夏氏瘧原蟲感染早期易感和抵抗小鼠樹突狀細胞亞群和表型的變化特點*
2011-06-06 13:24:20武靜靜潘艷艷武劍華曹雅明
中國人獸共患病學(xué)報 2011年1期
武靜靜,劉 軍,鄭 偉,潘艷艷,李 瑩,延 娟,武劍華,曹雅明,鄭 麗
2.徐州醫(yī)學(xué)院病原生物學(xué)與免疫學(xué)教研室,徐州221002;
3.沈陽軍區(qū)總醫(yī)院檢驗科,沈陽 110016
瘧疾與其他大多數(shù)感染性疾病一樣,需要通過免疫效應(yīng)機制對其控制和消除。目前,鼠瘧的研究已經(jīng)證實,CD4+T細胞在抗紅內(nèi)期瘧疾的保護性免疫應(yīng)答中起關(guān)鍵作用,夏氏瘧原蟲感染(P lasmodium chabaudi chabaudi,AS)期間,產(chǎn)生 IFN-γ的Th1細胞免疫應(yīng)答的適當(dāng)建立和隨后的Th2型細胞因子輔助B細胞產(chǎn)生抗體對控制和清除蟲血癥至關(guān)重要,明顯影響著感染進程和最終結(jié)局[1]。不同小鼠在感染同種瘧原蟲時,由于感染早期Th1應(yīng)答建立和水平的維持存在差異,最終會出現(xiàn)死亡和自愈兩種截然不同的結(jié)局。因此,Th1細胞免疫應(yīng)答誘導(dǎo)和調(diào)控機制的闡明將為瘧疾疫苗的研制提供新的靶點。
樹突狀細胞(Dendritic cells,DCs)在擴大固有免疫應(yīng)答,啟動適應(yīng)性免疫,形成Th應(yīng)答中發(fā)揮重要作用[2]。小鼠脾臟DCs由不同種類的細胞亞群構(gòu)成,即髓樣CD11c+CD11b+DCs和漿樣CD11c+CD45R/B220+DCs[3-4],兩種DCs亞群形態(tài)與功能不同,在病原體刺激下表現(xiàn)不同的增殖水平[3]。我們前期的研究顯示,P.c.AS感染早期,易感DBA/2小鼠和抵抗BALB/c小鼠均產(chǎn)生以IFN-γ分泌增加為主的Th1型細胞免疫應(yīng)答[5],并且相關(guān)研究表明P.c.AS感染的DBA/2小鼠死亡與免疫病變相關(guān)[6]。本研究進一步利用成功建立的AS感染的DBA/2和BALB/c小鼠模型,對比觀察了DCs亞群和表型的變化特點,以期探討瘧疾感染早期DCs在Th1細胞免疫應(yīng)答活化過程中的作用地位。
1 材料與方法
1.1 瘧原蟲及實驗動物感染 6~8w齡、雌性DBA/2和BALB/c小鼠各12只(中國醫(yī)學(xué)科學(xué)院實驗動物研究所提供,許可證編號:SCXK京200420001)經(jīng)腹腔感染1×106P.c.AS(日本愛媛大學(xué)分子寄生蟲學(xué)教研室惠贈)寄生的紅細胞(Parasitized Red Blood Cell,pRBC),感染不同時間的小鼠經(jīng)尾靜脈采血,制備薄血膜,Giemsa染色,鏡檢計數(shù)紅細胞感染率。
1.2 主要儀器及試劑 流式細胞儀(FACS Calibur,BD);FcγⅢ/Ⅱ封閉抗體(2.4G2克隆,BD Pharmingen)、抗CD11c-FITC單抗(H L3克隆,BD Pharmingen)、抗CD11b-PE單抗(M1/70克隆,BD Pharmingen),抗 CD45R/B220-PerCP單抗(RA 3-6B2克隆,BD Pharmingen)、抗 MHCII-PE單抗(M5/115.15.2克隆,BD Pharmingen),抗CD80-PE單抗(16.10A 1克隆,BD Pharmingen)。
1.3 流式細胞儀檢測脾臟DCs亞群的數(shù)量 無菌取出感染前和感染后第3、5、8d小鼠脾臟,常規(guī)方法制備脾細胞懸液,用0.17mo l/L NH4Cl裂解紅細胞。用含10%胎牛血清(FCS)的 RPMI1640調(diào)整脾細胞終濃度為 1×107/m L。每份樣品用抗CD11c-FITC單抗、抗CD11b-PE單抗和抗CD45R/B220-PerCP單抗進行三色分析,另設(shè)陰性對照管,在預(yù)先加入FcγⅢ/Ⅱ封閉抗體的流式細胞儀專用染色管中加入脾細胞懸液0.1m L,再加入抗CD11c-FITC單抗、抗CD11b-PE單抗和抗CD45R/B220-PerCP單抗進行表面染色,離心去上清后,用0.5m L PBS重懸浮細胞,流式細胞儀進行檢測。
1.4 流式細胞儀檢測脾臟DCs表面MHCⅡ類分子和共刺激分子CD80的表達水平 每份樣品用抗-CD11c-FITC單克隆抗體和抗-MHCⅡ-PE單克隆抗體或抗-CD80-PE單克隆抗體進行雙色分析,另設(shè)陰性對照管。在預(yù)先加入FcγⅢ/Ⅱ封閉抗體的流式細胞儀專用染色管中加入脾細胞懸液0.1m L,再加入抗-CD11c-FITC單克隆抗體和抗-MHCII-PE單克隆抗體或抗-CD80-PE單克隆抗體。離心去上清后,用0.5m L PBS重懸浮細胞,流式細胞儀進行檢測。
1.5 統(tǒng)計學(xué)處理 應(yīng)用SPSS11.5統(tǒng)計學(xué)分析軟件,單因素方差分析比較各組均值的顯著性差異,P<0.05為有統(tǒng)計學(xué)意義(結(jié)果為 3次結(jié)果的平均值)。
2 結(jié) 果
2.1 2種小鼠感染后不同時間的蟲體血癥水平及生存率 感染后第4d,兩種小鼠外周血中均出現(xiàn)瘧原蟲感染的紅細胞,且感染后前9d,原蟲血癥水平呈相同的上升趨勢。BA LB/c小鼠原蟲血癥水平至感染后9d達峰值后迅速下降,并于感染后21d左右自愈,其生存率100%。相比而言,DBA/2小鼠的紅細胞感染率至感染后9d達峰值后也迅速下降,70%~80%的小鼠死亡,幸存小鼠不能清除感染,有低水平的蟲體血癥(見圖1)。

圖1 DBA/2與BALB/c小鼠感染后不同時間蟲體血癥水平及生存率Fig.1 The parasitem ia and survival rate of DBA/2 and BALB/cmice at dif ferent time points after infection
2.2 2 種小鼠感染后不同時間脾臟DCs亞群的數(shù)量 BALB/c和DBA/2小鼠脾臟CD11c+CD11b+DCs的數(shù)量感染后3-5d均明顯升高(P<0.05),并于感染后第8d達到最高水平(P<0.01),在感染前和感染后第 3d,5d和 8d,BALB/c小鼠脾臟CD11c+CD11b+DCs的數(shù)量均低于 DBA/2小鼠,但僅在感染后第8d有統(tǒng)計學(xué)意義(P<0.01)(圖2A)。
與此相反,兩種小鼠脾臟CD11c+CD45R/B220+DCs的數(shù)量在感染后3~8d,盡管也呈逐漸增高趨勢(P<0.05或 P<0.01),但 BA LB/c小鼠CD11c+CD45R/B220+DCs的數(shù)量在各檢測時間點均高于DBA/2小鼠,但僅在感染后第8d有顯著意義(P<0.01)(圖2B)。
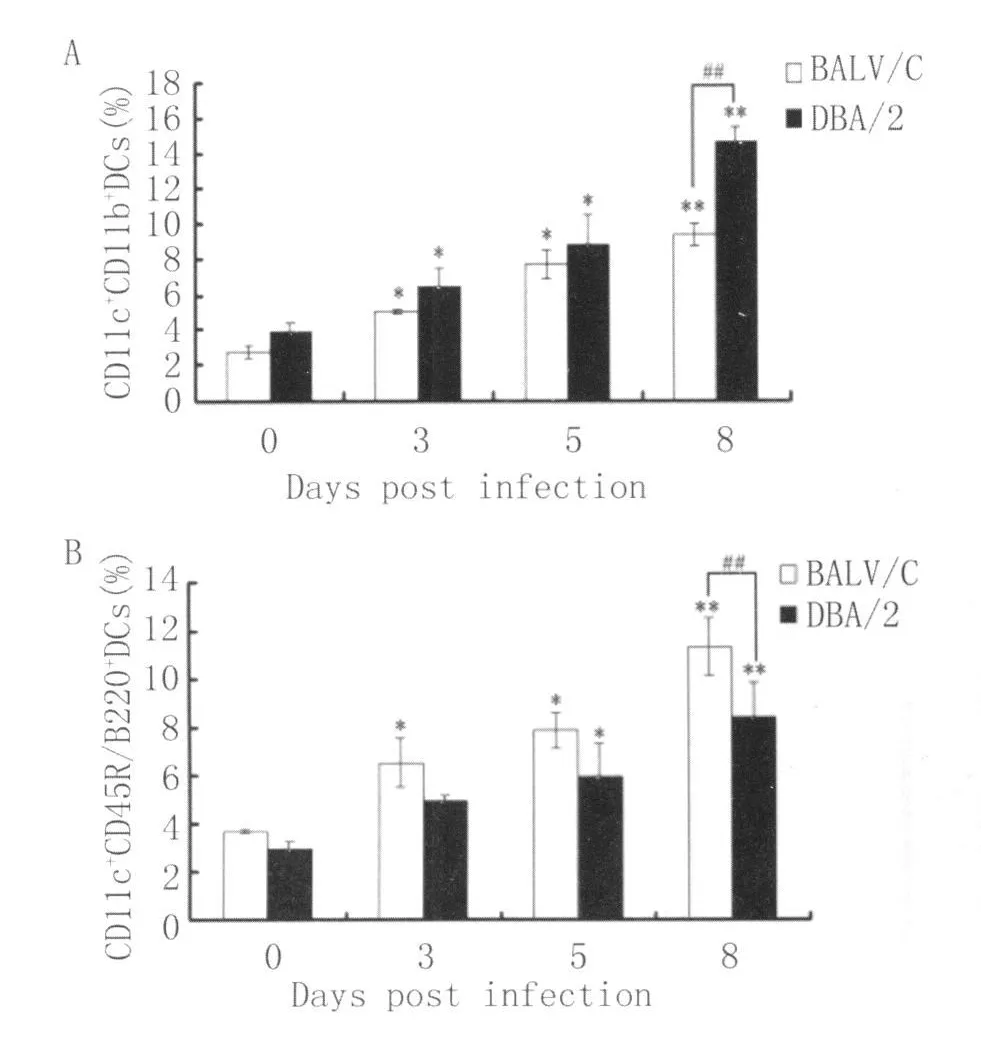
圖2 DBA/2和BALB/c小鼠感染后不同時間脾臟CD11c+CD11b+DCs(A)和CD11c+CD45R/B220+DCs(B)的數(shù)量P<0.05和**P<0.01表示與正常對照組相比差異顯著,##P<0.01表示 DBA/2與 BALB/c小鼠之間的比較Fig.2 Percentage of splenic CD11c+CD11b+DCs(A)and CD11c+CD45R/B220+DCs(B)from DBA/2 and BALB/cmiceatdifferent time points after infection;*P<0.05 and**P<0.01 indicate the significant difference com pared with controlmice(non-infected mice,0d),##P<0.01 indicates the comparison between DBA/2 and BALB/c mice
2.3 2種小鼠感染后不同時間脾臟DCs表面MHCⅡ類分子和CD80分子的表達水平BALB/c和DBA/2小鼠表達MHCⅡ類分子CD11c+DCs的數(shù)量在感染后3-5d明顯升高(P<0.05),并于感染后第8d達到最高水平(P<0.01),在感染前和感染后第3d,5d,8d,BALB/c小鼠表達MHCⅡ類分子的CD11c+DCs的數(shù)量均低于DBA/2小鼠,其中感染后第8d有顯著差異(P<0.01)(圖3A)。與此同時,DBA/2小鼠表達CD80分子的CD11c+DCs的數(shù)量在感染后3-5d顯著升高(P<0.05),并于感染后第 8d達到峰值水平(P<0.01)。而BA LB/c小鼠表達CD80分子的CD11c+DCs的數(shù)量僅從感染后第5d明顯升高(P<0.05),并于感染后第8d達到峰值水平(P<0.01),在感染前和感染后各檢測時間點,BALB/c小鼠表達MHCⅡ類分子的CD11c+DCs的數(shù)量均低于DBA/2小鼠,但僅在感染后第8d有統(tǒng)計學(xué)差異(P<0.01)(圖3B)。

圖3 DBA/2和BALB/c小鼠感染后不同時間脾臟DCs表面MHCⅡ類分子(A)和CD80分子(B)的表達水平。*P<0.05和**P<0.01表示與正常對照組相比差異顯著,##P<0.01表示 BA LB/c與DBA/2小鼠之間的比較Fig.3 Percentage of splenic CD11c+DCs expressing MHCⅡ(A)and CD80(B)from DBA/2 and BALB/cmice at different time points after infection.*P<0.05 and**P<0.01 indicate the significant difference com pared with controlmice(non-infectedmice,0d),##P<0.01 indicates the comparison between DBA/2 and BALB/c mice
3 討 論
在P.c.AS感染鼠瘧模型中,針對紅內(nèi)期瘧原蟲的免疫應(yīng)答已經(jīng)得到解釋,在此模型中,最初的適應(yīng)性免疫應(yīng)答要求CD4+Th1細胞活化效應(yīng)細胞(如巨噬細胞),隨后進一步轉(zhuǎn)化為Th2細胞因子和抗體的產(chǎn)生,以徹底清除寄生蟲。根據(jù)宿主的基因背景,P.c.AS感染鼠導(dǎo)致非致死型和致死型感染,抵抗鼠要求完整的T細胞,巨噬細胞及B細胞功能以最終清除瘧原蟲。根據(jù)蟲血癥水平,易感鼠與抵抗鼠感染過程相似。易感鼠沒有產(chǎn)生特異性抗體,B細胞和抗體對解除紅內(nèi)期感染很重要,缺乏B細胞的鼠不會清除P.yoelii和P.c.chabaudi感染[7]。P.c.AS感染早期,易感DBA/2小鼠和抵抗BALB/c小鼠均產(chǎn)生以IFN-γ分泌增加為主的Th1型細胞免疫應(yīng)答[5],過強的炎癥反應(yīng)可能引起宿主的病理損傷。有研究表明,P.c.AS感染的DBA/2小鼠死亡與免疫病變相關(guān)[6],而目前關(guān)于P.c.AS感染早期易感與抵抗小鼠Th1細胞免疫應(yīng)答差異調(diào)控相關(guān)機制尚未充分闡明。
有研究顯示,DCs在啟動免疫應(yīng)答過程中發(fā)揮關(guān)鍵作用,是固有免疫應(yīng)答向適應(yīng)性免疫應(yīng)答過渡的橋梁,具有活化初始T細胞以啟動初期免疫應(yīng)答的獨特能力[8-9]。在病原微生物的刺激下,不同DCs亞群可呈現(xiàn)不同的增殖水平[3]。瘧原蟲感染期間,髓樣DCs主要介導(dǎo)Th1應(yīng)答[10-12],漿樣DCs對漿細胞的產(chǎn)生和抗體應(yīng)答起關(guān)鍵作用,髓樣DCs/漿樣DCs比值的增加與Th1路徑的活化相關(guān),而其比值降低則有利于Th2路徑的活化[10]。本實驗結(jié)果顯示,P.c.AS感染后3~8d,兩種小鼠髓樣DCs的數(shù)量均出現(xiàn)有意義的升高,但在感染后第 8d,BALB/c小鼠髓樣DCs的數(shù)量明顯低于DBA/2小鼠。同樣,漿樣DCs的產(chǎn)生模式在兩種小鼠體內(nèi)也存在顯著差異,感染后3~8d,兩種小鼠漿樣DCs的數(shù)量盡管也呈逐漸增高趨勢,但BA LB/c小鼠漿樣DCs的數(shù)量在第8d顯著高于DBA/2小鼠。上述結(jié)果充分提示,P.c.chabaudi(AS)感染早期,DCs亞群的不同分化模式可能參與易感和抵抗宿主Th1細胞免疫應(yīng)答的誘導(dǎo)和調(diào)控。
成熟DCs與初始T細胞的活化以及隨后的適應(yīng)性免疫應(yīng)答的誘導(dǎo)密切相關(guān)[9,13]。MHCⅡ類分子和共刺激分子的上調(diào)是 DCs成熟的突出特征[2,14-16]。本實驗結(jié)果顯示,在感染后3~8d,兩種小鼠表達MHCⅡ類分子和CD 80分子的DCs的數(shù)量均呈現(xiàn)逐漸增高的趨勢,但BALB/c小鼠兩種分子的升高幅度均明顯低于DBA/2小鼠。以上結(jié)果提示,DCs成熟的幅度可能決定著Th1細胞免疫應(yīng)答的強度。
綜上,在 P.c.chabaudi(AS)感染早期,BALB/c和DBA/2小鼠DCs亞群的分化模式、成熟表型特征性分子的表達水平存在顯著差異。而這種差異性可能是導(dǎo)致兩種小鼠呈現(xiàn)不同程度 Th1細胞免疫應(yīng)答的前提條件。
[1]Voisine C,Mastelic B,Sponaas AM,et al.Classical CD 11c+dendritic cells,not plasmacytoid dend ritic cells,induce T cell responses to Plasmod ium chabaudi malaria[J].In t J Parasitol,2010,40(6):711-719.
[2]Stevenson MM,U rban BC.Antigen presentation and dendritic cellbiology in malaria[J].Parasite Immunol,2006,28(1-2):5-14.
[3]Gilliet M,Boonstra A,Patu rel C,et al.The development of m urine plasm acy toid dendritic cellprecu rsors is differentially regu lated by FLT3-ligand and granulocyte/m acrophage colony-stimu lating factor[J].JExp Med,2002,195(7):953-958.
[4]Shortman K,Liu YJ.Mouse and human dendritic cell sub types[J].Nat Rev Immunol,2002,2(3):151-161.
[5]金瑛,郭生玉,馮輝,等.CD4+CD25+T細胞在夏氏瘧原蟲感染鼠中的應(yīng)答差異[J].微生物學(xué)雜志,2007,27(2):33-36.
[6]Seixas E,O liveira P,Moura Nunes JF,et al.An experimen tal m odel for fatalmalaria due to TNF-alpha-dependent hepatic damage[J].Parasitology,2008,135(6):683-690.
[7]Seixas E,Ostler D.P lasmod ium chabaudi chabaud i(AS):differential cellular responses to infection in resistant and susceptib lemice[J].Exp Parasitol,2005,110(4):394-405.
[8]Zheng W,Wang QH,Feng H,et al.CD4+CD25+Foxp3+regu latory T cells prevent the development of Th1 immune response by inhibition of DC function du ring the early stage of P lasmod ium yoelii infection in susceptible BALB/cmice[J].Folia Parasitol(Praha),2009,56(4):242-250.
[9]Banchereau J,Briere F,Caux C,et al.Immunobiology of dend ritic cells[J].Annu Rev Immunol,2000,18:767-811.
[10]Jangpatarapongsa K,Chootong P,Sattabongkot J,et al.P lasmod ium vivax parasites alter the balance ofmyeloid and plasmacy toid dendritic cellsand the induction of regu latory T cells[J].Eur JImm unol,2008,38(10):2697-2705.
[11]Kuwana M.Induction of anergic and regu latory T cellsby plasmacytoid dendritic cells and other dend ritic cell subsets[J].Hum Immunol,2002,63(12):1156-1163.
[12]Liu YJ.Dendritic cell subsets and lineages,and their functions in innate and adaptive immunity[J].Cell,2001,106(3):259-262.
[13]Banchereau J,Steinman RM.Dendritic cells and the control of immunity[J].Nature,1998,392(6673):245-252.
[14]Ing R,Segu ra M,Thaw ani N,et al.Interaction ofmouse dendritic cells and m alaria-in fected eryth rocytes:uptake,m atu ration,and antigen presen tation[J].J Immunol,2006,176(1):441-450.
[15]Ing R,Stevenson MM.Dendritic cell and NK cell reciprocal cross talk prom otes gamma interferon-dependent immunity to b lood-stage P lasmodium chabaud i AS infection inmice[J].Infect Immun,2009,77(2):770-782.
[16]W ykes MN,Liu XQ,Beattie L,et al.Plasmod ium strain determines dendritic cell function essen tial for survival from malaria[J].PLoSPathog,2007,3(7):e96.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
美與時代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年12期)2019-05-21 02:55:32
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
云南中醫(yī)學(xué)院學(xué)報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
七彩語文·畫刊(2012年4期)2012-04-29 00:00:00
