三種不同鹽生植物下土壤動物群落研究
2012-08-08 12:23:44謝桐音謝桂林付榮恕
東北農業大學學報 2012年12期
付 穎,謝桐音,謝桂林,宋 杰,付榮恕
(1山東省疾病預防控制中心,濟南 250014;2.東北農業大學生命科學學院,哈爾濱 150030;3.山東師范大學生命科學學院,濟南 250014)
土壤動物是指其生活史中有一段時間在土壤中度過,且對土壤有一定影響的動物[1]。近年來,國內外學者研究的重點不僅集中在土壤動物分類、土壤動物在土壤物質循環、土壤發育、土壤理化性質、土壤生物群落維護等方面,對土壤動物與土壤微生物的相互關系,影響土壤動物耐受性的生態因子如:食物資源、生境狀況等,土壤動物在土壤質量評價體系中的指示作用,土壤動物在生態系統中作用等方面研究也很關注。當前對土壤動物的研究已有新進展[2-5]。在治理和改造鹽堿地的方法中,生物改良被認為是最具有生態效益、經濟效益的措施。開展黃河三角洲東營地區的鹽堿地土壤動物群落調查研究,可以比較全面認識該區域內土壤動物的種類及數量分布情況,客觀評價不同的耐鹽堿植物對鹽堿地的適應或改良效果,為合理開發鹽堿地及利用其生態系統資源提供基礎。
1 材料與方法
1.1 研究區概況
本研究在黃河三角洲東營鹽生植物園(118°04'~119°14'E,35°15'~38°16'N)內進行。該區位于暖溫帶,背陸面海,受歐亞大陸和太平洋的共同影響,屬于暖溫帶半濕潤大陸性季風氣候區。四季溫差明顯,年平均氣溫11.7~12.6℃。7月份溫度最高,平均氣溫26.7℃,極端最高氣溫41.9℃;1月份最冷平均氣溫-2.8℃,極端最低氣溫-23.3℃。年平均日照時數為2 590~2 830 h,各月平均日照時數以5月份最多,12月份最少;無霜期211 d;年平均降水量530~630 mm,70%的降雨集中在6~8月份;平均蒸發量為750~2 400 mm。土壤類型以鹽化潮土和濱海鹽土為主,含鹽量一般在0.4%~2.0%,養分含量低。
1.2 材料與方法
土壤動物群落組成結構與植被狀況密切相關[6-7]。該區地表植被主要為人工種植的鹽生植物,土壤易受到擾動,形成的局部小氣候差異明顯。依據人工種植植被、耐鹽程度和種植年限的不同,選取先鋒植物種:最大耐鹽度為1.6%的翅堿蓬(Suaeda heteropteraKitagawa)、最大耐鹽度為0.9%的馬藺(Iris lactea var.chinensi)和最大耐鹽度為1.2%的中亞濱藜(Atriplex centralasiatica)作為研究對象,共設6個采樣點。分別為:種植1年的翅堿蓬(I1)、連續種植10年的翅堿蓬(I2)、種植1年的馬藺(II1)、連續種植10年的馬藺(II2)、種植1年的中亞濱藜(III1)、連續種植3年的中亞濱藜(III2)。
在設置好的樣點內,用土壤環刀分層取樣,并利用WET土壤水分溫度電導率傳感器,同步測定水分、溫度、電導率三個生態因子。野外調查和室內試驗方法參見文獻[8]。
1.3 數據分析
土壤動物優勢類群為個體占總數的10%以上;常見類群為個體數占總數的1%~10%;稀有類群為個體數占總數的1%以下。
利用SPSS18.0、 Excel和BioDiversity Professional軟件分析統計數據,并計算土壤動物多樣性指數,分析土壤動物群落的結構。
Shannon-Weiner多樣性指數:H=-∑Pi·lnPi;
Pielou均勻性指數:E=H/lnS;
Simpson優勢度指數:C=∑(ni/N)2
式中,S為動物類群數,N為動物個體總數,ni為第i類群的個體數,Pi為i類群個體數占總個體數的比例。
種植1年的翅堿蓬(I1)、連續種植10年的翅堿蓬(I2)、種植1年的馬藺(Ⅱ1)、連續種植10年的馬藺(Ⅱ2)、種植1年的中亞濱藜(Ⅲ1)、連續種植3年的中亞濱藜(Ⅲ2)。
2 結果與分析
2.1 群落組成與數量分布
本調查共獲得各類土壤動物樣本10 195頭,隸屬于2門7綱,共計14個類群表1。由表1可見,蜱螨目(Acarina)和彈尾目(Collembola)占捕獲總量的比例最大,分別為79.66%(8121頭)和13.00%(1325頭),為黃河三角洲植物園區土壤動物群落的優勢類群;膜翅目(Hymenoptera)和鞘翅目(Coleoptera)占捕獲總量的比例為3.03%和2.07%,均大于1.00%,為常見類群;其余10個類群均不足捕獲總量的1.00%,為稀有類群。
土壤動物類群數在不同樣點中變化不明顯,最多的為II2樣點12種,最少的為I1、Ⅱ1、Ⅲ1樣點各9種。而土壤動物個體數變化有較大差異,順序依次為:Ⅱ2(48.69%)>Ⅲ2(11.35%)>Ⅲ1(11.32%)>Ⅱ1(10.81%)>I1(10.25%)>I2(7.58%)。
蜱螨目與彈尾目的數量比常可作為不同緯度上土壤動物區系的一種特征指標[9]。由于本文選擇的3種不同的鹽生植物及其種植年限的差異,導致對各樣點土壤鹽度的變化存在較大差異。由表1可見,A/C(蜱螨目個體數/彈尾目個體數):Ⅱ2(9.90)>Ⅱ1(6.58)>Ⅲ2(5.64)>I1(5.43)>Ⅲ1(3.53)> I2(1.80)偏離溫帶A/C約等于1這一規律,支持鹽度這一生態因子為該區土壤動物數量和種類的限制因素。
彈尾目和蜱螨目動物在調查的6個樣點中都有生存,說明二者具有廣泛的生態位。本文中蜱螨目與彈尾目捕獲的個體數量比例相差顯著,也表明該區較高的鹽度可能是導致彈尾目動物數量較少的直接原因。
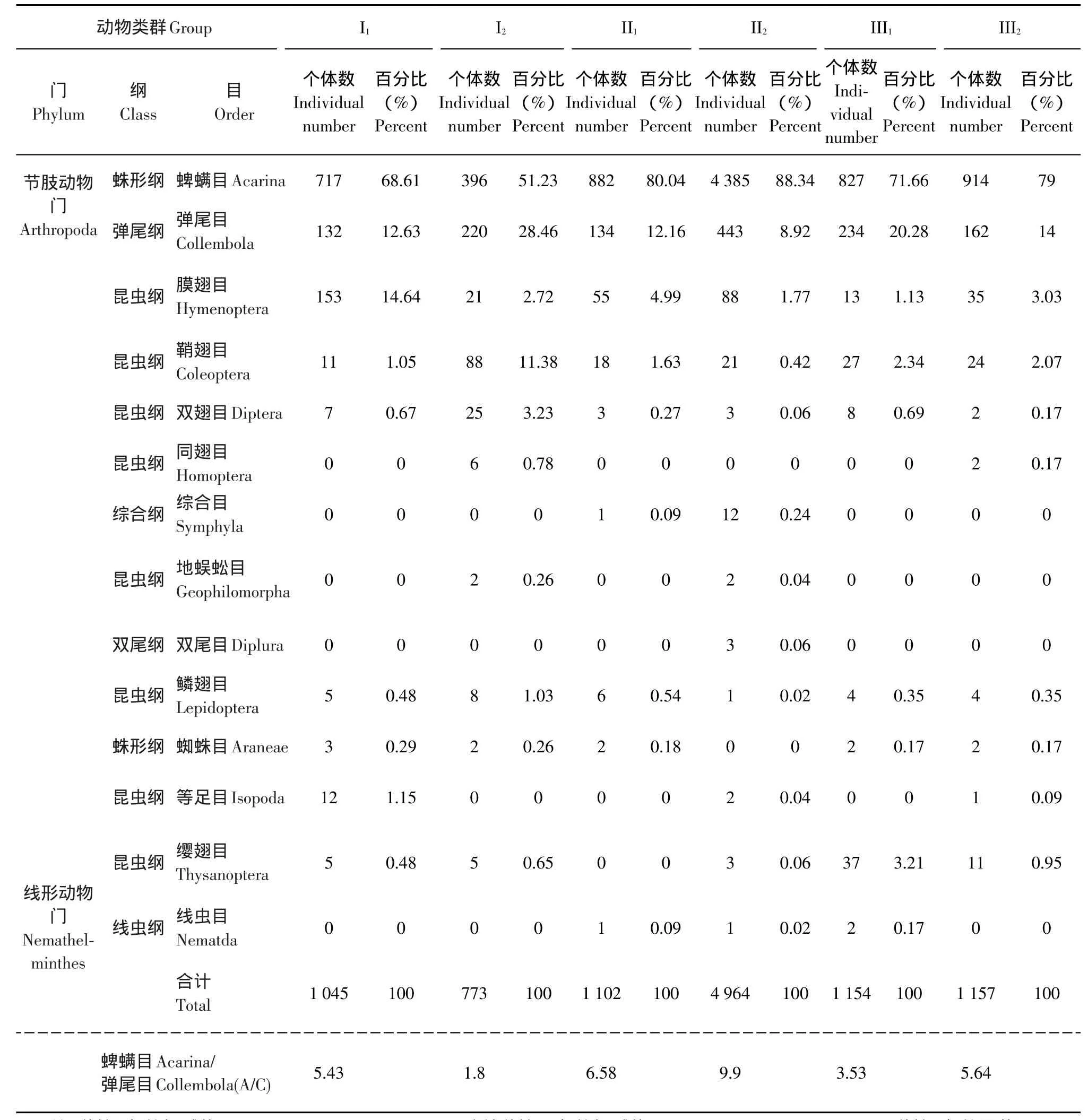
表1 土壤動物群落組成與數量分布Table 1 Soil animals component and quantity distribution
連續種植翅堿蓬(Suaedaheteroptera Kitagawa)、馬藺(Iris lacteachinensi)和中亞濱藜(Atriplex centralasiatica)這3種耐鹽堿先鋒植物,可以降低土壤中的含鹽量(鹽度)。有文獻研究表明:栽種翅堿蓬和馬藺后含鹽量降低了50%~81%不等[10]。由表1可知:連續種植10年的馬藺(Irislactea chinensi)(II2)捕獲的動物數量高出種植1年的馬藺(Iris lactea chinensi)(II1)3倍以上。說明含鹽量的降低,減輕了鹽度對土壤動物的脅迫作用,使土壤動物中耐受性強的種類個體數量得以維持在較高水平。
種植1年的中亞濱藜(Atriplex centralasiatica)(III1)和連續種植3年的中亞濱藜(Atriplex centralasiatica)(III2)捕獲的動物數量相當,而連續種植10年的翅堿蓬(Suaeda heteroptera Kitagawa)(I2)捕獲的動物數量比種植1年的翅堿蓬(Suaeda heteroptera Kitagawa)(I1)低,但均少于馬藺(Iris lactea chinensi)(II2)捕獲的動物數量。主要原因是翅堿蓬(Suaeda heteroptera Kitagawa)和中亞濱藜(Atriplex centralasiatica)為一年生草本植物,秋季多被收割,使得種植翅堿蓬和中亞濱藜的樣地一年中較長時間為光板地,水分運輸由植物蒸騰轉而被土壤地表水分蒸發取代,土壤返鹽堿嚴重,影響土壤動物的數量分布。
土壤動物數量多寡可反映耐鹽堿植物對環境的改良效果。連續種植10年的馬藺(Iris lactea chinensi)(II2)捕獲的動物數量高出種植1年的馬藺(II1)捕獲的動物數量,而種植1年的馬藺(Iris lactea chinensi)(II1)捕獲的動物數量與種植1年的中亞濱藜(Atriplex centralasiatica)(III1)、連續種植3年的中亞濱藜(Atriplex centralasiatica)(III2)捕獲的動物數量、種植1年的翅堿蓬(Suaeda heteroptera Kitagawa)(I1)捕獲的土壤動物的數量相當。說明新種植的馬藺(Iris lactea chinensi)由于根系還不發達,在改良土壤鹽度方面與翅堿蓬(Suaeda heteroptera Kitagawa)和中亞濱藜(Atriplex centralasiatica)相似。連續種植多年的馬藺(Iris lacteachinensi)根系發達,可吸收并減少土壤深處的含鹽量,同時植物蒸騰作用取代地表水分的直接蒸發,可以較大程度地減少所在地的含鹽量,使土壤動物維持在較高水平。
此外,草本植物由于根系分布深淺不同,對含鹽量的影響也存在差異[10]。翅堿蓬(Suaeda heteroptera Kitagawa)為一年生草本植物,根系分布較淺,對10~20 cm土壤層鹽分含量影響最大,種植后使土壤含鹽量減少81%,隨著土層的加深,影響逐漸減少;而馬藺(Iris lactea chinensi)是多年生宿根草本植物,根莖葉粗壯,須根稠密發達,長度可達1 m以上,呈傘狀分布。對土層20~50 cm影響最大,達到65%,但對表層土層含鹽量變化的影響逐漸減少。因此可以嘗試間作馬藺(Iris lactea chinensi)和其他一年生鹽生草本植物改良鹽堿地,以期達到更好效果。
2.2 中小型土壤動物群落的多樣性
土壤動物類群數、密度和土壤動物多樣性是土壤動物研究的基礎性內容[11]。多樣性指數、均勻性指數和優勢度指數等可以表示群落的結構和功能特征。群落的多樣性指數、均勻性指數和優勢度指數,取決于群落的類群數與各類群動物的個體數量,是可以直接測定的指標[12]。電導率是測定土壤水溶性鹽的指標,是判定土壤中鹽類離子是否限制土壤動物生存生長的重要因素。
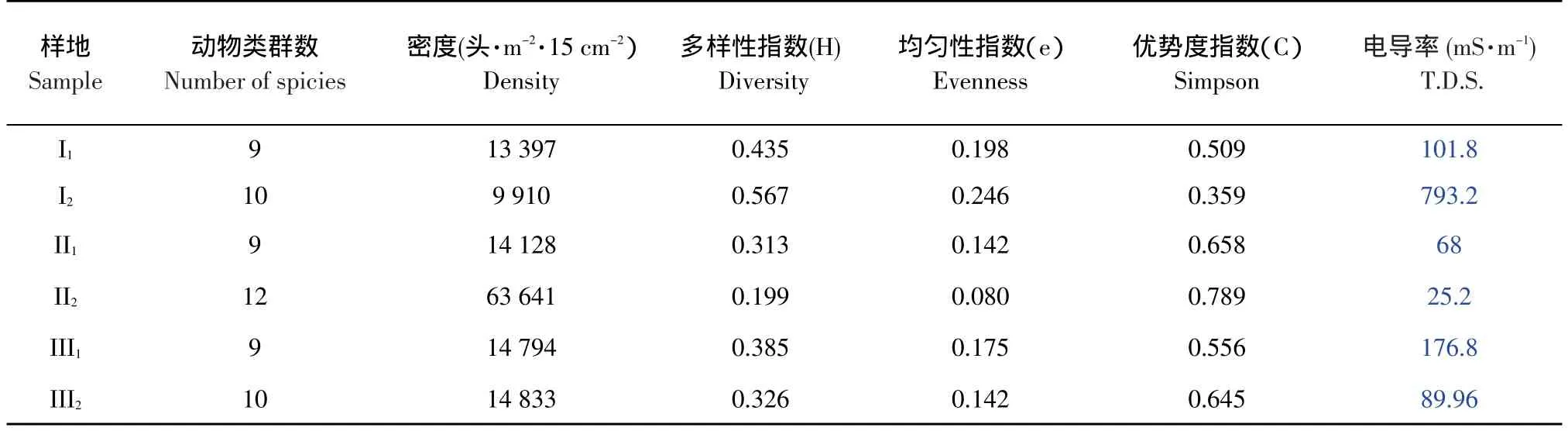
表2 土壤動物群落結構的主要指標Table 2 Some indexes of soil mesofauna structure
由表2可見,在各個樣地中,群落多樣性指數,最高的為連續種植10年的堿蓬樣地(Ⅰ2),H=0.567,最低的為連續種植10年的馬藺樣地(Ⅱ2),H=0.199,群落多樣性指數的這一變化與群落的類群數和各類群動物的個體數量相關,群落類群數越多、各類群動物個體數越均勻,多樣性指數就越大,反之越小。均勻性指數最高為連續種植10年的堿蓬樣地(Ⅰ2)為0.246,最低的為連續種植10年的馬藺樣地(Ⅱ2),僅為0.080。優勢度指數與均勻性指數相反,均勻性指數越大,優勢度指數越低。優勢度指數最高為連續種植10年的馬藺樣地(Ⅱ2),為0.789。優勢度指數最低的為連續種植10年的堿蓬樣地(I2),為0.359。
一般來說,土壤水分中含鹽量越大,電導率就越大。但由于不同的離子導電性和質量的不同,含鹽量與電導率之間并無嚴格意義上的對應關系。隨著耐鹽堿植物種植年限的增加,土壤水分中的含鹽量降低,鹽度下降,電導率值就越小,土壤動物類群數和密度增加。因此,電導率與密度成反比這一關系,可以粗略估計土壤含鹽量變化較大的地區土壤動物的數量高低。
在馬藺和中亞濱藜樣點,電導率下將的原因是它們生長茂密,蓋度較高,植物的蒸騰作用取代土壤中水分的蒸發作用,使土壤水分中的鹽分積存在土壤深層或積累在鹽生植物中,從而避免土壤耕作層鹽分的積累。而堿蓬雖能夠在生長期內調節小氣候,減少水分蒸發,但是在抑制鹽度上升、防止土壤返鹽方面效果不如馬藺和中亞濱藜明顯,使電導率不降反升。
2.3 土壤動物的垂直分布
由于土壤各層內有機質和營養物質的含量、理化特性和水熱條件有差異,導致了土壤動物在各層的分部差異[13]。土壤動物有明顯的表聚現象,一般同大多數優勢類群和常見類群有明顯的表聚性有關[14],黃河三角洲鹽生植物園土壤動物垂直分布結果見圖1。
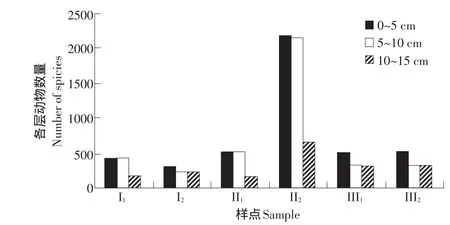
圖1 土壤動物垂直分布Fig.1 Vertical distribution of soil animals
垂直分布結果顯示,該地區種植1年的馬藺(II1)、種植1年的中亞濱藜(III1)、連續種植3年的中亞濱藜(III2)的土壤動物在0~5 cm,5~10 cm,10~15 cm土壤層具有明顯的表聚現象。受表層鹽度梯度影響,種植1年的堿蓬(I1)、連續種植10年的馬藺(II2)中的中小型土壤動物表聚現象不明顯,在0~10cm范圍內土壤動物均勻分布;連續種植10年的堿蓬(I2)、種植1年的中亞濱藜(III1)、連續種植3年的中亞濱藜(III2),在5~15 cm范圍內土壤動物均勻分布。
另外,同一類群個體數量在不同樣點中隨土層加深而遞減的程度不同,彈尾目下降速度最快,蜱螨目次之。蜱螨目、彈尾目和蜘蛛目主要集中分布在0~5 cm層,鞘翅目在5~10 cm土壤層中個體數量最高。
3 討論與結論
本研究共獲得各類土壤動物樣本10 195頭,隸屬于2門7綱,共計14個類群。蜱螨目(Acarina)和彈尾目(Collembola)占捕獲總量的比例最大為該地區土壤動物群落的優勢類群;膜翅目(Hymenoptera)和鞘翅目(Coleoptera)占捕獲總量的比例,均大于1.00%,為常見類群;其余10個類群均不足捕獲總量的1.00%,為稀有類群。優勢類群和常見類群占到全部捕獲量的98.09%,構成了黃河三角洲東營植物園區土壤動物的主體,對土壤動物群落特征起著決定性作用。
各類土壤動物受鹽度影響,差異較大,優勢類群中彈尾目動物—跳蟲捕獲量偏少,使得A/C(蜱螨目個體數/彈尾目個體數):II2(9.90)> II1(6.58)> III2(5.64)> I1(5.43)> III1(3.53)> I2(1.80),偏離溫帶A/C約等于1這一規律。
中小型土壤動物多樣性指數(Shannon-Weiner多樣性指數)全周年內都沒有超過1.0,這主要是蜱螨目和彈尾目兩個優勢類群占總體的比例過大造成的。對這種現象可能的解釋是,蜱螨目動物和彈尾目動物中的個別種類相比其它生物種類,對該地區土壤有較高的pH(平均為7.8)、較高的含鹽量(平均約為1.0%)和養分含量低的生境,有更強的耐受性。
土壤動物的優勢類群彈尾目對土壤pH具有指示作用[15]。本次調查發現球角跳在該地區密度較大,連續種植10年的堿蓬(I2)、種植1年的馬藺(II1)、連續種植10年的馬藺(II2)、種植1年的中亞濱藜(III1)樣點中類符跳個體數量較高。在立地條件較好的連續種植10年的馬藺(II2)樣點中跳蟲種類比較豐富且所獲個體數量較其它樣點多,其它樣點跳蟲種類和個體數量較少,這與已有報道[16]的結果一致。
本研究分析該區域土壤動物的種群數量和群落結構,并對影響土壤動物數量動態的生態因子進行分析,揭示人工種植植被下土壤動物多樣性變化特點及主要影響因素,為鹽堿地改良與生態建設提供了科學依據。
[1] 尹文英.中國亞熱帶土壤動物[M].北京:科學出版社,1992:4-6.
[2] 尹文英.中國土壤動物檢索圖鑒[M].北京:科學出版社,1996:1-6.
[3] 朱永恒,趙春雨,王宗英,等.我國土壤動物群落生態學研究綜述[J].生態學雜志,2005,24(12):1477-1481.
[4] 葛寶明,孔軍苗,程宏毅,等.不同利用方式土地秋季大型土壤動物群落結構[J].動物學研究,2005,26(3):272-278.
[5] Filser J.The role of Collembola in carbon and nitrogen cycling in soil[J].Pedobiologia,2002,46:234-235.
[6] 焦向麗,朱教君,閆巧玲.遼東山區次生林生態系統大、中型土壤動物組成與季節動態[J].生態學報,2009,29(5):2631-2638.
[7] 張雪萍,李春艷,殷秀琴,等.不同使用方式林地的土壤動物與土壤營養元素的關系[J].應用與環境生物學報,1999,5(1):26-31.
[8] 謝桐音,付榮恕,田家怡.黃河三角洲貝殼堤島中小型土壤動物群落結構[J].山東師大學報:自然科學版,2010,25(2):112-118.
[9] 付榮恕,田家怡,張蓬軍,等.鶴伴山國家森林公園土壤動物群落結構的研究[J].山東師范大學學報:自然科學版,2005,20(4):76-79.
[10] 王玉珍,劉永信,魏春蘭,等.6種鹽生植物對鹽堿地土壤改良情況的研究[J].安徽農業科學,2006,34(5):951-954.
[11] 吳海濤,呂憲國,楊青,等.土壤動物主要生態特征與生態功能的研究進展[J].土壤學報,2006,43(2):314-323.
[12] 馬克平,劉玉明.生物群落多樣性的測定方法I:α多樣性的測定方法(下)[J].生物多樣性,1994,2(4):231-239.
[13] Fu B Q,Chen W,Dong X H,et al.The composition and structure of the four soil macrofaunas inSongshan mountainin Beijing[J].Acta Ecologica Sinica,2002,22:215-233.
[14] Liu H,Yuan X Z.Ecological distribution of the soil animals in Taishan Mountain[J].Chinese Journal of Ecology,1999,18:13-16.
[15] 陳建秀,麻智春,嚴海娟,等.跳蟲在土壤生態系統中的作用[J].生物多樣性,2007,15(2):154-161.
[16] Ke X,Yue Q Y,Fu R H,et al.Soil fauna community and bio-assessment of soil acidity in beach land of eastern Shanghai[J].Zoological Research,2002.23(2)129-135.
