基于轉錄組和蛋白質組分析篩選五龍鵝產蛋相關候選基因
2025-01-27 00:00:00魯秀張名愛孔敏張晶王秉翰侯中一滕興怡姜雅靜凡文磊王寶維
畜牧獸醫學報 2025年1期


摘 要: 旨在通過轉錄組和蛋白組聯合分析來挖掘五龍鵝產蛋量相關候選基因,為其產蛋性能遺傳機制研究提供理論基礎。本研究選取200只體重相近的36周齡健康五龍鵝種鵝,記錄其36~54周齡產蛋情況。在產蛋后期(54周齡)時,根據產蛋水平,將產蛋量最高的30只鵝定為高產組,產蛋量最低的30只鵝定為低產組,從高產組(H)和低產組(L)鵝中各采集3個卵巢組織,提取RNA和蛋白質,通過轉錄組測序(RNA-Seq)和4D-DIA蛋白組定量分析篩選差異表達基因(differentially expressed genes,DEGs)和差異表達蛋白(differentially expressed proteins,DEPs)。之后對篩選的差異基因和蛋白進行RT-qPCR驗證、功能富集分析和蛋白互作(PPI)網絡的構建與分析,篩選鵝產蛋性能相關的候選基因。結果:共鑒定到450個DEGs和333個DEPs,其中,AFAP1、INHA、FNDC1、INHBB在基因和蛋白水平上表達出相同的差異趨勢。富集分析顯示,DEGs和DEPs顯著富集到364個GO條目和75個KEGG通路,涉及生殖結構發育(gland development)、性別分化(sex differentiation)、性激素活性(hormone activity)、TGF-β通路(TGF-β signaling pathway)、卵母細胞減數分裂(oocyte meiosis)、GnRH信號通路(GnRH signaling pathway)、孕激素介導的卵母細胞成熟(progesterone-mediated oocyte maturation)等多個與產蛋相關的通路和關鍵生物過程。選取9個潛在候選基因,對高產組和低產組(包含轉錄組測序樣品)各8個卵巢組織進行RT-qPCR驗證,表達趨勢與測序結果一致。基于上述研究結果,本研究最終篩選到6個與五龍鵝產蛋性能密切相關的候選基因(PTTG1、LRP2、TNFSF10、INHA、INHBB、FST),豐富了五龍鵝產蛋性能的相關分子機制。
關鍵詞: 五龍鵝;轉錄組;蛋白組;產蛋量;候選基因
中圖分類號:S835.3""""" 文獻標志碼:A""""" 文章編號: 0366-6964(2025)01-0232-14
收稿日期:2024-07-22
基金項目:國家重點研發計劃(2023YFD1300300);國家水禽產業技術體系(CARS-43-11);山東省重點研發計劃(2021CXGC010805);農業農村部政府購買服務:鵝高繁性狀精準評價和分子鑒定(19230551)
作者簡介:魯 秀(1997-),女,山東新泰人,碩士生,主要從事動物遺傳與育種研究,E-mail: luxiu911@163.com
*通信作者:凡文磊,主要從事動物遺傳與育種方向的研究,E-mail: fanwenlei@qau.edu.cn;王寶維,主要從事動物營養與保健方向的研究,E-mail: wangbw1959@qq.com
Screening for Candidate Genes Related to Egg Production in Wulong Geese Based on Transcriptome
and Proteome Analyses
LU" Xiu ZHANG" Ming’ai 2022年商品鵝出欄量達到4.68億只,肉鵝產值高達526.73億元[1]。然而,在鵝產業快速發展的同時,正在面臨著繁殖性能差的卡脖子問題[2]。因此,深入挖掘影響鵝產蛋量的關鍵基因,對鵝產蛋性狀分子標記輔助育種與鵝產業的可持續發展具有重要意義。
家禽產蛋量受卵巢和卵泡發育水平的影響,而生長因子、激素以及卵泡閉鎖等多個因素均能通過調控卵泡發育水平影響家禽的繁殖性能。利用基因組學技術對揚州鵝、四川白鵝的基因進行測序,發現了大量與產蛋相關的SNPs位點,以及GnRH和GnIH等基因[3],證實這些基因在性腺軸中起關鍵作用,揭示了與產蛋量之間的關聯性。通過轉錄組學技術對山麻鴨[4]、浙東白鵝[5]等產蛋性狀進行研究,篩選出了INHA、GDF9、FST[6]、BMP、EGR[7]等基因,這些基因在卵泡發育和閉鎖過程中發揮重要作用,對于維持繁殖功能的穩定具有重要意義。黃宣等[8]采用蛋白組學技術對高低產雞卵巢進行深度分析后發現,CYP11A1、HSD3B1等蛋白可能與類固醇激素合成過程有關,對卵泡的發育和排卵過程具有重要作用。盡管,目前對家禽產蛋調控機制已經有了一定程度的了解,但仍存在很多問題有待闡明,如具體影響產蛋量的基因和通路,高產和低產鵝在卵巢表達上有何差異等。
豁眼鵝(五龍鵝)是我國產蛋性能最優秀的地方鵝品種之一,年產蛋量可達100枚左右。但是在生產中發現,同一群體中的不同個體,在產蛋水平上存在較大差異,群體內變異較大[9]。因此,本研究通過在同一五龍鵝群體中篩選高產和低產個體,對其卵巢組織進行轉錄組和蛋白組聯合分析,旨在篩選影響五龍鵝產蛋量的關鍵候選基因,為后續利用分子標記輔助育種培育鵝高繁配套系奠定基礎。
1 材料與方法
1.1 試驗動物及樣本采集
選擇200只36周齡體重相近、健康的五龍鵝母鵝為研究對象,試驗群體飼養于青島農業大學優質水禽研究所育種基地,飼養模式為單籠飼養。在36~54周齡期間記錄全部產蛋量,根據產蛋水平分為高產組和低產組,其中產蛋量最高的30只鵝定為高產組,產蛋量最低的30只鵝定為低產組。在產蛋后期(54周齡)屠宰,屠宰前12 h內禁食、自由飲水。屠宰后,采集卵巢組織(去除等級卵泡),用RNase-free水沖洗干凈,吸干表面水分,迅速放入液氮速凍,置于-80 ℃保存備用。每組隨機選取3個卵巢組織,即3個高產組(H)樣品和3個低產組(L)樣品,送至武漢邁特維爾生物科技有限公司,將卵巢樣勻漿后取部分樣品進行RNA和蛋白質提取,用于后續轉錄組測序與蛋白質測序。
1.2 轉錄組測序數據處理
采用Trizol法提取總RNA,利用Qsep400生物分析儀精確檢測RNA完整性,質檢合格后,用帶有Oligo(dT)的磁珠富集mRNA,隨后用fragmentation buffer將RNA打斷成短片段,以此為模板,用六堿基隨機引物和逆轉錄酶合成cDNA第一鏈,并以第一鏈cDNA為模板進行第二鏈cDNA的合成。最后用PCR法富集cDNA,構建鵝卵巢cDNA文庫。構建完成后,使用高通量核酸蛋白分析系統對文庫的insert size進行檢測,完成文庫質檢。采用Illumina平臺測序,使用fastp 0.23.2對原始數據進行處理;利用Hisat2 2.2.1將樣本clean reads與鵝參考基因組進行序列比對。建庫及測序過程均由武漢邁特維爾生物科技有限公司完成。
1.3 蛋白組測序數據處理
取勻漿后的樣品,加入含有1 mmol·L-1 PMSF(苯甲基磺酰氟)和2 mmol·L-1 EDTA(乙二胺四乙酸)以及8 mol·L-1尿素的混合裂解緩沖液中孵育5 min,充分裂解,離心取上清。通過BCA法測定總蛋白濃度后,根據濃度取等量蛋白溶液,用8 mol·L-1尿素將體積補全至200 μL,經10 mmol·L-1 DTT(二硫蘇糖醇)還原、50 mmol·L-1 IAM(碘乙酰胺)烷基化處理后,用預冷丙酮沉淀蛋白。離心后,將蛋白沉淀風干并重懸于碳酸氫銨溶液和胰蛋白酶(Promega)中,37℃消化過夜。消化產物經C 18柱料脫鹽,真空離心濃縮,并溶解于1 mL·L-1甲酸中。此后,使用NanoElute高效液相色譜儀和timsTOF Pro2質譜儀進行分析;使用DIA-NN軟件以Library free方法搜庫,進行質譜解析,完成蛋白定量。
1.4 DEGs、DEPs篩選及功能富集分析
使用String Tie進行新基因預測,使用Feature Counts計算基因在樣本中的表達量。使用DESeq2對基因表達量數據進行標準化處理和差異分析,差異表達基因(differentially expressed genes,DEGs)篩選的標準為P<0.05且log 2FC≥1.0。利用t檢驗對兩組樣本間蛋白表達量進行組間差異顯著性的檢驗,差異表達蛋白(differentially expressed proteins,DEPs)篩選的標準為P<0.05且FC≥1.2或FC≤0.83。合并DEGs和DEPs,并通過在線網站DAVID進行GO功能富集和KEGG信號通路分析。
1.5 熒光定量PCR驗證
在篩選出的與產蛋相關候選基因中隨機選擇出9個DEGs(LRP2、TNFSF10、A2M、SNRPA1、PTTG1、BMP5、FST、PRLHR、INHBB),以GAPDH作為內參基因,并且在高產組和低產組的卵巢組織中進行RT-qPCR驗證。利用NCBI Primer-BLAST (https:∥www.ncbi.nlm.nih.gov/tools/primer-blast/)在線工具設計引物,引物由青島蔚來生物科技有限公司合成。熒光定量PCR試劑盒采購自湖南艾科瑞生物工程有限公司,熒光定量PCR反應體系如下:上、下游引物(表1)各0.4 μL,cDNA模板2 μL,酶Mix 10 μL,ddH 2O 7.2 μL;反應程序為:95℃預變性30 s;95℃變性5 s,60℃退火延伸30 s,40個循環。采用2-ΔΔCt方法計算基因的相對mRNA表達量,并通過t檢驗進行統計分析。
1.6 DEGs、DEPs蛋白互作網絡構建及可視化
利用在線數據庫STRING對與產蛋相關的候選基因和候選蛋白進行檢索,并結合Cytoscape軟件構建蛋白互作網絡,利用Cyto Hubba插件統計網絡中節點(degree)和節點的中介中心性(between centrality),探究互作網絡的結構特征,并進行可視化與分析,進一步篩選產蛋相關的關鍵候選基因。
2 結 果
2.1 高、低產鵝產蛋情況
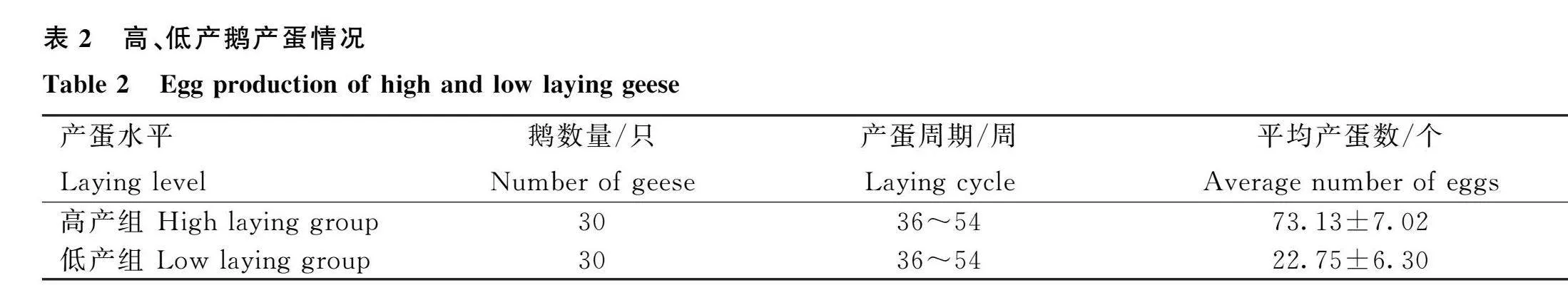
在36~54周齡的產蛋周期內,將200只鵝根據產蛋量進行排序,排名前30的鵝劃分為高產組,排名后30的鵝劃分為低產組,其中高產組平均產蛋量為73個,低產組平均產蛋量為22個(表2)。
2.2 DEGs和DEPs篩選
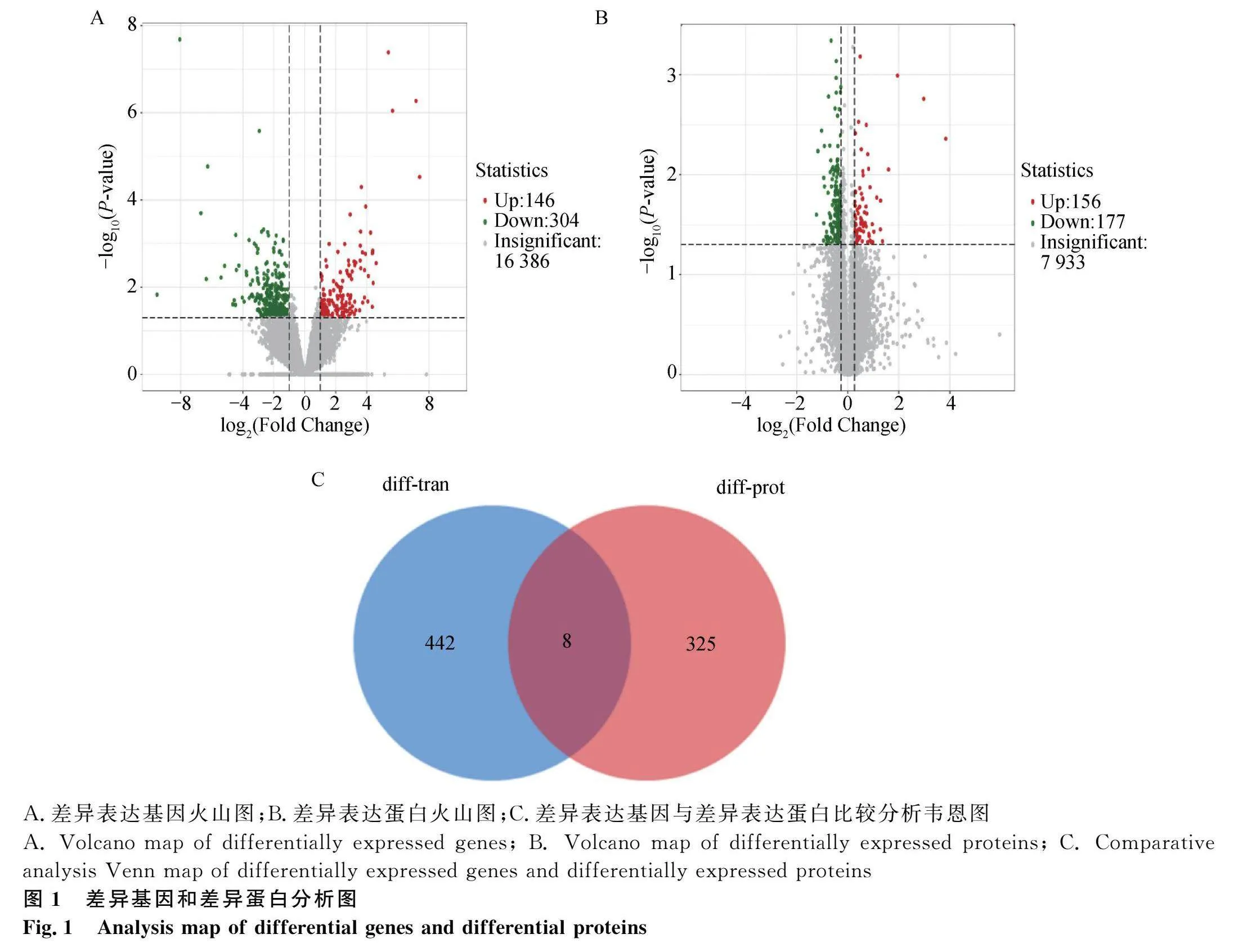
經過對原始轉錄組測序數據質控,平均每個樣品獲得4.37×107條clean reads,平均注釋效率為90.94%,最終獲得23 324個基因的mRNA表達量。利用DESeq2包進行基因表達量差異分析,成功鑒定出450個DEGs,其中有146個基因高產組表達量顯著高于低產組,304個基因高產組表達量顯著低于低產組(圖1A)。
樣本蛋白的質譜分析結果經過定量后獲得8 266個基因的蛋白表達量數據。對兩組樣本的蛋白表達量進行t檢驗,共篩選到333個DEPs,其中156個蛋白的表達量高產組顯著高于低產組,177個蛋白的表達量高產組顯著低于低產組(圖1B)。
此外,在DEGs和DEPs聯合分析中,特別關注到同時在基因和蛋白層面均表現出差異表達的交集基因(圖1C),經過仔細比對,共得到8個基因AFAP1、MSC、CFAP206、MRPL51、INHA、FNDC1、INHBB和CNGB1(表3)。值得注意的是,其中AFAP1、INHA、FNDC1、INHBB這4個基因的mRNA和蛋白表達趨勢完全一致,為后續的研究提供了重要線索。
2.3 DEGs、DEPs的功能富集分析
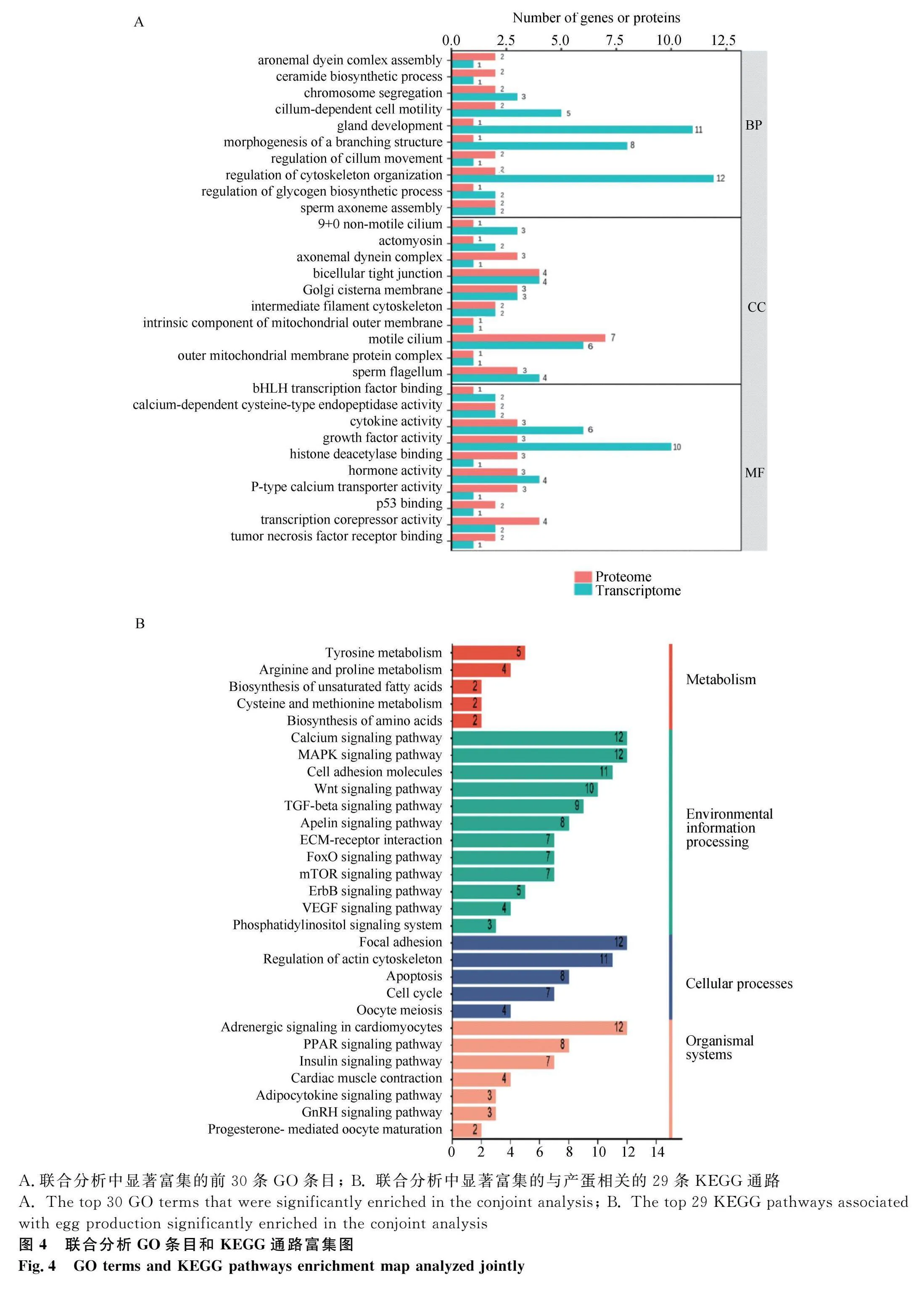
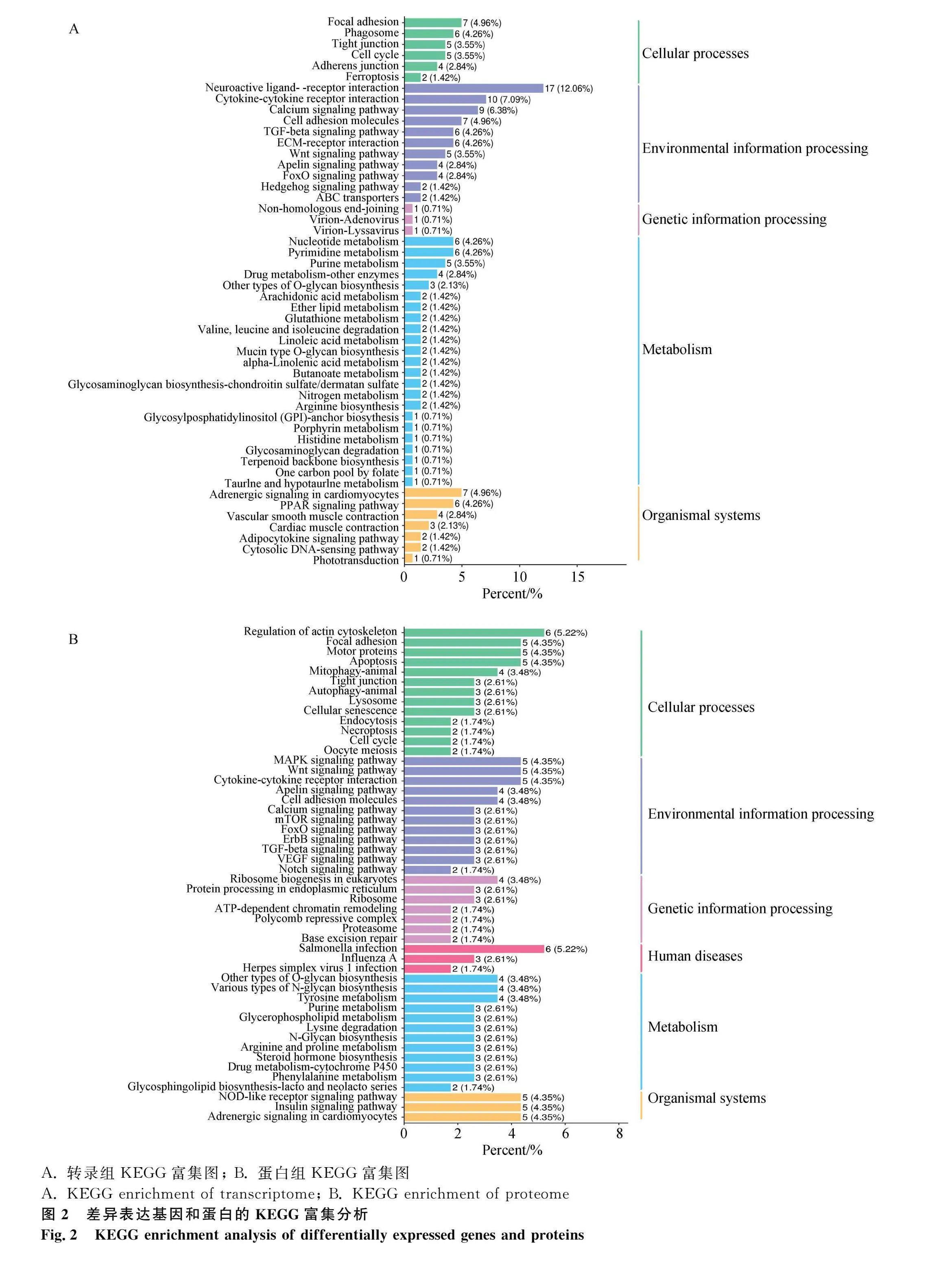
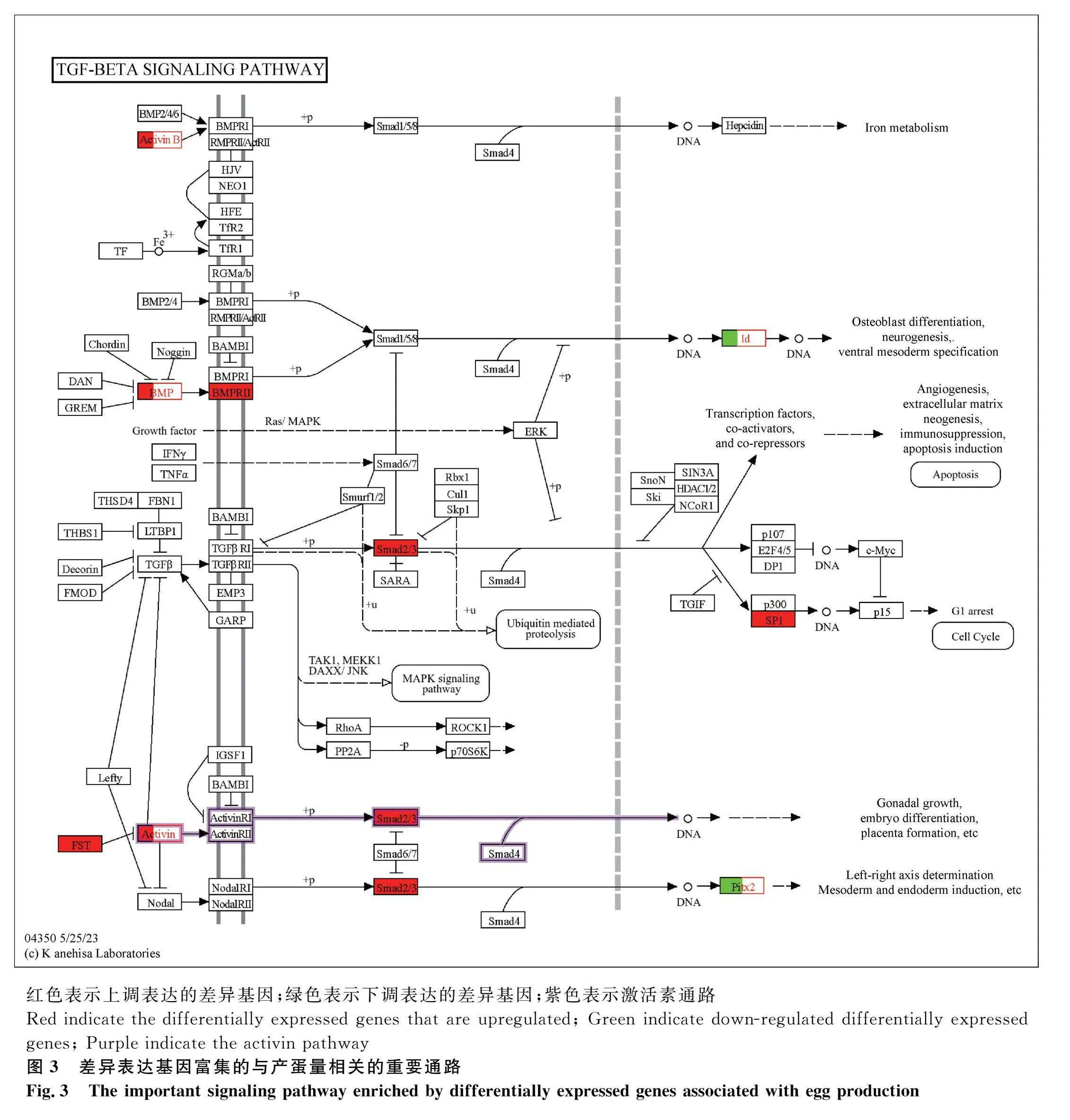
對DEGs進行KEGG富集分析,共富集到108條通路,發現DEGs主要富集于代謝途徑(metabolic pathways)、神經活性配體-受體相互作用(neuroactive ligand-receptor interaction)、細胞因子-細胞因子受體相互作用(cytokine-cytokine receptor interaction)、TGF-β信號通路(TGF-β signaling pathway)等通路,其中與卵泡發育相關的通路有TGF-β信號通路(TGF-β signaling pathway)、卵母細胞減數分裂(oocyte meiosis)、孕激素介導的卵母細胞成熟(progesterone-mediated oocyte maturation)、GnRH信號通路(GnRH signaling pathway)等。根據顯著性通路進一步篩選到INHBB、BMP5、PTTG1等與調控產蛋性狀相關的DEGs(圖2A)。其中,TGF-β信號通路的分支,激活素通路可能是調控產蛋的關鍵通路。在激活素信號通路中,FST通過作用于抑制素(INHA、INHBB)阻礙激活素與受體相結合,使SMAD2/3磷酸化增強,從而促進性腺生長以及胚胎分化(圖3)。對DEPs進行KEGG富集分析,共富集到101個相關的信號轉導過程,排名前50的信號通路(圖2B)顯示,DEPs主要富集于肌動蛋白細胞骨架(regulation of actin cytoskeleton)、MAPK信號通路(MAPK signaling pathway)、胰島素信號通路(insulin signaling pathway)、TGF-β信號等通路(TGF-β signaling pathway)等。
將DEGs和DEPs進行共同富集,分析結果顯示,共有364個顯著的GO條目(209個生物學過程,61個細胞組分,94個分子功能)和75個顯著的KEGG通路(圖4)。其中,GO條目主要涉及生殖結構發育(gland development)、性別分化(sex differentiation)、性激素活性(hormone activity)等,KEGG通路主要富集到TGF-β信號通路(TGF-β signaling pathway)、卵母細胞減數分裂(oocyte meiosis)、GnRH信號通路(GnRH signaling pathway)、孕激素介導的卵母細胞成熟(progesterone-mediated oocyte maturation)等,它們與產蛋性能高度相關,這些相關條目和通路主要涉及INHBB、FST、BMP5、PITX2、PTTG1、PRKCA、INHA、INSR、PPP3CB、PPP2R5D、EGR1、AKT3和MSC等基因。
2.4 蛋白互作網絡分析
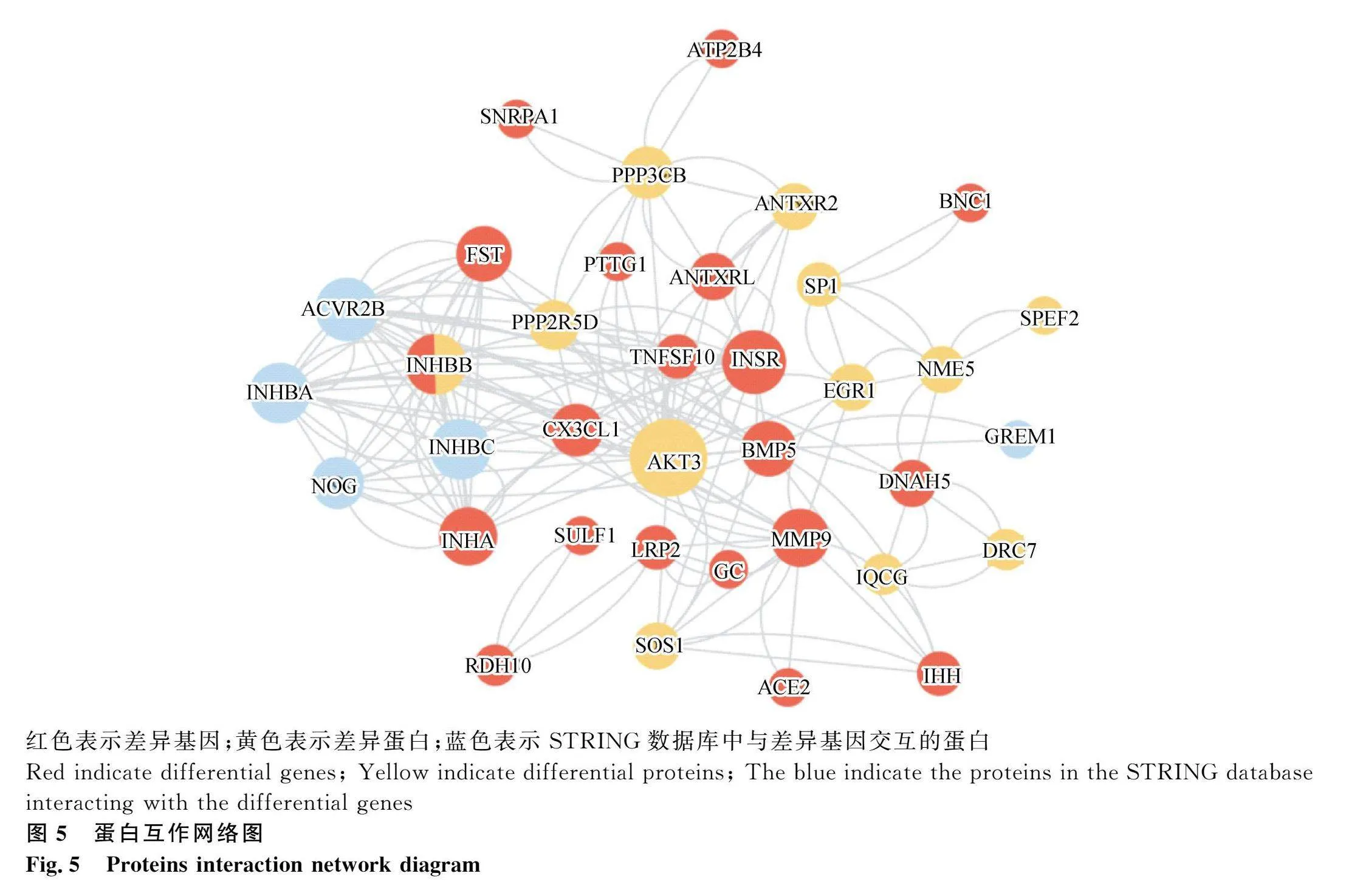
基于GO和KEGG的分析結果,將篩出的產蛋性能相關的候選基因和交集基因,利用在線數據庫STRING構建蛋白互作網絡(圖5),結果共匹配到38個編碼蛋白,富集到36個節點。經過對網絡中節點數和節點中心性的評估與分析,發現AKT3位于網絡節點中心,周圍與INSR、INHBB、INHA等多個蛋白節點連接,從而揭示其在產蛋過程的核心樞紐地位。綜合以上分析,最終確定了AKT3、INSR、INHBB、FST、BMP5、INHA、PPP2R5D、PTTG1、LRP2、TNFSF10、EGR1這11個基因為產蛋性能相關的核心基因,極有可能與產蛋量之間存在顯著且重要的關聯。
2.5 熒光定量PCR驗證
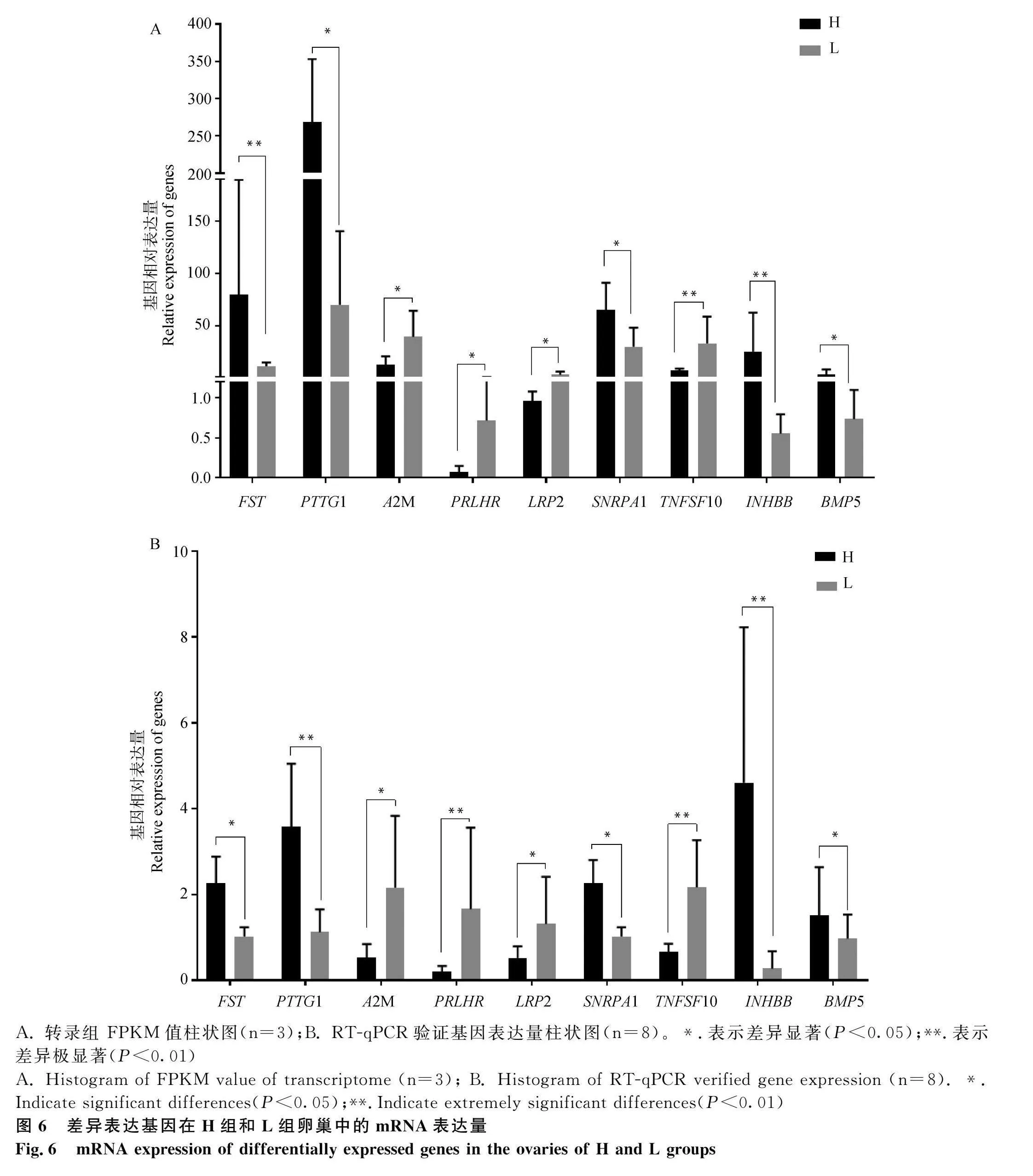
在產蛋相關的潛在候選基因中,由于轉錄組測序中,AFAP1、INHA、FNDC1基因的FPKM值和原始reads數值較低,可能對驗證結果的精確性構成不利影響,因此,在GO注釋和KEGG分析結果等的綜合考慮下,決定選擇9個DEGs對高產組和低產組的卵巢組織樣本(每組8個,且包含轉錄組測序的樣品)進行RT-qPCR驗證,以L組為對照做均一化處理,結果表明,9個基因的表達量變化趨勢與轉錄組測序結果相同,并且表達水平在高低產五龍鵝卵巢組織中均呈現顯著差異,其中PTTG1、PRLHR、TNFSF10、INHBB基因表現出極顯著差異(P<0.01)(圖6)。
3 討 論
家禽產蛋量與卵泡發育密切相關,卵泡發育包括原始卵泡、生長卵泡、成熟卵泡三個階段的發育,其中涉及生長因子、激素、血管生成和肌肉組織等相關基因的交互作用。為解析影響產蛋量的分子機制,本研究利用高、低產蛋水平的五龍鵝進行轉錄組和蛋白組聯合分析,篩選與產蛋量相關的候選基因,為后續利用分子標記輔助育種培育鵝高繁配套系奠定基礎。
在本研究中,通過對五龍鵝高產組和低產組轉錄組和蛋白組差異比較,發現DEGs主要富集在細胞因子-細胞因子受體相互作用(cytokine-cytokine receptor interaction)、鈣離子信號通路(calcium signaling pathway)、MAPK信號通路(MAPK signaling pathway)、TGF-β信號(TGF-β signaling pathway)、黏著斑(focal adhesion)等通路。有研究發現,王瑩等[10-11]在雞卵巢顆粒細胞轉錄組調控研究中,也顯著富集到GnRH信號通路(GnRH signaling pathway)、鈣離子信號通路(calcium signaling pathway)、細胞因子-細胞因子受體相互作用(cytokine-cytokine receptor interaction)、TGF-β信號通路(TGF-β signaling pathway)、MAPK信號通路(MAPK signaling pathway)、VEGF信號通路(VEGF signaling pathway)等通路。兩者在通路富集結果種類中有相同之處,進一步強調了這些通路的在卵巢發育中的重要性。其中,細胞因子-細胞因子受體相互作用(cytokine-cytokine receptor interaction)通路可能在促進雞卵泡顆粒細胞增殖起關鍵作用,而且參與輸卵管的收縮和蛋殼的形成,影響蛋的排出和蛋殼的質量[12-13]。劉建高[14]通過蛋白組學技術在蛋鴨卵泡發育調控研究中發現,鈣離子信號通路(calcium signaling pathway)可能通過影響卵泡顆粒細胞的凋亡和能量代謝來調控卵泡的發育和排卵;并且通過影響輸卵管上皮細胞的鈣吸收和分泌功能來確保蛋殼的正常形成。MAPK信號通路(MAPK signaling pathway)的激活可以促進顆粒細胞分泌FSH(促卵泡激素)和LH(促黃體生成素)受體等多種生長因子和激素,促進卵泡的生長發育;并且通過調控卵泡壁細胞的收縮和卵泡液的分泌,確保卵泡正常破裂釋放卵母細胞,完成排卵[15]。綜上研究表明,這些通路在卵泡發育、蛋殼形成以及排卵等過程中具有重要作用。
卵泡發育是一個復雜的過程,在這個過程中細胞凋亡起著重要的調控作用,卵泡中的顆粒細胞和卵母細胞之間的相互作用以及卵泡微環境的變化均會影響卵泡的發育和成熟[16-17]。TNFSF10作為一種促凋亡因子,屬于腫瘤壞死因子(TNF)配體家族的成員,通過誘導細胞凋亡來清除異常或受損細胞,減少炎癥因子的來源,從而間接調控炎癥反應[18]。已有研究發現,炎癥因子如腫瘤壞死因子(tumor necrosis factor,TNF)[19]、白細胞介素(interleukin,IL)等通過影響HPO軸(下丘腦-垂體-卵巢軸)的神經內分泌活動來調控卵泡的發育和排卵。本研究發現,TNFSF10在五龍鵝卵巢的表達量高產組顯著低于低產組,提示該基因可能在五龍鵝卵泡顆粒細胞凋亡過程中發揮重要作用,進而對卵泡的發育產生影響。因此推測,低產組鵝體內可能存在炎癥反應,進而影響其卵泡發育質量,導致產蛋量降低。
卵母細胞減數分裂是卵母細胞發育成熟的最后一個階段,是受精、胚胎著床前不可或缺的先決條件,PTTG1、PPP2R5D、PPP3CB這3個基因均在此過程中發揮重要作用。PTTG1在豬胚胎發生過程中參與卵母細胞成熟和合子基因組激活的過程[20],并且在馬和驢等動物的GV期和MⅡ期的卵母細胞中也發現PTTG1都有高豐度表達[21]。與之相似,本研究也發現了PTTG1在高產組鵝卵巢中高豐度表達。由此推測,該基因可能通過促進卵母細胞的成熟發育,進而影響五龍鵝的產蛋水平。此外,PPP2R5D、PPP3CB這兩個基因是磷蛋白磷酸酶(PPP)家族重要成員。有研究證實,PPP3CB參與調控豬青春期卵母細胞減數分裂過程[22],而且,當PPP2R5D亞基中的絲氨酸-丙氨酸發生突變時,會抑制小鼠卵泡的正常去磷酸化[23],從而影響正常的卵母細胞減數分裂過程,提示該基因可能通過干預卵泡生長發育影響五龍鵝產蛋水平。
產蛋量與蛋重之間存在一定的關系,但這種關系受多種因素的影響。多項研究證實,蛋重與產蛋量之間存在較強的負相關[24]。Yi等[25]也在蛋清形成的膨大部發現,LRP2是影響蛋重的重要基因。LRP2作為一種多功能內吞受體[26],通過內吞作用有效攝取與其載體蛋白相結合的重要維生素和激素,包括維生素D結合蛋白[27]、視黃醇結合蛋白[28]和性激素結合球蛋白[29-30]等,這些物質的攝取對卵巢的發育具有重要作用。當維生素D攝取不足時,會影響抗繆勒管激素的分泌,這是一種由卵巢顆粒細胞分泌的激素,對卵巢的儲備功能和排卵過程有重要影響[31-33]。此外,維生素D的缺乏還可能導致生殖系統微血管功能異常,影響子宮內膜的營養供應和修復過程,從而影響卵巢的正常功能[34]。因此,LRP2作為母胎脂質轉運途徑的胚胎脂蛋白受體,介導脂蛋白攝取到卵黃囊中,對卵泡的發育具有重要意義。在本研究中,低產組LRP2表達量顯著高于高產組,這種高表達促進了營養物質向卵泡的有效輸送,確保了卵泡發育過程中獲得充足的營養支持。因此,這種營養分配的差異可能是導致低產鵝的種蛋較重、產蛋量較低的重要因素之一。
通過蛋白互作網絡分析得知,INHBB、INHA、FST(卵泡抑素)這3個基因相互之間有著密切的關聯,有研究證實其在影響產蛋性狀中起重要作用。在雌性動物卵巢發育過程中,抑制素、激活素和卵泡抑素都是重要的調節因子,這三者組成的抑制素-激活素-卵泡抑素系統在動物的各個生殖階段都發揮著重要的調控作用[35]。有研究證實,在激活素信號通路中,FST作用于抑制素,從而阻止激活素與其受體的結合[36],導致SMAD2/3磷酸化增強,這進一步促進了性腺的生長和胚胎的分化(圖3)[37-38]。本研究中,這3個基因在高產組的mRNA表達水平高于低產組,當敲除INHBB時,會影響顆粒細胞的生長和凋亡[39-40],而過表達時,其會和FSH相互作用影響類固醇激素下調,導致卵泡發育受阻[41]。當然,在正常情況下,一定水平的INHBB會適當的抑制FSH分泌。由此推斷,在產蛋過程中,這3個基因通過促進性腺生長發育和調控生殖激素平衡,從而促進產蛋量增加。
4 結 論
本研究通過轉錄組和蛋白組進行聯合分析,最終篩選到6個與產蛋性能相關的候選基因(PTTG1、LRP2、TNFSF10、INHA、INHBB、FST),為五龍鵝產蛋性能的分子育種和產蛋性能調控機制的闡明提供參考。
參考文獻(References):
[1] 侯水生,劉靈芝.2022年水禽產業現狀、未來發展趨勢與建議[J].中國畜牧雜志,2023,59(3):274-280.
HOU S S,LIU L Z.Current status,future development trends,and suggestions for the waterbird industry in 2022[J].Chinese Journal of Animal Science,2023,59(3):274-280.(in Chinese)
[2] 楊景晁,李顯耀,王寶維,等.山東省水禽產業的發展基礎及“十四五”高質量發展策略[J].家畜生態學報,2023,44(6):92-96.
YANG J C,LI X Y,WANG B W,et al.Development foundation of Shandong waterfowl industry and its 14th five-year high quality development strategy[J].Acta Ecologae Animalis Domastici,2023,44(6):92-96.(in Chinese)
[3] 胡彥競科.四川白鵝GnRH、GnIH基因克隆、多態性及其與產蛋量的關聯性研究[D].重慶:西南大學,2017.
HU Y J K.Cloning,polymorphism of GnRH and GnIH genes and their association with egg production in Sichuan white goose[D].Chongqing:Southwest University,2017.(in Chinese)
[4] CHANG Y G,GUO R B,ZENG T,et al.Analysis of transcriptomic differences in the ovaries of high- and low-laying ducks[J].Genes (Basel),2024,15(2):181.
[5] WANG Y,WANG S,ZANG Z,et al.Molecular and transcriptomic analysis of the ovary during laying and brooding stages in Zhedong white geese (Anser cygnoides domesticus)[J].Br Poult Sci,2024,65(5):631-644.
[6] 陳 靜,吳薛蓓,苗冬枝,等.產蛋間隔前期鴿卵泡轉錄組比較分析揭示卵泡發育相關基因[J].畜牧獸醫學報,2024,55(8):3503-3515.
CHEN J,WU X B,MIAO D Z,et al.Comparative analysis of transcriptome of pigeon follicles at early stage of laying interval reveals genes related to follicular development[J].Acta Veterinaria et Zootechnica Sinica,2024,55(8):3503-3515.(in Chinese)
[7] 袁素珍.EGR1調控的顆粒細胞凋亡在卵巢衰老中的作用及其機制研究[D].武漢:華中科技大學,2017.
YUAN S Z.The role of EGR1-regulated granulosa cell apoptosis in ovarian aging and its mechanism[D].Wuhan:Huazhong University of Science and Technology,2017.(in Chinese)
[8] 黃 宣,尹兆正,徐春暉,等.基于TMT技術的不同產蛋性能母雞卵巢蛋白組學研究[J].中國畜牧雜志,2024,60(5):139-146.
HUANG X,YIN Z Z,XU C H,et al.Ovarian proteomics of hens with different laying performance based on TMT technology[J].Chinese Journal of Animal Science,2024,60(5):139-146.(in Chinese)
[9] 孫金艷,彭福剛,趙秀華,等.種鵝繁殖性能的影響因素[J].黑龍江動物繁殖,2020,28(5):41-43.
SUN J Y,PENG F G,ZHAO X H,et al.Factors affecting reproductive performance of breeding geese[J].Heilongjiang Journal of Animal Reproduction,2020,28(5):41-43.(in Chinese)
[10] 王 瑩,李婉晴,賀文慶,等.紅光對京海黃雞卵巢轉錄組模式調控分析[J].中國畜牧雜志,2023,59(12):205-211.
WANG Y,LI W Q,HE W Q,et al.Analysis of red light on the regulation of transcriptome pattern in the ovary of Jinghai yellow chicken[J].Chinese Journal of Animal Science,2023,59(12):205-211.(in Chinese)
[11] 冉明霞.miR-202-5p調控鵝顆粒細胞增殖凋亡、脂質沉積和類固醇激素合成的機制研究[D].成都:四川農業大學,2023.
RAN M X.Research on the mechanism of miR-202-5p regulating the proliferation,apoptosis,lipid deposition and steroid hormone synthesis of goose follicular granulosa cells[D].Chengdu:Sichuan Agricultural University,2023.(in Chinese)
[12] 韓昆鵬.高、低產蛋量京海黃雞卵巢組織轉錄組學分析[D].揚州:揚州大學,2016.
HAN K P.Transcriptomics analysis of ovaries of high and low egg production Jinghai yellow chicken[D].Yangzhou:Yangzhou University,2016.(in Chinese)
[13] LIU Z P,CHAO J R,XU P T,et al.Lonicera flos and Cnicus japonicus extracts improved egg quality partly by modulating antioxidant status,inflammatory-related cytokines and shell matrix protein expression of oviduct in laying hens[J].Poult Sci,2023,102(4):102561.
[14] 劉建高.鈣相關信號參與蛋鴨卵泡發育調控的作用及機制研究[D].長沙:湖南農業大學,2021.
LIU J G.Functions of calcium signalling in mediating follicular development in laying ducks and its mechanism[D].Changsha:Hunan Agricultural University,2021.(in Chinese)
[15] MATHIEN S,TESNI RE C,MELOCHE S.Regulation of mitogen-activated protein kinase signaling pathways by the ubiquitin-proteasome system and its pharmacological potential[J].Pharmacol Rev,202 73(4):263-296.
[16] LIN F,FU Y H,HAN J,et al.Changes in the expression of FoxO1 and death ligand genes during follicular atresia in porcine ovary[J].Genet Mol Res,2014,13(3):6638-6645.
[17] KULUS M,KRANC W,SUJKA-KORDOWSKA P,et al.The processes of cellular growth,aging,and programmed cell death are involved in lifespan of ovarian granulosa cells during short-term IVC-Study based on animal model[J].Theriogenology,2020,148:76-88.
[18] 侯增鑫.尼羅羅非魚TNFSF10(TRAIL)免疫功能初步研究[D].廣州:中山大學,2017.
HOU Z X.Primary study on immune function of TNFSF10(TRAIL)in Tilapia,Oreochromis niloticus[D].Guangzhou:Sun Yat-Sen University,2017.(in Chinese)
[19] 鐘 佩.生殖道微生物及血清代謝物對胚胎移植后妊娠結局的影響及機制探究[D].合肥:安徽醫科大學,2023.
ZHONG P.The impact and mechanism of reproductive tract microbiota and serum metabolites on pregnancy outcomes after embryo transfer[D].Hefei:Anhui Medical University,2023.(in Chinese)
[20] XIE B K,QIN Z X,LIU S,et al.Cloning of porcine pituitary tumor transforming gene 1 and its expression in porcine oocytes and embryos[J].PLoS One,2016,11(4):e0153189.
[21] 嚴嘉耕.比較轉錄組揭示影響馬和驢卵母細胞體外成熟基因的初步研究[D].南寧:廣西大學,2023.
YAN J G.Comparative transcriptome reveals a preliminary study of genes influencing invitro maturation of horse and donkey oocytes[D].Nanning:Guangxi University,2023.(in Chinese)
[22] LI Q N,PAN X C,LI N,et al.Identification of circular RNAs in hypothalamus of gilts during the onset of puberty[J].Genes (Basel),202 12(1):84.
[23] EGBERT J R,SILBERN I,ULIASZ T F,et al.Phosphatases modified by LH signaling in ovarian follicles:testing their role in regulating the NPR2 guanylyl cyclase[J].Biol Reprod,2024,110(1):102-115.
[24] 嚴西萍.鴨產蛋量和蛋重關鍵基因的篩選及其調控機制研究[D].成都:四川農業大學,2023.
YAN X P.Screening the candidate genes and regulatorypathways associated with the trait of egg-production and egg weight in ducks[D].Chengdu:Sichuan Agricultural University,2023.(in Chinese)
[25] YI G Q,SHEN M M,YUAN J W,et al.Genome-wide association study dissects genetic architecture underlying longitudinal egg weights in chickens[J].BMC Genomics,2015,16(1):746.
[26] FARIDI R,YOUSAF R,GU S J,et al.Variants of LRP2,encoding a multifunctional cell-surface endocytic receptor,associated with hearing loss and retinal dystrophy[J].Clin Genet,2023,103(6):699-703.
[27] TOI N,INABA M,ISHIMURA E,et al.Significance of urinary C-megalin excretion in vitamin D metabolism in pre-dialysis CKD patients[J].Sci Rep,2019,9(1):2207.
[28] CHARLTON J R,HARER M W,SWAN C,et al.Immature megalin expression in the preterm neonatal kidney is associated with urinary loss of vitamin carrier proteins[J].Pediatr Res,2019,85(3):405-411.
[29] MARKO H L,HORNIG N C,BETZ R C,et al.Genomic variants reducing expression of two endocytic receptors in 46,XY differences of sex development[J].Hum Mutat,2022,43(3):420-433.
[30] GARCIA J,KRIEGER K D,LOITZ C,et al.Regulation of prostate androgens by Megalin and 25-hydroxyvitamin D status:Mechanism for high prostate androgens in African American Men[J].Cancer Res Commun,2023,3(3):371-382.
[31] SIMPSON S,PAL L.Vitamin D and infertility[J].Curr Opin Obstet Gynecol,2023,35(4):300-305.
[32] NELSON S M,DAVIS S R,KALANTARIDOU S,et al.Anti-Müllerian hormone for the diagnosis and prediction of menopause:a systematic review[J].Hum Reprod Update,2023,29(3):327-346.
[33] MORIDI I,CHEN A,TAL O,et al.The association between vitamin D and Anti-Müllerian Hormone:A systematic review and meta-analysis[J].Nutrients,2020,12(6):1567.
[34] V RB R "S,TAK CS I,TU′′U′′ L,et al.Effects of vitamin D on fertility,pregnancy and polycystic ovary syndrome-a review[J].Nutrients,2022,14(8):1649.
[35] SEACHRIST D D,KERI R A.The activin social network:Activin,inhibin,and follistatin in breast development and cancer[J].Endocrinology,2019,160(5):1097-1110.
[36] THOMPSON T B,LERCH T F,COOK R W,et al.The structure of the follistatin:Activin complex reveals antagonism of both type I and type II receptor binding[J].Dev Cell,2005,9(4):535-543.
[37] 付獻歐.鵝胚性腺形態結構發育特征及MAPK、TGF-β/Smad信號通路的表達調控研究[D].長春:吉林農業大學,2023.
FU X O.Morphological structural and developmental characteristics of goose embryonic gonads and the MAPK and TGF-β/Smad expression and regulation of the signaling pathway[D].Changchun:Jilin Agricultural University,2023.(in Chinese)
[38] 郭澤媛,杜張勝,張雅琦,等.Smad7介導TGF-β信號通路對綿羊卵泡顆粒細胞增殖的影響[J].中國農業科學,2023,56(13):2597-2608.
GUO Z Y,DU Z S,ZHANG Y Q,et al.Effects of Smad7-mediated TGF-β signaling pathway on proliferation of sheep granulosa cells[J].Scientia Agricultura Sinica,2023,56(13):2597-2608.(in Chinese)
[39] 李芝豐,孫 偉,儲明星.TGF-β通路相關基因在綿羊性腺軸組織的表達分析[J].東北農業大學學報,202 52(2):43-49.
LI Z F,SUN W,CHU M X.Expression analysis of TGF-β pathway related genes in gonadal axis-associated tissues[J].Journal of Northeast Agricultural University,202 52(2):43-49.(in Chinese)
[40] M’BAYE M,HUA G H,KHAN H A,et al.RNAi-mediated knockdown of INHBB increases apoptosis and inhibits steroidogenesis in mouse granulosa cells[J].J Reprod Dev,2015,61(5):391-397.
[41] 張智慧,高鴻霞,王國慶,等.VEGF對小鼠卵巢類固醇合成相關基因表達的影響及其機制[J].解放軍醫學雜志,2024,49(6):679-685.
ZHANG Z H,GAO H X,WANG G Q,et al.Effect of VEGF on the expression of genes related to ovarian steroid synthesis in mice and its mechanism[J].Medical Journal of Chinese People’s Liberation Army,2024,49(6):679-685.(in Chinese)
(編輯 郭云雁)
