受態(tài)細胞DH5α,37℃培養(yǎng)過夜;挑取5個陽性克隆由上海生工生物工程有限公司進行DNA測序。
1.4 pET32a-pIH重組表達質粒的構建
上述含“pMD19-pIH”重組質粒的陽性克隆經(jīng)LB液體培養(yǎng)過夜后,進行質粒提純(天根生物科技公司,北京),然后對其進行EcoRI和HindⅢ雙酶切,酶切產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳檢驗后,切下目的片段“pIH”進行DNA純化。另一方面,對pET32a(+)表達質粒(Novagen,Darmstadt,Germany)進行同樣的雙酶切,酶切產(chǎn)物經(jīng)0.5%瓊脂糖凝膠電泳檢驗后,切下線性質粒條帶進行DNA純化。將含酶切位點的目的片段“pIH”與線性的pET32a(+)表達質粒按7:1比例、在T4DNA連接酶的作用下,16℃保溫16 h;然后轉化感受態(tài)細胞DH5α同上進行DNA測序確證。
1.5 Hepcidin原前體肽的原核表達及表達產(chǎn)物的純化
上述含“pET32a-pIH”重組表達質粒的陽性克隆經(jīng)LB液體培養(yǎng)后,進行質粒提純。5 μL重組表達質粒轉化工程菌E.coliBL21(DE3),37℃培養(yǎng)過夜;挑選陽性克隆接種于20 mL含氨芐青霉素(100 mg·mL-1)的LB培養(yǎng)液中,37 ℃、150 r·min-1培養(yǎng)至OD600為0.6~1.0后,按1:50轉接到1 000 mL抗性LB培養(yǎng)液中,37℃、220 r·min-1培養(yǎng)至OD600為0.6~0.8;加入誘導劑IPTG至終濃度為1 mmol·L-1,37℃、110 r·min-1培養(yǎng)12 h后,將菌液離心10 min(4 ℃,8 000 r·min-1);用1/10培養(yǎng)液體積的1×PBS緩沖液(pH 7.4)重懸沉淀,冰上進行超聲波細胞破碎后,離心10 min(4 ℃,12 000 r·min-1);上清液通過Profinia蛋白純化系統(tǒng)(Bio-rad,Laboratories,Inc.,USA),經(jīng)固化金屬離子親和層析(IMAC)純化得到目的“融合蛋白”。
1.6 Tricine-SDS-PAGE分析
Tricine-SDS-PAGE分析參照Schagger的方法[12]。濃縮膠濃度4%,分離膠濃度16%,各樣品的上樣量為20 μL。電泳結束后用0.1%的考馬斯亮藍R-250對凝膠進行染色,用25%的甲醇和7%的醋酸混合液脫色。低分子質量標準蛋白由天根生物科技公司提供。
2 結果與分析
2.1 斑點叉尾鮰hepcidin原前體肽的cDNA克隆及其推斷的氨基酸序列
以斑點叉尾鮰肝臟的第一股cDNA為模板,擴增得到編碼hepcidin全長氨基酸序列的約291 bp的片段(見圖1A);進一步以該片段為模板,根據(jù)巢式PCR的原理,通過設計添加EcoRI和HindIII酶切位點的引物,擴增得到編碼hepcidin原前體肽(除去信號肽的氨基酸序列)的約236 bp的片段“pIH”(見圖1B)。由圖2可見,根據(jù)推斷的氨基酸序列,該片段編碼了由72個氨基酸殘基組成的hepcidin原前體肽(47個殘基組成的前體肽+25個殘基組成的成熟肽),且在成熟肽區(qū)域存在8個保守的半胱氨酸殘基,它們在空間結構上可形成四對二硫鍵。本文克隆的編碼原前體肽的cDNA序列與參考序列相比存在2個核苷酸差異,因而導致前體肽的2個氨基酸差異。經(jīng)DNA測序,證明兩個酶切位點已成功引入該片段(見圖3A)。
2.2 斑點叉尾鮰與其他生物之間的hepcidin原前體肽氨基酸序列比較
圖2顯示斑點叉尾鮰與其他魚類及哺乳動物之間的hepcidin原前體肽的氨基酸序列比較。結果表明,斑點叉尾鮰與長鰭叉尾鮰、黃顙魚之間的序列同源性分別為96%和72%,與斑馬魚的2個同工型之間顯示47%~48%的同源性;而與哺乳動物之間顯示很低的同源性,只有10%~15%。另一方面,不同生物之間在hepcidin原前體肽的氨基酸序列上存在多個保守性位點或區(qū)域,其中8個半胱氨酸殘基是高度保守的,證明它們與hepcidin的抗菌活性密切相關。
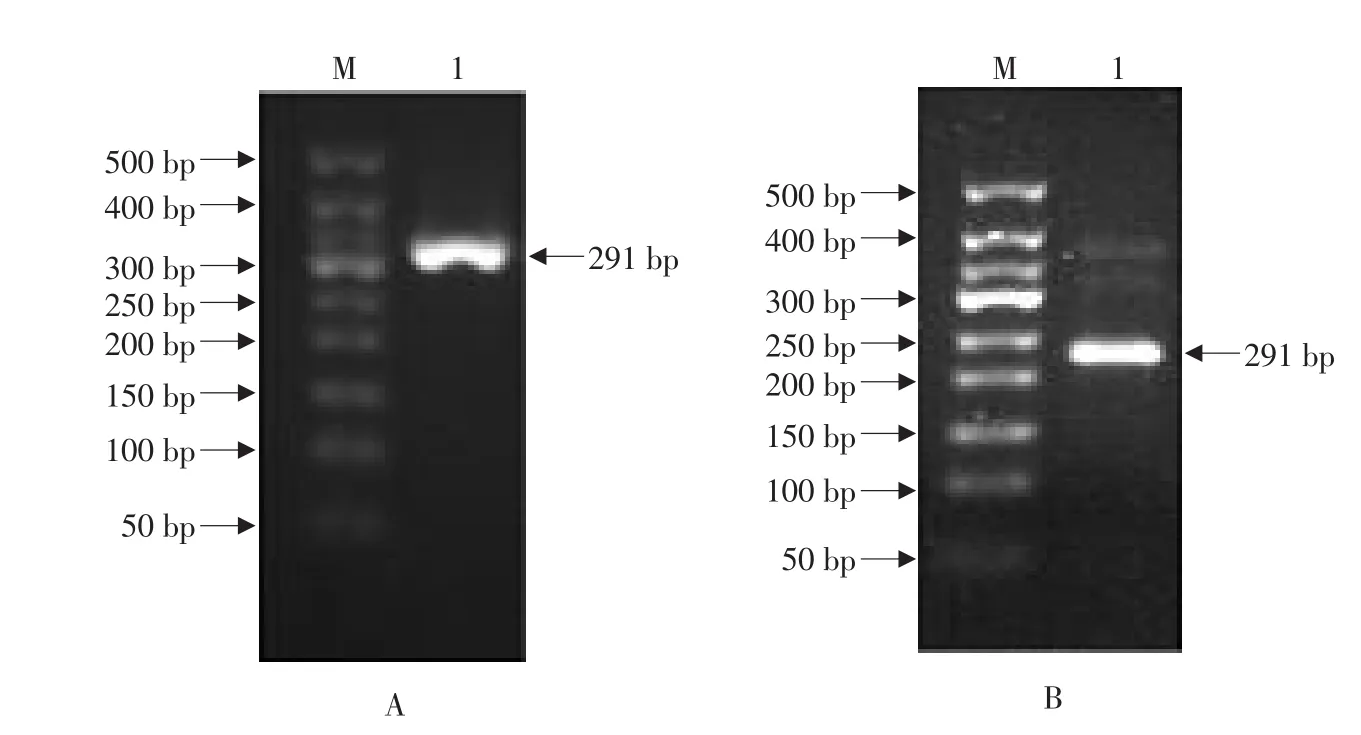
圖1 斑點叉尾鮰hepcidin全長(A)和原前體肽(B)cDNA的PCR擴增Fig.1 PCR amplification of cDNAs encoding the full-length hepcidin(A)and prepro-peptide(B)of channel catfish

圖2 斑點叉尾鮰與其他生物之間hepcidin原前體肽的氨基酸序列比對Fig.2 Comparison of the amino acid sequences for hepcidin prepro-peptides among channel catfish and other species
2.3 “pET32a-pIH”重組表達質粒的構建及鑒定
將含EcoR I和HindIII酶切位點的“pIH”片段與事先用這兩種酶處理過的pET32a(+)連接,得到重組表達質粒“pET32a-pIH”(見圖3B)。
在該重組表達質粒中,pET32a(+)所帶的高溶解性的硫氧還蛋白“trxA”的編碼基因與目的片段融合,形成的融合基因有利于高溶解性的融合蛋白的表達。通過菌落PCR鑒定,發(fā)現(xiàn)在約824 bp處有清晰條帶(見圖4A),與理論相符,初步確定為陽性菌落;進一步對陽性菌落中的質粒進行提純,通過EcoR I和HindIII對質粒進行雙酶切處理,經(jīng)1%瓊脂糖凝膠電泳后,發(fā)現(xiàn)存在大、小分子質量的兩條譜帶,其中一條約219 bp的小分子質量譜帶屬于目的片段(見圖4B)。最終通過對陽性克隆的測序,證明目的片段“pIH”已正確連接到pET32a(+)表達載體上,并且核苷酸序列未發(fā)生任何堿基突變。

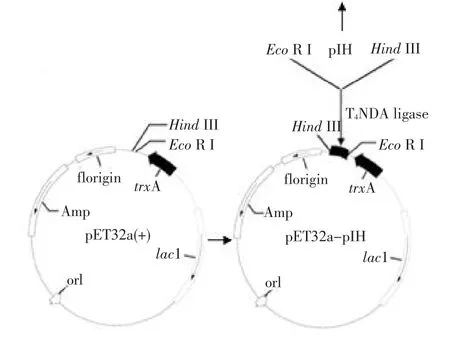
圖3 克隆的原前體肽序列(A)和重組表達質粒“pET32a-pIH”構建圖(B)Fig.3 Cloned prepro-peptide sequence(A)and construction of recombinant expression plasmid“pET32a-pIH”(B)
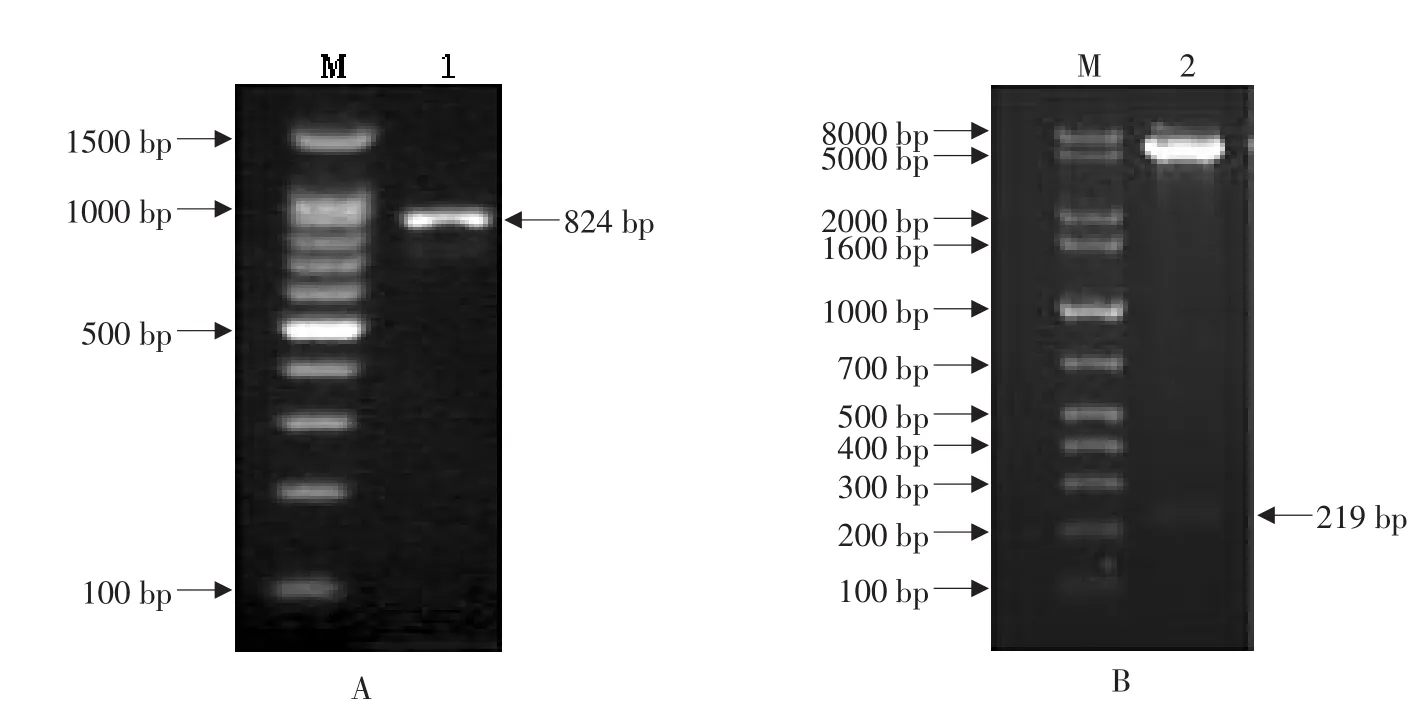
圖4 重組表達表質粒的PCR插入檢驗(A)和雙酶切鑒定(B)Fig.4 Identification of recombinant expression plasmid by colony PCR(A)and restriction enzyme digestion(B)
2.4 重組hepcidin原前體肽在E.coli BL21(DE3)中的融合表達及表達產(chǎn)物的純化
含重組表達質粒“pET32a-pIH”的工程菌E.coliBL21(DE3)在含氨芐青霉素的LB培養(yǎng)液中生長至OD600為0.6~0.8,經(jīng)1 mmol·L-1IPTG誘導,在37 ℃培養(yǎng)12 h后,成功表達了融合蛋白“trxA-pIH”(見圖5,泳道5)。細胞經(jīng)超聲破碎離心后,對上清和沉淀(包涵體)進行Tricine-SDS-PAGE分析,發(fā)現(xiàn)目的蛋白存在于上清(見圖5,泳道6)和沉淀(見圖5,泳道7)中,但前者較后者顯示更深的譜帶;經(jīng)Quantity One software,Version 4.4.0(Bio-Rad,Laboratories,Inc.,USA)對凝膠進行分析,發(fā)現(xiàn)融合蛋白的70%以上是可溶的,并且表達的融合蛋白約占細胞總蛋白的22.5%。而在陰性對照組中無任何表達(見圖5,泳道1-4)。通過IMAC進一步對上清中的含6xHis標簽的融合蛋白進行純化,經(jīng)Tricine-SDS-PAGE分析,可見分子質量約為33 ku的融合蛋白“trxA-pIH”的條帶,與預期的分子質量相符(見圖6)。
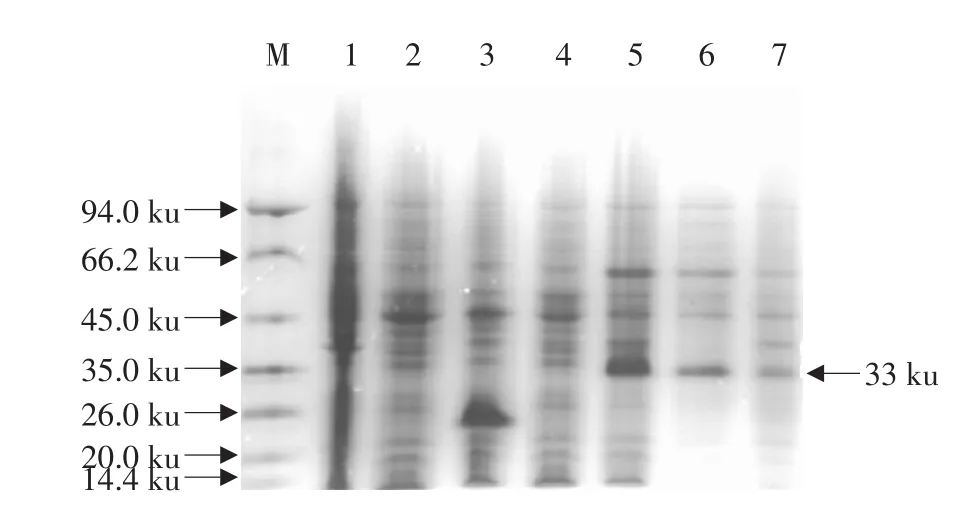
圖5 在E.coli BL21(DE3)中表達的融合蛋白的Tricine-SDS-PAGE分析Fig.5 Tricine-SDS-PAGE analysis for fusion proteins expressed in E.coli BL21(DE3)
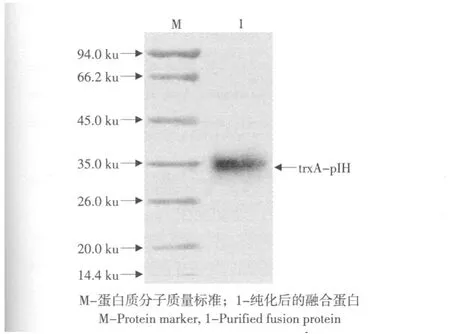
圖6 純化的融合蛋白的Tricine-SDS-PAGE分析Fig.6 Tricine-SDS-PAGE analysis for the purified fusion protein
3 討論與結論
通常在肝臟中合成的hepcidin抗菌肽的氨基端含有一段信號肽,在蛋白質翻譯后的修飾中該肽段被蛋白酶切除,剩下包括前體肽和成熟肽的原前體肽轉運進入血液中,再由前體肽轉化酶酶切掉前體肽后,形成具有生物活性的小分子成熟肽[18]。因成熟肽僅由20~25個氨基酸殘基組成,分子質量較小,可能不利于其在大腸桿菌中的表達,因此,本文以斑點叉尾鮰hepcidin原前體肽為研究對象,參考Bao等報道的斑點叉尾鮰hepcidin全長cDNA序列[11],通過RT-PCR從其肝臟克隆到編碼hepcidin原前體肽的cDNA序列。然而,與參考序列相比,該cDNA序列顯示了2個不同的核苷酸殘基,進而導致了原有的纈氨酸和精氨酸由丙氨酸和亮氨酸取代。據(jù)報道,Hepcidin基因在一些魚類中具有多個拷貝,例如,黑鯛(Acanthopagrus schlegelii)含七種hepcidin序列[19];Huang等從莫桑比克羅非魚(Oreochromis mossambicus)中分離到三種編碼hepcidin 的 cDNA 序列(TH1-5、TH2-2、TH2-3)[20]。因此,本文獲得的hepcidin原前體肽也可能屬于斑點叉尾鮰hepcidin抗菌肽的另外一種同工型。雖然來自不同生物的hepcidin原前體肽之間顯示了在氨基酸序列上的差異,但在它們的成熟肽區(qū)域均存在8個高度保守的半胱氨酸殘基,以致在空間構象上形成的4對二硫鍵決定了hepcidin的抗菌活性[21-22]。
本文選擇pET32a(+)作為表達質粒,它所攜帶的高溶解性的硫氧還蛋白“trxA”的編碼基因與目的片段“pIH”形成的融合基因有利于目的蛋白以高溶解性的融合蛋白的形式表達。事實證明,本文在E.coliBL21(DE3)中表達的“trxA-pIH”融合蛋白的70%以上是可溶的。通常,由于大腸桿菌細胞基質的還原環(huán)境,使得在大腸桿菌中直接表達的大部分含半胱氨酸的陽離子抗菌肽是沒有活性的,初步的抑菌試驗也證實了這一點。進一步的研究將關注于表達產(chǎn)物的復性,以使目的蛋白形成正確折疊的三維空間結構而具生物學活性。嘗試對于hepcidin成熟肽區(qū)域的重組DNA表達,也是進一步研究的目標。
[1] Boman H G,Nilsson I,Rasmuson B.Inducible antibacterial defense system inDrosophila[J].Nature,1972,237:232-235.
[2] 朱勇,徐建雄.抗菌肽飼料添加劑的研究進展[J].飼料工業(yè),2006,27(3):14-16.
[3] 張虹,陶妍.斑點叉尾鮰肝臟表達的抗菌肽-2(LEAP-2)在E.coli中的融合表達[J].上海交通大學學報,2010,28(1):70-75.
[4] 黃平,章懷云.魚類抗菌肽研究進展[J].中南林業(yè)科技大學學報,2009,29(2):170-174.
[5] Krausea A,Neitz S,Magerth J,et al.LEAP-1,a novelhighly disulfide-bonded human peptide,exhibits antmi icrobial activity[J].FEBS Letters,2000,480:147-150.
[6] Park C H,Valore E V,Waring A J,et al.Hepcidin,a urinary antimicrobial peptide synthesized in the liver[J].Biological Chemistry,2001,276(11):7806-10.
[7] 黨曉偉,李夢云,常娟,等.Hepcidin的研究進展[J].安徽農(nóng)業(yè)科學,2010,38(7):3351-3353.
[8] Douglas S E,Gallant J W,Liebscher R S,et al.Identification and expression analysis of hepcidin-like antimicrobial peptides in bony fish[J].Developmental and Comparative Immunology,2003,27(6-7):589-601.
[9] Huang P H,Chen J Y,Kuo C M.Three different hepcidins from tilapia,Oreochromis mossambicus:Analysis of their expressions and biological functions[J].Molecular Immunology,2007,44(8):1922-1934.
[10] Huang W S,Li S J,Cai L,et al.Cloning sequence analysis and gene organization of antimicrobial peptide hepcidin gene from Nile Tilapia(Oreochromis niloticus)[J].Journal of Xiamen University(Natural Science),2007,46(3):390-395.
[11] Bao B L,Peatman E,Li P,et al.Catfish hepcidin gene is expressed in a wide range of tissues and exhibits tissue-specific upregulation afterbacterialinfection[J].Developmentaland Comparative Immunology,2005,29:939-950.
[12] Schagger H,Von Jagow G.Tricine-sodium dodecyl sulfatepolyacrylamide gel electrophoresis for the separation of protein in the range from 1 to 100 ku[J].Analytical Biochemistry,1987,166:368-379.
[13] Gibert Y,Lattanzi V J,Zhen A W.BMP signaling modulates hepcidin expression in zebrafish embryos independent of hemojuvelin[J].Public Library of Science One,2011,6(1):372-379.
[14] Shike H,Shimizu C,Lauth X,et al.Organization and expression analysis of the zebrafish hepcidin gene,an antimicrobial peptide gene conserved among vertebrates[J]. Developmental and Comparative Immunology,2004,28(7-8):747-754.
[15] Harrison-Findik D D,Schafer D,Klein E,et al.Alcohol metabolism-mediated oxidative stress down-regulates hepcidin transcription and leads to increased duodenal iron transporter expression[J].Biological Chemistry,2006,281(32):22974-22982.
[16] Gordeuk V R,Miasnikova G Y,Sergueeva A I.Chuvash polycythemia VHLR200W mutation is associated with downregulation of hepcidin expression[J].Blood,2011,118(19):5278-5282.
[17] Mural R J,Adams M D,Myers E W.A comparison of whole-genome shotgun-derived mouse chromosome 16 and the human genom[J].Science,2002,296(5573):1661-1671.
[18]Shike H,Lauth X,Westerman M E,et al.Bass hepcidin is a novel antimicrobial peptide induced by bacterial challenge[J].European Journal of Biochemistry,2002,269:2232.
[19] Yang M,Wang K J,Chen J H,et al.Genomic organization and tissue-specific expression analysis of hepcidin-like genes from black porgy(Acanthopagrus schlegeliiB.)[J].Fish and Shellfish Immunology,2007,23:1060-1071.
[20] Huang P H,Chen J Y,Kuo C M.Three different hepcidins from tilapia,Oreochromis mossambicus:Analysis of their expressions and biological functions[J].Molecular Immunology,2007,44:1922-1934.
[21] 李華,安利國.鯉魚抗菌肽hepcidin的基因克隆、表達及其功能的研究[D].濟南:山東師范大學,2010.
[22] Papagianni M.Ribosomally synthesized peptides with antimicrobial properties:Biosynthesis,structure,function and applications[J].Biotechnology Advances,2003,21(6):465-499.
猜你喜歡
中學生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
數(shù)學年刊A輯(中文版)(2022年4期)2022-02-16 08:17:34
今日農(nóng)業(yè)(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
無線電通信技術(2021年4期)2021-07-13 08:58:28
無線電通信技術(2021年3期)2021-06-08 03:33:48
中學生數(shù)理化(高中版.高考數(shù)學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現(xiàn)代出版(2020年3期)2020-06-20 07:10:34
福利中國(2015年4期)2015-01-03 08:03:38
