民豬CIRP基因的克隆與冷誘導研究
2012-08-08 12:23:28張冬杰楊國偉
東北農業大學學報 2012年12期
張冬杰,劉 娣,汪 亮,別 墅,楊國偉
(黑龍江省農業科學院,哈爾濱 150086)
冷誘導RNA結合蛋白(Cold inducible RNA-binding protein,CIRP)是在哺乳動物細胞中發現的首種冷應激或冷休克蛋白,伴隨冷應激過程過量表達[1]。日本Nishiyama實驗室首次從小鼠睪丸細胞中分離到冷誘導RNA結合蛋白,發現溫和冷處理(29~22℃)可顯著提高小鼠CIRP mRNA水平,相反熱處理(39和42℃)則降低其表達水平 。日本Saito實驗室研究證實牛蛙腦中CIRP在冬季強烈表達,而在夏季表達量很低。這表明CIRP表達可能在動物冬眠過程中發揮一定作用[3]。日本Aoki實驗室和Nishiyama實驗室分別證實CIRP具有RNA分子伴侶活性[4]。英國Banks實驗室在發育的非洲爪蟾神經組織內發現該基因瞬時表達,這表明在神經發生過程中發揮重要作用[5]。我國從2005年對CIRP進行研究。李安民實驗室報道CIRP在腦缺血時表達及亞低溫下冷誘導CIRPmRNA在大鼠腦內的表達情況[6];楊煥民實驗室克隆BALB/C鼠睪丸組織中CIRP,并制備與鑒定CIRP多克隆抗體[7]。為進一步探討CIRP基因在豬上的生物學功能,本研究克隆民豬的CIRP基因mRNA序列,對該基因在不同組織的表達水平進行了檢測,并分析冷誘導下該基因的表達變化情況。
1 材料與方法
1.1 材料
75日齡民豬3窩共計12頭,將每窩個體隨機分成2組即常溫組和低溫組,每組各6頭,購自黑龍江省農科院畜牧研究所。2009年12月25日到2010年1月6日期間,將常溫組置于正常舍內飼養,溫度控制在(10±2)℃,低溫組在舍外半敞式大棚內飼養,溫度為(-20±3)℃,兩組均飼喂相同的飼料。處理結束后,屠宰,取腿部肌肉組織各10 g,以液氮保存帶回實驗室,-80℃冰箱凍存。
感受態細胞(購自北京原平皓生物技術有限公司);rTaq酶、限制性內切酶BamHⅠ和HindⅢ、T 載體、TaKaRa RNA PCR Kit(AMV)Ver3.0(購自TaKaRa公司);膠回收試劑盒和質粒提取試劑盒(購自天根生化科技有限公司);SYBR Green PCR Master Mix(購自美國ABI公司);TRIZOL(購自Invitrogen公司)。
1.2 方法
1.2.1 民豬CIRP基因的克隆
以GenBank上已公布的人CIRP基因序列(NM_001280)為源序列,使用BLAST在線軟件進行相似序列搜索,找到一條豬的完整EST序列(AK235139),經反復比對后,初步認定為該EST序列即為豬的CIRP基因序列,據此共設計2對引物進行CDS全長序列的擴增,引物序列如下:CIRP1F: 5'GCCGTTGTGCTGTGCTGTCT 3', CIRP2R:5'TAGTCCCTGGAGCCGCCATA 3',預計擴增長度510 bp左右;CIRP3F:5'GCTCCAGGGACTACTA CAGC 3',CIRP4R:5'CCTTAGCCACTTCAAATA CAACAT 3',預計擴增長度800 bp。PCR擴增結束后使用膠回收試劑盒回收目的片段,連入T載體后,轉化入大腸桿菌,過夜培養后,搖菌提質粒,送交北京華大生物公司測序。
1.2.2 生物信息學分析
用DNAMAN6.0和DNAStar7.0軟件對所測得序列進行拼接和分析;用ProtParam tool軟件進行氨基酸組成、等電點分析;利用用NCBI的Conserved Domains程序對蛋白的保守結構域進行預測;使用TMpred和TMHMM對蛋白質進行跨膜區分析;利用NetPhos 2.0 Server和NetPhosK 1.0 Server軟件對蛋白質進行磷酸化位點的預測。
1.2.3 冷誘導后CIRP基因的表達變化
將常溫組和低溫組個體肌肉的cDNA分別混合,獲得2個cDNA池,分別記為常溫組和低溫組。設計1對特異性引物用以Real-time PCR檢測,選擇β-actin基因作為看家基因,引物序列如下 : CIRP5F: 5'TGTTCCTGAGCGTAGCAT 3',CIRP6R: 5'TCCTTAGCCACTTCAAATA 3', actinF:5'CGGGACCTGACCGACTACCT 3',actinR:5'GG GCCGTGATCTCCTTCTG 3'。分別以這2個cDNA池為模板,采用二步法進行Real-time PCR擴增。反應體系如下:上、下游引物各0.8 μL,Mix 10 μL,模板0.8 μL,H2O 7.6 μL,共計20 μL。反應程序如下:95℃10 min,95℃30 sec,60℃1 min,40個循環。反應結束后,獲得每個樣本的Ct值,使用如下換算公式進行目的基因最終的相對定量結果:2-△△ct,其中ΔΔCt=(Ct,目的基因-Ct,管家基因)實驗組-(Ct,目的基因-Ct,管家基因)對照組。
2 結果與分析
2.1 民豬CIRP基因序列分析
克隆測序獲得了民豬CIRP基因的3個轉錄變異體,轉錄變異發生在編碼區內。CIRP變異體1基因cDNA長1 278 bp(GenBank登錄號:HQ 908794),其中包括93 bp的5'UTR區,519 bp完整的CDS編碼區以及666 bp的3'UTR區,編碼172個氨基酸,終止密碼子為TAA。ORF由113個腺嘌呤(A)、191個鳥嘌呤(G)、98個胸腺嘧啶(T)和117個胞嘧啶(C)組成,四種堿基的含量依次為21.77%、36.80%、18.88%和22.55%,G、C(59.36%)含量高于A、T(40.65%)含量。
CIRP變異體2基因cDNA長1 653 bp(GenBank登錄號:HQ908795),與變異體1相比,在其編碼區的第501堿基處,出現了一段375 bp的插入片段,該插入片段的第46~48個堿基為TAG,使翻譯提前終止,共編碼182個氨基酸。ORF由115個腺嘌呤(A)、207個鳥嘌呤(G)、105個胸腺嘧啶(T)和122個胞嘧啶(C)組成,四種堿基的含量依次為20.95%、37.70%、19.13%和22.22%,G、C(59.92%)含量高于A、T(40.08%)含量。
CIRP變異體3長1 765 bp(GenBank登錄號:HQ908796),與變異體2相比,在其編碼區的第428堿基處,出現了一段115 bp的插入片段,該插入片段的第5~7個堿基為TGA,使翻譯提前終止,共編碼144個氨基酸。ORF由91個腺嘌呤(A)、164個鳥嘌呤(G)、86個胸腺嘧啶(T)和94個胞嘧啶(C)組成,四種堿基的含量依次為20.92%、37.70%、19.77%和21.61%,G、C(59.31%)含量高于A、T(40.69%)含量。3種轉錄變異體的核苷酸和氨基酸序列比對結果見圖1和圖2。

用NCBI的Conserved Domains程序(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)預測該蛋白的保守區域,結果發現在該基因的氨基端具有1個高度保守的RNA結合域(RNA recognition motif)。該保守區域由73個氨基酸組成(第7到第80位氨基酸),其中包含2個高度保守序列,分別為1個氨基酸六聚體(序列為LFVGGL,位置:第8到第13位氨基酸)和1個氨基酸八聚體(序列為RDFGFVTF,位置:第47到第54位氨基酸),其他一些疏水性氨基酸摻雜在這些保守序列中。使用TMpred(http://www.ch.embnet.org/software/TMPRED_form.html)和 TMHMM(http://www.cbs.dtu.dk/services/TMHMM/)在線軟件分析,CIRP不存在跨膜結構域,初步認為冷誘導RNA結合蛋白應是一種存在于細胞質中的蛋白質。NetPhos 2.0 Server(http://www.cbs.dtu.dk/services/NetPhos/)預測 CIRP 三種轉錄蛋白的磷酸化位點結果如表1所示。使用在線軟件NetPhosK 1.0 Server預測特異蛋白激酶的磷酸化位點,臨界值設為0.6,預測結果顯示,變異體1存在7個特異蛋白激酶的磷酸化位點(3個PKA,1個PKB,3個PKC),變異體2存在7個特異蛋白激酶的磷酸化位點(2個PKA,1個PKB,4個PKC),變異體3存在4個特異蛋白激酶的磷酸化位點(2個PKA,1個PKB,1個PKC)。

表1 NetPhos 2.0 Server軟件預測CIRP蛋白的磷酸化位點Table 1 Prediction phosphorylation sites of CIRP protein by NetPhos 2.0 Server
2.2 冷誘導后CIRP基因表達變化結果
將常溫組民豬的CIRP基因mRNA表達量設定為1,以相對表達水平的差異倍數為縱坐標,測得3個重復的CIRP基因mRNA動態表達水平見圖3。t-test結果顯示民豬的CIRP基因的mRNA水平經冷誘導后出現了顯著的上升(P<0.05)。
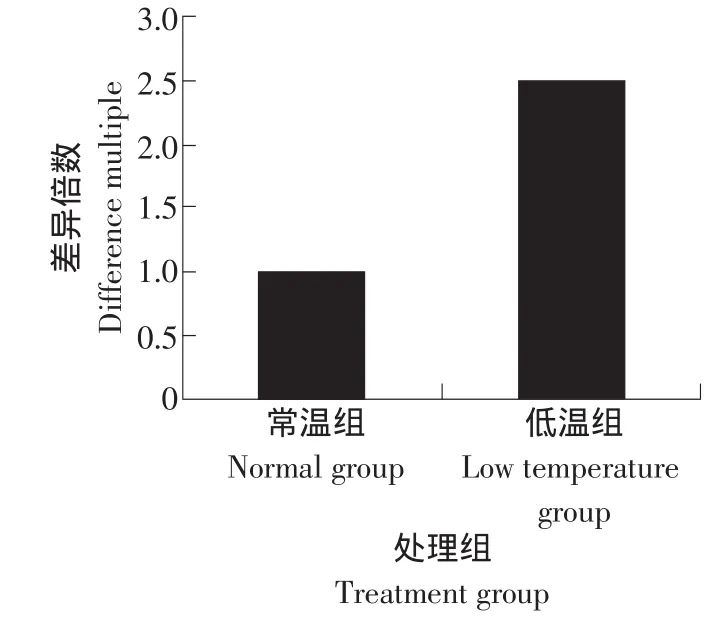
圖3 冷誘導后CIRP基因表達變化結果Fig.3 Expression of CIRP gene after cold-induced
3 討論與結論
溫度降低可以降低組織器官的代謝率,并且對心、腦、肝臟等主要臟器功能有明顯的保護作用,這種保護作用很可能是通過冷休克蛋白(Cold shock proteins,CSPs)在低溫條件下的高表達來實現的,但具體是通過何種途徑產生保護作用的機理目前仍不清楚,高等生物中CIRP和RBM3是研究相對較多的CSPs。CIRP廣泛存在于植物、人類、小鼠、大鼠、非洲爪蟾、牛蛙、鮭魚等生物體內,在神經系統發育、胚胎發育等許多進化過程中發揮重要的作用。但到目前為止,還沒有見到關于豬的CIRP基因的相關報道。因此本研究以人的CIRP基因序列為源序列,BLAST比對后發現了一條已公布的完整的豬的EST序列,但按照此序列進行克隆測序后,竟發現3個轉錄變異體,它們在5'端高度保守,但在3'端出現較大變異,而且由于插入序列的存在,使得變異體2和變異體3均出現轉錄提前終止情況。這與Fageeh等在鼠上的研究結果略有不同[8],他們在擴增鼠的CIRP基因5'-UTR區時發現3個轉錄本,由于轉錄本長度的不同造成轉錄起始位點的改變,進而產生了3個轉錄變異體,而且3個轉錄變異體在不同溫度下NIH-3T3細胞系的穩定性和活性均存在差異,據此推測CIRP表達受到不同長度的mRNA所調控。
冷應激對活體生物基因表達的影響少見報道,和細胞相比生物體具有更加復雜的冷應激調節機制,冷應激會對動物的生產性能[9]、免疫[10]、神經內分泌系統[11]均產生不同程度影響。已報道的關于溫度變化對CIRP基因的表達影響結果基本一致,即高溫會對該基因的表達產生抑制作用[12],而低溫則可明顯地誘導該基因的表達。本研究結果與前人的研究結果一致,即對民豬低溫冷處理后,肌肉中CIRPmRNA的表達水平出現顯著上升。“冷應激會上調CIRP基因的表達”這一結論已基本得到證實,但其上調表達的作用機理以及生物學意義仍是目前研究的熱點。有人認為CIRP可能是通過激活細胞外信號調節激酶(Extracellular signalregulated kinase,ERK)信號通路來保護細胞免受低溫傷害[12]。
本研究克隆獲得了民豬CIRP基因的3種變異體,3種轉錄變異體的核苷酸序列同源性為86.45%,氨基酸序列同源性為87%。冷誘導后,CIRP基因在肌肉中的表達水平出現顯著升高(P<0.05)。
[1] 李士澤,金福厚,趙巧香,等.冷誘導RNA結合蛋白的生物學功能研究進展[J].生理科學進展,2009,40(3):271-274.
[2] Nishiyama H,Itoh K,Kaneko Y,et a1.A glycine-rich RNA-binding protein mediating cold-inducible suppression of mammalian cell growth[J].J Cell Biol,1997,137:899-908.
[3] Saito T,Sugimoto K,Adachi Y,et al.Cloning and characterization of amphibian cold inducible RNA-binding protein[J].Biochem Physiol B Biochem Mol Biol,2000,125(2):237-245.
[4] Aoki K,Ishii Y,Matsumoto K,et a1.Methylation of Xenopus CIRP2 regulates its arginine and glycine-rich region·mediated nucleocytoplasmie distribution[J].Nucleic Acids Res,2002,27:5182-5192.
[5] Banks S,King SA,Irvine DS,et a1.Impact of a mild scrotal heat stress on DNA integrity in murine spermatozoa[J].Reproduction,2005,126:505-514.
[6] 薛菁暉,張志文,李安民,等.冷誘導RNA結合蛋白(CIRP)在腦缺血時的表達[C].中國醫師協會神經外科醫師分會首屆全國代表大會論文集,2005.
[7] 趙巧香,張秀玲,紀麗麗,等.重組BALB/C鼠冷誘導RNA結合蛋白CIRP多克隆抗體的制備與鑒定[J].應用與環境生物學報,2009,15(5):634-637.
[8] Al-Fageeh MB,Smales CM.Cold-inducible RNA binding protein(CIRP)expression is modulated by alternative mRNAs[J].RNA,2009,15(6):1164-1176.
[9] 喬巖瑞.冷應激對豬生產性能的影響[J].飼料廣角,2004(3):23-26.
[10] 李瑞剛.冷應激對小鼠免疫系統的影響[D].呼和浩特:內蒙古農業大學,2005.
[11] 屠云潔,耿照玉,陳國宏,等.冷應激對家禽神經內分泌系統的影響[J].安徽農業科學,2009,37(24):11555-11557.
[12] Sakurai T,Itoh K,Higashitsuji H,et al.Cirp protects against tumor necrosis factor-alpha-induced apoptosis via activation of extracellular signal-regulated kinase[J].Biochim Biophys Acta.,2006,1763(3):290-295.
