微生物及酶固定二氧化碳的研究進(jìn)展
2010-06-04 09:15:06彭艷麗,趙華章,楊親正等
化學(xué)與生物工程 2010年7期
關(guān)鍵詞:體系
溫室效應(yīng)是指人類向大氣中排放CO2等氣體阻礙了地球的熱量擴(kuò)散到外層空間,從而使氣溫升高的現(xiàn)象。溫室效應(yīng)會導(dǎo)致高緯度地區(qū)降雨量增加、高山積雪融化、兩極冰山融化、海平面上升,已成為影響地球生態(tài)環(huán)境的重要原因。溫室效應(yīng)氣體有CO2、氯氟代烷、CH4等30多種,其中CO2影響最大。據(jù)聯(lián)合國氣候變化專門委員會(IPCC)分析[1],全世界約10%~15%的廢氣來自于煤燃燒排放的CO2,每年約200億 t,溫室效應(yīng)將導(dǎo)致地球的平均溫度上升1.5~4.5℃。除減少排放外,還原和固定CO2也將有助于減輕溫室效應(yīng)。CO2的固定和利用方法主要有物理法、化學(xué)法和生物法。物理法和化學(xué)法雖然可以制得新的有用化合物,但會產(chǎn)生一些副產(chǎn)物,而且消耗能量,對環(huán)境造成二次污染。生物法固定CO2是通過植物或微生物的循環(huán)途徑將CO2轉(zhuǎn)化成化學(xué)物質(zhì)或其自身生長的營養(yǎng)物質(zhì)。固定CO2的生物主要是植物和自養(yǎng)微生物。綠色植物的葉綠體中有一個特有的酶促機(jī)構(gòu),催化CO2轉(zhuǎn)變成還原性的有機(jī)化合物,這個過程為CO2的固定過程。但是地球上還有植物不能生長的特殊環(huán)境,自養(yǎng)微生物固定CO2的優(yōu)勢便顯現(xiàn)出來。從整個生物圈的物質(zhì)、能量流來看,自養(yǎng)微生物可以利用可見光作為主要能源驅(qū)動CO2的固定。使用特定的酶可更加直接快速地實(shí)現(xiàn)CO2的固定。作者在此介紹了近年來微生物和酶固定CO2的研究進(jìn)展。
1 微生物固定CO2的研究進(jìn)展
1.1 固定CO2的微生物種類
自養(yǎng)微生物利用光能或無機(jī)物氧化時產(chǎn)生的化學(xué)能同化CO2,構(gòu)成細(xì)胞物質(zhì)。固定CO2的自養(yǎng)微生物較多[2],一般有兩類:光能自養(yǎng)型和化能自養(yǎng)型微生物,如表1所示。
表1 固定CO2的微生物種類
Tab.1Microbialtypesofcarbondioxideimmobilization
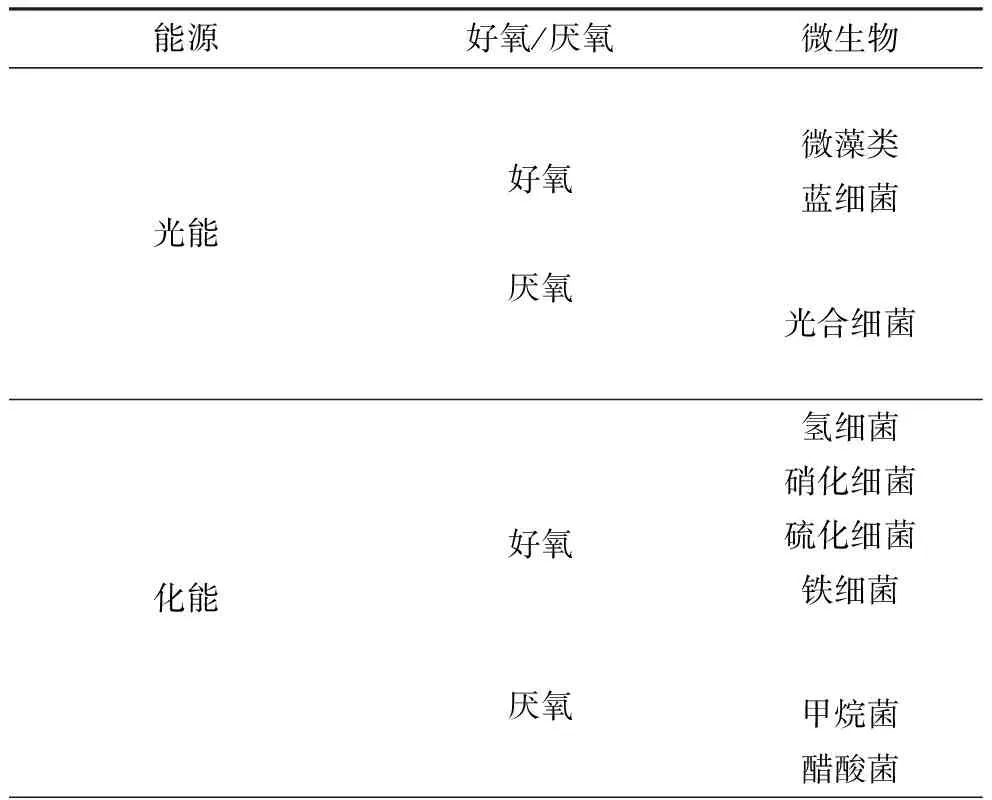
能源好氧/厭氧微生物光能好氧厭氧微藻類藍(lán)細(xì)菌光合細(xì)菌化能好氧厭氧氫細(xì)菌硝化細(xì)菌硫化細(xì)菌鐵細(xì)菌甲烷菌醋酸菌
光能自養(yǎng)型微生物包括微藻類、藍(lán)細(xì)菌和光合細(xì)菌,含有葉綠素,以光為能源、CO2為碳源合成菌體物質(zhì)或代謝產(chǎn)物;化能自養(yǎng)型微生物以CO2為碳源,能源主要有H2、H2S、NH4+、NO2-、Fe2+等。微藻(包括藍(lán)細(xì)菌)和氫細(xì)菌具有生長速度快、適應(yīng)性強(qiáng)等特點(diǎn)。國內(nèi)外現(xiàn)已大規(guī)模生產(chǎn)的微藻主要有:柵列藻(Scenedesmus)、小球藻(Chlorella)、鹽藻(Dunaliella)和螺旋藻(Spirulina)等。Ishizaki等[3]利用產(chǎn)堿桿菌以CO2為碳源在限氧條件閉路循環(huán)發(fā)酵系統(tǒng)中培養(yǎng)60 h,其菌體濃度高于60 g·L-1、PHB達(dá)36 g·L-1。化能自養(yǎng)菌中固定CO2生長速度最快的是氫細(xì)菌,已發(fā)現(xiàn)的氫細(xì)菌有18個屬,近40個種。Nishihara[4]篩選的中溫氫細(xì)菌在固定CO2時還分別積累大量的胞外多糖和胞內(nèi)糖原型多糖。
1.2 微生物固定CO2
微藻類具有光合速率快、繁殖快、環(huán)境適應(yīng)性強(qiáng)、固定效率高等優(yōu)點(diǎn),常與其它工程技術(shù)結(jié)合用于固定CO2。de Morais等[1]將螺旋藻中的Spirulinasp.和斜生柵藻S.obliquus培養(yǎng)在恒溫30℃的三級連續(xù)管狀光生物反應(yīng)器中,觀察CO2的固定狀況和這兩種微生物對CO2的耐受力。結(jié)果發(fā)現(xiàn),螺旋藻Spirulinasp.的最大比生長速率和最高產(chǎn)率分別為0.44 d-1和0.22 g·L-1·d-1;最高CO2固定率在CO2體積分?jǐn)?shù)為6%時是53.29%,在CO2體積分?jǐn)?shù)為12%時為45.61%。斜生柵藻S.obliquus的最高CO2固定率在CO2體積分?jǐn)?shù)為6%時為28.08%,在CO2體積分?jǐn)?shù)為12%時為13.56%。
Hanagata等[5]培養(yǎng)的Chlorella在CO2體積分?jǐn)?shù)為10%時,產(chǎn)率為0.15 g·L-1·d-1,在CO2體積分?jǐn)?shù)為40%時,產(chǎn)率為0.18 g·L-1·d-1。Sung等[6]培養(yǎng)的Chlorella菌株KR-1在不同體積分?jǐn)?shù)CO2下有不同的產(chǎn)率,CO2體積分?jǐn)?shù)為10%時產(chǎn)率為1.1 g·L-1·d-1;CO2體積分?jǐn)?shù)為30%時產(chǎn)率為0.8 g·L-1·d-1;CO2體積分?jǐn)?shù)為50%時產(chǎn)率為0.6 g·L-1·d-1,CO2體積分?jǐn)?shù)為70%時產(chǎn)率為0.1 g·L-1·d-1。有的菌株在CO2環(huán)境中可以加速生長,Chang等[7]培養(yǎng)的Chlorella菌株NTU-H15和NTU-H25在通入5%CO2時細(xì)胞生長速度加快,最高產(chǎn)率達(dá)到0.31 g·L-1·d-1。
隨著基因工程的發(fā)展,固定CO2細(xì)菌和基因工程有了契合點(diǎn)。杜翠紅[8]以沼澤紅假單胞菌為受體菌株,以卡爾文循環(huán)中的關(guān)鍵酶核酮糖-1,5-二磷酸羧化酶/氧化酶(RubisCO)的基因?yàn)槟康幕颍柚訚杉t假單胞菌/大腸桿菌穿梭載體,將目的基因轉(zhuǎn)入受體菌株中,構(gòu)建了一株固定CO2的基因工程菌。
研究人員在研究CO2還原產(chǎn)物的同時,發(fā)現(xiàn)CO2被固定后的某些產(chǎn)物還可以作為中間物質(zhì)維持還原反應(yīng)的循環(huán)進(jìn)行。Ishizaki等[9]和Tanaka[10]利用AlcaligeneseutrophusATCC 17697T固定CO2生產(chǎn)聚-3-羥基丁酸酯。辛嘉英[11]利用甲基彎菌IMV3011催化CO2生物轉(zhuǎn)化生成甲醇,在細(xì)胞懸浮液中充入CO2反應(yīng)一段時間后,在反應(yīng)液中檢測到甲醇。還發(fā)現(xiàn)細(xì)胞內(nèi)貯存的聚-β-羥基丁酸分解后能夠產(chǎn)生還原當(dāng)量,通過改變培養(yǎng)基中氮和銅的起始濃度對PHB積累量進(jìn)行調(diào)節(jié),可提高甲基彎菌IMV3011還原CO2生成甲醇的能力。
有研究人員進(jìn)行了自養(yǎng)固定CO2細(xì)菌的研究。Berg等[12]發(fā)現(xiàn)古細(xì)菌屬硫化葉菌目的自養(yǎng)型細(xì)菌Metallosphaerasedula可以利用乙酰輔酶A或丙酰輔酶A羧化酶固定CO2。Ramos-Vera等[13]報道了兩種新的自養(yǎng)型細(xì)菌固定CO2的循環(huán)。硫化葉菌目利用3-羥基丙酸鹽/4-羥基丁酸鹽循環(huán),以乙酰輔酶A/輔酶A羧化酶作為催化酶。Ignicoccus hospitalis(除硫球菌目)利用二甲酸亞乙基酯/4-羥基丁酸鹽循環(huán),以丙酮酸合酶和磷酸烯醇丙酮酸羧激酶作為催化酶固定CO2。在這兩個循環(huán)中,乙酰輔酶A和兩個無機(jī)碳通過不同的途徑被轉(zhuǎn)化成琥珀酰-輔酶A,而從琥珀酰-輔酶A到乙酰輔酶A的再生途徑是相同的,該循環(huán)是嚴(yán)格厭氧環(huán)境。
近年來,電化學(xué)的研究引起了人們的關(guān)注。Cao等[14]在MFC中利用生物陰極,用溶解的CO2作為電子受體,利用外加電源提供0.242 V的電勢(相對于SHE)。經(jīng)過一個月的調(diào)整最終得到1 mA的穩(wěn)定電流,每1 mol e-還原(0.28 ±0.02) mol重碳酸鹽,這是首次利用陰極和微生物間直接電子轉(zhuǎn)移固定CO2。
2 酶法固定CO2的研究進(jìn)展
微生物法固定CO2實(shí)質(zhì)上是微生物中的某些酶在生物體內(nèi)一系列的催化反應(yīng)。早在1984年就有人提出酶法還原CO2,即以酶為催化劑通過一連串酶促反應(yīng),將CO2轉(zhuǎn)化為有用物質(zhì)。目前研究還原CO2較多的是多酶體系,包括三種脫氫酶:甲酸脫氫酶(FDH)、甲醛脫氫酶(FADH)、甲醇脫氫酶(MDH),這三種酶分別可以將甲酸、甲醛、甲醇氧化成CO2。多酶系統(tǒng)固定CO2是向該系統(tǒng)中供給逆反應(yīng)進(jìn)行方向的物質(zhì),使得CO2被還原。
Parkinson等[15]于1984年提出了酶固定CO2的途徑,以甲酸脫氫酶作催化劑將CO2還原為甲酸。Susumu等[16]以甲酸脫氫酶為催化劑,以甲基紫精或PQQ作為電子傳遞體,以飽和CO2磷酸緩沖溶液(pH=7)為電解液,在-0.7~-0.9 V(相比于SCE飽和甘汞電極)電極電勢下將溶解的CO2轉(zhuǎn)化成甲酸鹽,轉(zhuǎn)化率高達(dá)90%,甲酸脫氫酶在暗處的酶活力較為持久。在以甲醇脫氫酶為催化劑、以甲基紫精為電子傳遞體、以甲酸鈉飽和磷酸鹽溶液為電解液、電極電勢為-0.7~-0.9 V(相比于SCE飽和甘汞電極)的條件下,酶濃度低時生成甲醛;酶濃度高時,甲醛積累到一定程度生成甲醇;當(dāng)PQQ代替甲基紫精作為電子中間體時只產(chǎn)生甲醇。在以上結(jié)果的基礎(chǔ)上,以甲酸脫氫酶和甲醇脫氫酶共同作為催化劑,以PQQ作為電子中間體可將CO2還原成為甲醇。
生物體內(nèi)的酶催化化學(xué)反應(yīng)是連續(xù)的,前一種酶的作用產(chǎn)物往往是后一種酶的作用底物。Obert等[17]采用多酶體系將CO2還原成甲醇。CO2先在甲酸脫氫酶作用下還原為甲酸鹽,然后在甲醛脫氫酶作用下還原為甲醛,最后在甲醇脫氫酶作用下還原為甲醇,每一步脫氫酶反應(yīng)的最終電子供體都為還原性NADH。
天然酶穩(wěn)定性差、易失活、不能重復(fù)使用,為了保證多酶體系作用連續(xù)有效,用溶膠-凝膠多孔納米材料將多酶體系包埋起來。結(jié)果表明,同多酶體系直接在溶液中還原CO2相比,包埋于多孔溶膠-凝膠中的多酶體系還原CO2為甲醇的產(chǎn)率明顯提高。
姜忠義等[18]采用改進(jìn)的溶膠-凝膠法固定多酶體系,以NADH為電子供體,在低溫低壓條件下將CO2還原成甲醇,通過改變反應(yīng)物配比、催化劑用量及其它反應(yīng)條件,摸索出較適宜的凝膠化條件為:37℃、pH=7.0,在此條件下甲醇產(chǎn)率達(dá)92.4%。過程如下:
上述反應(yīng)體系需加入昂貴的NADH電子供體,且不能連續(xù)反應(yīng)。
崔俊儒等[19]發(fā)現(xiàn)含多酶體系的甲烷氧化細(xì)菌在溫和條件下經(jīng)過一系列反應(yīng)能夠?qū)O2還原成甲醇,CO2還原而成的甲醇在細(xì)胞外得以積累。在反應(yīng)體系中加入甲烷作為底物時,產(chǎn)生的還原能量推動反應(yīng)連續(xù)穩(wěn)定進(jìn)行。
為優(yōu)化反應(yīng)條件,Sun等[20]將三種脫氫酶通過溫和的仿生礦化過程包埋于二氧化鈦粒子體系中,用于還原CO2成甲醇。包埋后的多酶體系在NADH作為電子供體的條件下依次將CO2還原為甲酸、甲醛、甲醇。和開放型反應(yīng)系統(tǒng)相比,包埋后的多酶體系將CO2還原為甲醇的產(chǎn)率有了很大的提高。
Reda等[21]以甲基紫精為電子中間體,在嚴(yán)格厭氧環(huán)境中將甲酸脫氫酶吸附到電極表面用于電化學(xué)還原CO2生成甲酸鹽。甲酸脫氫酶電催化在熱力學(xué)上是可逆的,且只需要很低的過電壓,此反應(yīng)發(fā)生在溫和條件下,甲酸鹽是唯一產(chǎn)物。
上述研究都是使用多酶體系來模擬生物體的光合作用而還原CO2,反應(yīng)無需光照,且不添加光催化劑或光敏劑。在酶固定轉(zhuǎn)化CO2的研究中,還有使用光敏劑對CO2進(jìn)行光催化還原的報道。如Willner等[22]在上述體系中添加了光敏劑釕,利用酶ICDH和電子傳輸介質(zhì)MV2+成功進(jìn)行了CO2的光化學(xué)固定及還原。
3 展望
微生物及酶固定CO2的研究取得了一些進(jìn)展,目前有望實(shí)現(xiàn)大規(guī)模工業(yè)生產(chǎn)的是微藻類和氫細(xì)菌。微藻類在自身繁殖菌體時,可將CO2轉(zhuǎn)化為生物柴油等液體燃料或者利用CO2生產(chǎn)有用物質(zhì),如:類脂和蛋白質(zhì)等;氫細(xì)菌在固定CO2的同時還能積累大量胞內(nèi)糖原。通過相應(yīng)的酶作用可以將CO2還原為蘋果酸和異檸檬酸、甲烷、甲醇、乙醇等。微生物固定CO2仍有一些瓶頸問題,如細(xì)胞生長速度慢、密度低,可用的菌種資源有限,催化氧化過程需要還原性輔酶參與。酶法固定CO2存在酶種類少、途徑單一、轉(zhuǎn)化效率低等缺點(diǎn)。微生物和酶固定CO2的機(jī)理也較復(fù)雜,還需要不斷的探索。
微生物和酶固定CO2今后的研究方向主要是:(1) 開發(fā)光生物反應(yīng)器以滿足微藻類生長所需的光強(qiáng),實(shí)現(xiàn)CO2在溫和條件下轉(zhuǎn)化為有機(jī)碳;(2) 通過基因工程構(gòu)建固定CO2的高效菌株;(3) 進(jìn)一步研究微生物和酶固定CO2的機(jī)理,為CO2固定研究提供理論支持;(4)開發(fā)新的高效分離技術(shù)以分離還原產(chǎn)物;(5)同其它還原CO2的方式(如光催化和電化學(xué)還原等)結(jié)合,用于合成各種化學(xué)品及高分子材料。
參考文獻(xiàn):
[1] de Morais Michele Greque,Costa Jorge Alberto Vieira.Biofixation of carbon dioxide bySpirulinasp.andScenedesmusobliquuscultivated in a three-stage serial tubular photobioreactor [J].Journal of Biotechnology,2007,12(9):439-445.
[2] Hiroyuki H.Effective lactic acid production by two-stage extractive fermentation[J].Ferment Bioeng,1995,79(6):589-593.
[3] Ishizaki A,Tanaka K.Batch culture ofAlcaligeneseutrophus
ATCC 17697T using recycled gas closed circuit culture system[J].Ferment Bioeng,1990,69(3):170-174.
[4] Nishihara H.Production and properties of glycogen in the marine obligate chemolithoautotroph,Hydrogenovibriomarinus[J].Ferment Bioeng,1993,75(6):414-416.
[5] Hanagata N,Takeuchi T,Fukuju Y,et al.Tolerance of microalgae to high CO2and high temperature[J].Phytochemistry,1992,31(10):3345-3348.
[6] Sung K D,Lee J S,Shin C S,et al.CO2fixation byChlorellasp.KR-1 and its cultural characteristics[J].Biores Technol,1999,68(3):269-273.
[7] Chang E H,Yang S S.Some characteristics of microalgae isolated in Taiwan for biofixation of carbon dioxide[J].Bot Bull Acad Sin,2003,44(1):43-52.
[8] 杜翠紅.沼澤紅假單胞菌RubisCO基因的克隆與表達(dá)及其固定二氧化碳特性的研究[D].大連:大連理工大學(xué),2003.
[9] Ishizaki A,Taga N,Takeshita,et al.Microbial production of biodegradable plastics from carbon dioxide and agricultural waste material[J].Ferment Bioeng,1991,71(4):254-257.
[10] Tanaka K.Production of poly-d-3-hydroxybutyric acid from carbon dioxide by a two-stage culture method employingAlcaligeneseutrophusATCC 17697T[J].Ferment Bioeng,1994,77(4):425-427.
[11] 辛嘉英.甲基彎菌IMV3011細(xì)胞生物催化二氧化碳制甲醇[J].分子催化,2008,22(4):356-361.
[12] Berg Ivan A,Kockelkorn Daniel,Buckel Wolfgang,et al.A
3-hydroxypropionate/4-hydroxybutyrate autotrophic carbon dioxide assimilation pathway inArchaea[J].Science,2007,318(5857):1782-1786.
[13] Ramos-Vera W H,Berg I A,Fuchs G.Autotrophic carbon dioxide assimilation in thermoproteales revisited[J].Journal of Bacteriology,2009,191(13):4286-4297.
[14] Cao Xiaoxin,Huang Xia,Liang Peng,et al.A completely anoxic microbial fuel cell using a photo-biocathode for cathodic carbon dioxide reduction[J].Energy Environ Sci,2009,2(5):498-501.
[15] Parkinson B A,Weaver Paul F.Photoelectrochemical pumping of enzymatic CO2reduction[J].Nature,1984,309:148-149.
[16] Susumu Kuwabata,Tsuda Ryo,Yoneyama Hiroshi.Electrochemical conversion of carbon dioxide to methanol with the assistance of formate dehydrogenase and methanol dehydrogenase as biocatalysts[J]. J Am Chem Soc,1994,116(12):5437-5443.
[17] Obert Robyn,Dave Bakul C.Enzymatic conversion of carbon dioxide to methanol:Enhanced methanol production in silica sol-gel matrices[J].J Am Chem Soc,1999,121(51):12192-12193.
[18] 姜忠義,吳洪,許松偉,等.溶膠-凝膠固定化多酶催化二氧化碳轉(zhuǎn)化為甲醇反應(yīng)初探[J].催化學(xué)報,2002,23(2):162-164.
[19] 崔俊儒,辛嘉英,牛建中,等.甲烷氧化細(xì)菌催化二氧化碳生物合成甲醇的研究[J].分子催化,2004,18(3):214-218.
[20] Sun Qianyun,Jiang Yanjun,Jiang Zhongyi,et al.Green and efficient conversion of CO2to methanol by biomimetic coimmobilization of three dehydrogenases in protamine-templated titania[J].Ind Eng Chem Res,2009,48(9):4210-4215.
[21] Reda Torsten,Plugge Caroline M,Abram Nerilie J,et al.Reversible interconversion of carbon dioxide and formate by an electroactive enzyme[J].PNAS,2008,105(31):10654-10658.
[22] Willner I,Mandler D,Rinklin A.Photoreduction of CO2to CH4in aqueous solutions using visible light[J].J AM Chem Soc,1998,108:8100-8101.
猜你喜歡
商品與質(zhì)量(2021年43期)2022-01-18 05:31:22
杭州(2020年23期)2021-01-11 00:54:42
新世紀(jì)智能(數(shù)學(xué)備考)(2020年11期)2021-01-04 00:38:16
中國外匯(2019年17期)2019-11-16 09:31:14
中國衛(wèi)生(2015年12期)2015-11-10 05:13:40
現(xiàn)代企業(yè)(2015年1期)2015-02-28 18:43:18
汽車零部件(2014年5期)2014-11-11 12:24:28
新高考·高一物理(2014年1期)2014-09-18 01:26:07
浙江人大(2014年1期)2014-03-20 16:19:53
終身教育研究(2012年4期)2012-03-25 10:41:11
