根損傷誘導煙草根系分泌物變化及對疫霉生長的影響
2024-10-31 00:00:00李振張本強楊帆楊俊杰張鑫陳淑童姚慧解程浩白瑞李義強尤祥偉時軍
江蘇農業科學 2024年17期

摘要:為探究地下害蟲咬食影響煙草黑脛病的發病機制,選擇機械損傷為陽性對照,研究無損傷(CK)、機械損傷(T1)和害蟲咬食(T2)對煙草不同部位酶活性和次級代謝產物含量的影響,從根損傷影響根系分泌物的代謝組學和疫霉菌的化感作用角度來解析蟲害加重病害發生的作用機制。結果表明:(1)與CK相比,T1組地上部和地下部的過氧化氫酶(CAT)、過氧化物酶(POD)、超氧化物歧化酶(SOD)活性顯著升高,T2組地上部酶活性提高,而地下部SOD活性顯著下降;(2)T1、T2組誘導煙草地下部次級代謝產物發生顯著變化,降低有機酸、可溶性糖含量,提高丙二醛、多酚含量;(3)代謝組學分析共發現293種差異代謝物,T1、T2與CK組比較后差異代謝物總數、上調數和下調數排序為T2(271、235、36)>T1(30、24、6)。(4)T2組根系分泌物濃度為50、150 μg/mL對疫霉菌趨化移動指數(CMI)的影響呈顯著正趨化作用,且各處理對煙草疫霉菌游動孢子萌發率的相對促進率分別為8.13%、44.13%、45.20%。綜上所述,蟲害可通過影響煙草防御性酶活性、改變次級代謝產物含量、誘導根系分泌物對疫霉的化感作用等方式加重病害發生。
關鍵詞:黑脛病;小地老虎;根系分泌物;化感作用
中圖分類號:S435.72 文獻標志碼:A
文章編號:1002-1302(2024)17-0108-07
收稿日期:2023-10-23
基金項目:廣西中煙有限公司橫向科技項目(編號:2023370200240167)。
作者簡介:李 振(1999—),男,山東濟南人,碩士研究生,主要從事植物病理學研究。E-mail:lzz15953171758@163.com。
通信作者:時 軍,碩士,高級政工師,主要從事煙葉生產方面的研究。E-mail:619045869@qq.com。
蟲害與病害的發生存在一定關聯[1-2]。煙草生長過程中,害蟲的咬食是導致煙草病害發生的主要原因之一。盡管煙草會通過改變某些途徑來抵御這些損害帶來的負面影響,但害蟲咬食導致細胞壁受損,內容物外流,增大了病原菌侵染的機會。同時,內容物的外流還為病原菌提供營養物質,有利于微生物的生長繁殖[3-5]。此外,植物會誘導化感物質來直接抵御或增強病原菌的侵染,已知的一些氣味物質如β-丁烷、1-辛醇等,可以激活植物本身的免疫反應,有助于植物抵御病原菌的入侵[6-7]。相反,煙草根系分泌物中對土傳類病原菌具有正趨化作用的肉桂酸、苯甲酸等有機酸類自毒物質,使植物根系活性降低、養分吸收減少,增加了植物受病原菌侵染的易感性[5,8]。
蟲害加重病害發生不是單一因素決定,而是涉及多種因素的復雜過程[9-11]。目前關于蟲害加重煙草根莖病害發生的研究仍局限于傷口損傷產生的影響上,本研究為了進一步探討蟲害對煙草根莖病害加重的相關機制,以地下害蟲小地老虎咬食后的煙草根系分泌物作為研究對象,解析煙草-地下害蟲-病原菌三者生物相互作用關系,對病蟲害的防治具有理論指導意義。
本研究通過將機械損傷作為害蟲咬食的傷口對照,通過試驗驗證以下假設:相較于機械損傷,地下害蟲的咬食會對煙草的酶活性產生改變;同時,它也會誘導根系分泌物的改變,形成適宜疫霉菌生長繁殖的微環境,從而大大增加疫霉菌侵染煙草的可能性。
1 材料與方法
1.1 試驗材料
供試病原菌菌株為煙草疫霉(Phytophthora nicotianae)JM01,供試昆蟲為小地老虎,供試煙草品種為煙草黑脛病易感品種小黃金1025,均由中國農業科學院煙草研究所提供。燕麥培養基(OA)用于煙草疫霉培養。小地老虎飼料的制作與存儲:用玉米麩、大豆粉、酵母粉、瓊脂及復合維生素微波加熱后冷卻凝固4 ℃保存。
1.2 煙草樣品處理及根系分泌物收集
在中國農業科學研究院煙草研究所開展的盆栽試驗中,參照煙草水培法育苗[12-13]。試驗在3月份開始育苗,經過7 d后進行假植,15 d后將煙苗移栽至水培裝置中進一步培養,15 d左右進行后續處理。試驗設置了對照組(CK)、機械損傷處理組(T1,根部損傷一半)以及小地老虎咬食處理組(T2,持續咬食48 h),并在處理前用滅菌水清洗根部。處理期間使用滅菌水代替霍格蘭營養液,并每天補充至50 mL。在機械損傷后48 h取樣,將小地老虎提前饑餓處理12 h后放入煙草裸露的根部旁,待 48 h 后取樣。根系分泌物的收集參照張成省的方法[14]略作修改,待煙草處理結束后收集液體,將收集到的浸提液過0.22 μm微孔濾膜,-80 ℃冷凍再抽真空。
1.3 酶活性及代謝物測定
采用鄰苯三酚自氧化法進行超氧化物歧化酶(SOD)活性測定,以 25 ℃ 時抑制鄰苯三酚自氧化速率50%時消耗酶量定義為1個酶活性單位;采用紫外吸收法測定過氧化氫酶(CAT)活性,以1 min D240 nm值減少0.01為1個酶活性單位[U/(g·min)];采用愈創木酚法測定過氧化物酶(POD)活性,以1 min D470 nm的變化值表示酶活性大小[U/(g·min)];苯丙氨酸解氨酶活性測定參照植物L-苯丙氨酸解氨酶(PAL)ELISA檢測試劑盒使用說明書[15-19]。蛋白質、多糖等代謝物含量的測定參照《植物生理學實驗指導》[20]。
1.4 根系分泌物代謝組學
將收集到的不同處理下的根系分泌物進行冷凍干燥后,基于液質聯用(LC-MS)技術[21-22]進行非靶向代謝組學研究,試驗流程主要包括:樣本的代謝物提取、LC-MS檢測以及數據分析等。由北京諾禾致源科技股份有限公司采用LC-MS技術,完成非靶標代謝組學分析。
1.5 化感作用
參照徐周洋的化學趨向性響應方法[23],通過使用0.2%二甲基亞砜(DMSO)溶液溶解冷凍干燥后的根系分泌液,制備了50、100、500 μg/mL的分泌液濃度梯度。在燕麥培養基上等距離打孔(直徑為5 mm),按照添加物依次為對照品A(30 μL 2% DMSO)、疫霉菌B、趨化液C(溶于2% DMSO的根系分泌物30 μL)。每2孔之間用無菌濾紙條(長25 mm、寬2.5 mm)連接。在黑暗條件下,保持在28 ℃下培養3 d,每個處理重復3次。菌絲邊緣從B處移動到A的距離記作a,將菌絲邊緣從B處移動到C孔的距離記作b。利用趨化性指數(CMI)=b/a來衡量化學趨向性,當CMI-1>0時,表示測得的趨化液對疫霉菌具有正的化學趨向性。
1.6 孢子萌發試驗
參照張成省的孢子萌發試驗方法[14],取 0.5 mL 濃度分別為5、50、150 μg/mL的根系分泌物溶液與等體積疫霉菌游動孢子懸浮液(106 CFU/mL)。充分混勻后,用移液槍滴加到血細胞計數板中,28 ℃保濕培養10 h。用超景深顯微鏡觀察載玻片下孢子的萌發情況,并計算孢子萌發率。
1.7 數據處理與統計分析
將質譜檢測得到的根系分泌物通過Compound Discoverer 3.1,得到代謝物的定性定量結果。對代謝物進行多元統計分析,包括主成分分析(PCA)、偏最小二乘法判別分析(PLS-DA)和代謝富集通路。采用SPSS 26.0軟件進行差異顯著性檢驗(Duncan’s新復極差法,α=0.05),并采用Origin軟件繪圖。
2 結果與分析
2.1 不同根損傷對煙草不同部位防御性酶活性的影響
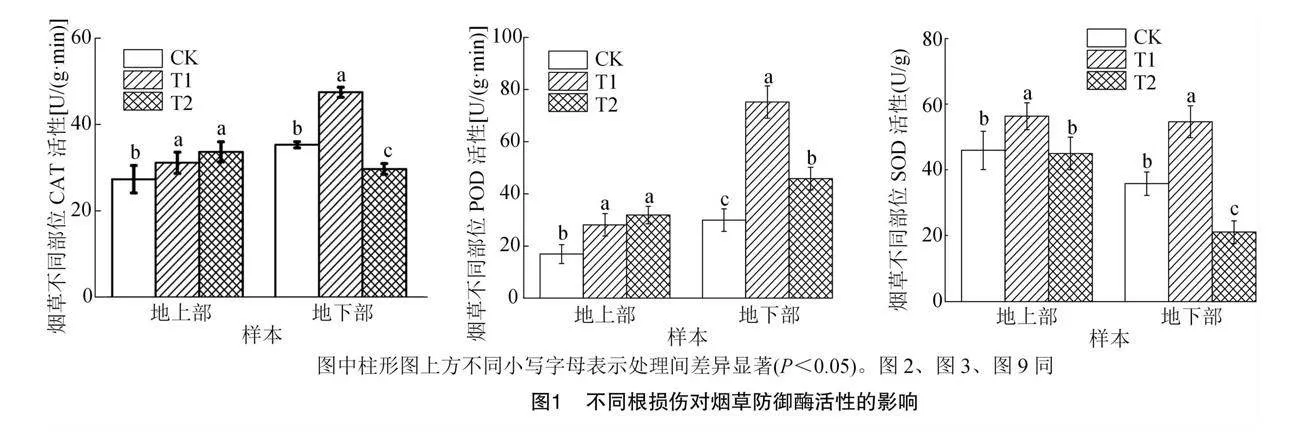
由圖1可知,T1和T2處理組中煙草不同部位酶活性響應不同,與對照相比,T1組地上部和地下部中與防御相關的酶活性均顯著升高,T2組地上部CAT、POD活性顯著升高,SOD活性無顯著變化,而地下部中SOD活性顯著降低,害蟲咬食降低了煙草地下部防御性酶活性,而誘導了地上部酶活性。
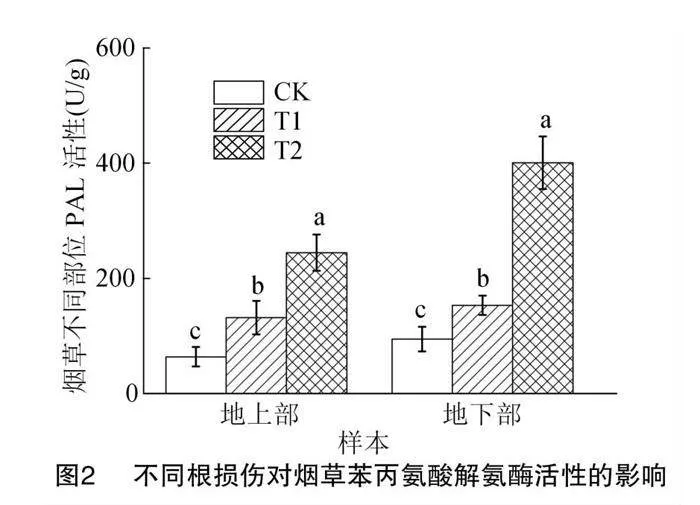
2.2 不同根損傷對煙草不同部位PAL酶活性及代謝產物的影響
苯丙氨酸解氨酶(PAL)是植物苯丙烷次生代謝中重要的限速酶類,對植物生長發育、次生代謝物合成等方面具有重大作用[24]。由圖2可知,T1和T2組都誘導了煙草中PAL活性的增加,且T2組酶活性增加程度高于T1。
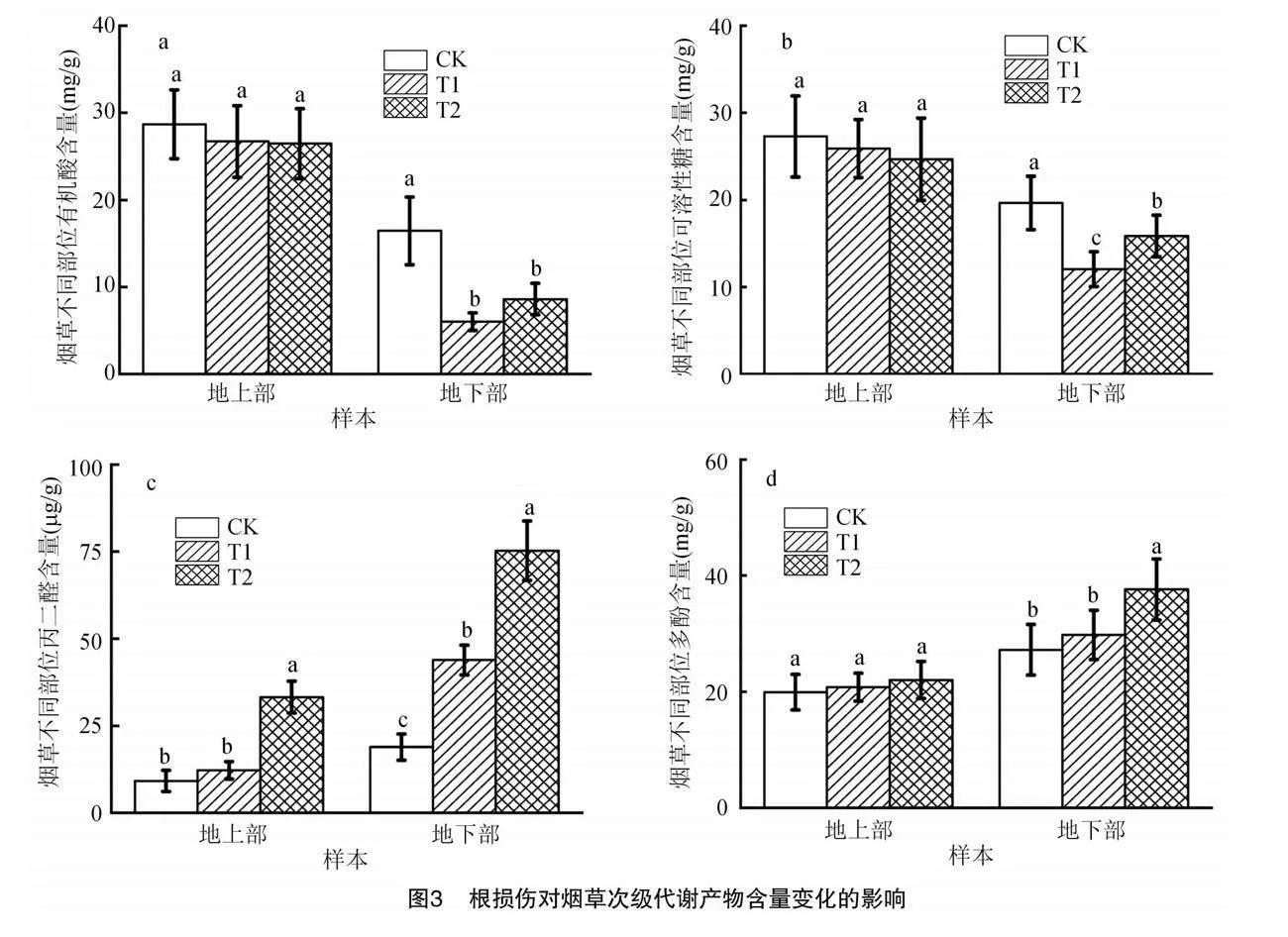
圖3為根損傷對煙草次級代謝產物的影響,其中地上部有機酸、可溶性糖、多酚含量在T1、T2和CK組之間均沒有顯著差異,而丙二醛含量在T2組中顯著增加。T1、T2組地下部有機酸、可溶性糖含量顯著低于CK組,T1組下降幅度更大(圖3-a、圖 3-b);T2組地下部多酚含量顯著高于CK和T1組(圖3-d);T1、T2組地下部丙二醛酚含量顯著高于CK組,T2組含量上升幅度更大(圖3-c)。
2.3 不同根損傷下煙草根系分泌物的代謝組學分析
2.3.1 PCA與OPLS-DA
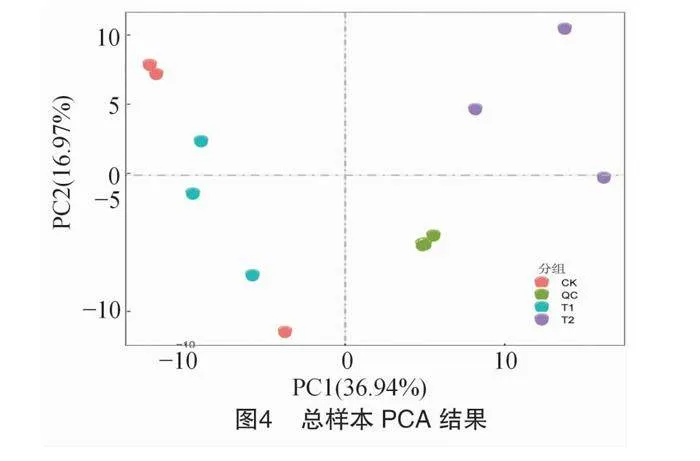
通過PCA分析不同處理方式下的代謝物樣品發現,PC1和PC2對方差的貢獻分別為36.94%和16.97%(圖4)。具體來說,PC1(36.94%)可以有效區分T2樣品與其他處理方式的差異,而CK和T1之間的差異并不顯著。此外,質控樣品(QC)呈現出緊密聚集的趨勢,這表明數據具有良好的穩定性和重復性。
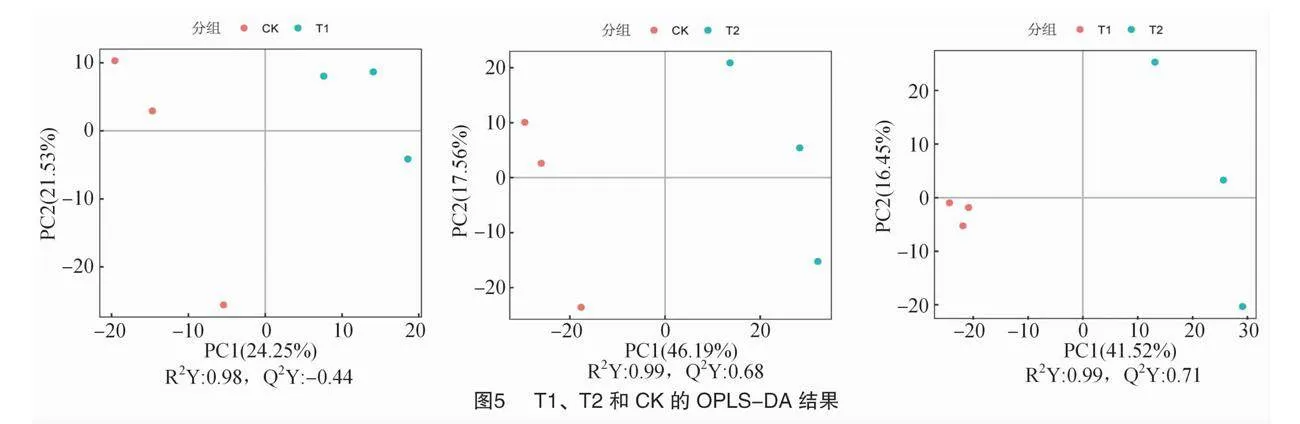
另外,通過OPLS-DA分析結果,將組間差異最大化反映在PC1上可以直接區分各處理之間的差異。如圖5所示,在T1、T2和CK之間分別進行OPLS-DA分析時,CK與T1比較中,可以看到它們沿著第1主成分進行分離,解釋了總變異的24.25%、21.53%;CK與T2比較中,也沿著第1主成分進行分離,解釋了總變異的46.19%、17.56%;T1與T2比較中,它們沿著第1主成分進行分離,解釋了總變異dT+QBCe79nXYhiBWu35QMg==的41.52%、16.45%;綜上可知,T1、T2和CK之間的代謝物組分存在差異,T2與對照(CK)之間組分差異顯著。
2.3.2 差異代謝物分析
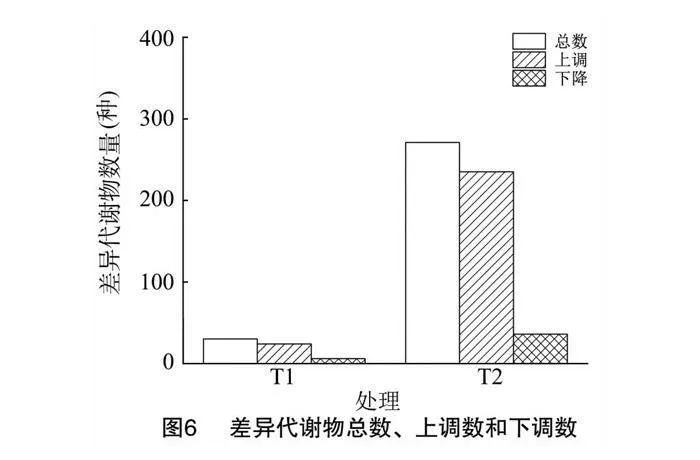
使用HMDB、Lipidmaps(v2.3)、METLIN數據庫等匹配已獲取的代謝物名稱,比較不同處理對煙草根系分泌物組分的影響,對T1、T2組中差異代謝物總數、上調數和下調數進行統計分析。圖6結果顯示,T1、T2組與對照組比較差異代謝物的總數、上調和下調數排序為T2(271、235、36)>T1(30、24、6)。
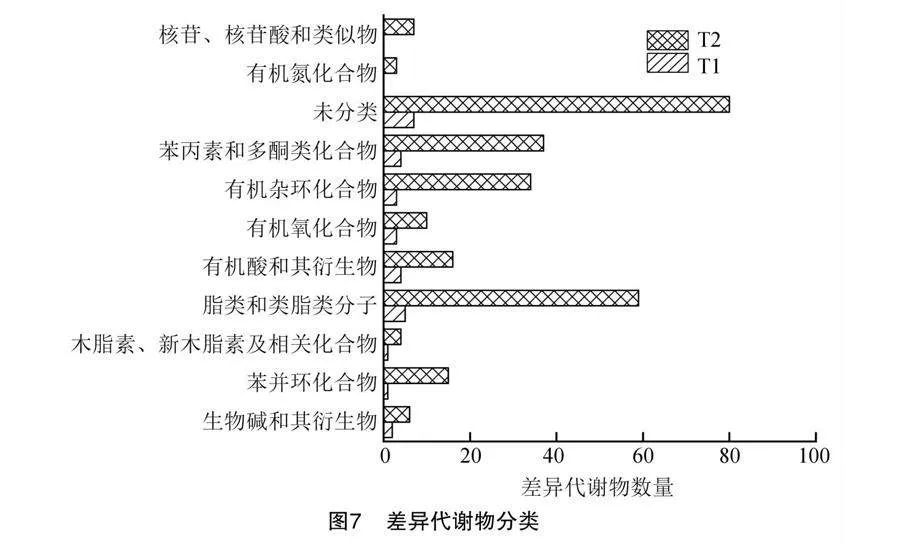
將T1、T2與對照比較后,對VIP>1的代謝物進行了統計分析,共發現293種差異代謝物。如圖7所示,主要包括生物堿和其衍生物(2.66%)、苯丙環化物(5.32%)、木質素、新木質素及相關化合物(1.66%)、脂類和類脂類分子(21.26%)、有機酸和其衍生物(6.64%)、有機氧化合物(4.32%)、有機雜環化合物(12.29%)、苯丙素和多酮類化合物(13.62%)、有機氮化合物(1.00%)、核苷、核苷酸和類似物(2.33%)及未分類的物質(28.90%)。其中有機氮化合物和核苷酸類化合物在T1組中未檢測到與該分類相關的代謝物,這些結果表明,T2組誘導產生的差異代謝物遠高于T1組,該影響主要集中在脂類分子,其次是有機酸、多酚類以及有機雜環化合物。
將VIP值排前28名的具體差異代謝物進行了統計分析。由表1可知,在與對照組相比后得到的不同處理組差異代謝物中,T1上調松柏苷、馬鞭草苷、5′-磷酸吡哆醛一水合物、N1-[3,5-二(三氟甲基)苯基]-2-環戊基-2-苯乙酰胺等10種物質,下調尼古丁、紫丁香苷、原百部堿等5種物質。與此類似,T2也展現出了更多不同的差異代謝物。T2上調了α-萘醌、穿心蓮內酯等11種物質,下調了丙酰肉堿、芽子堿甲酯。
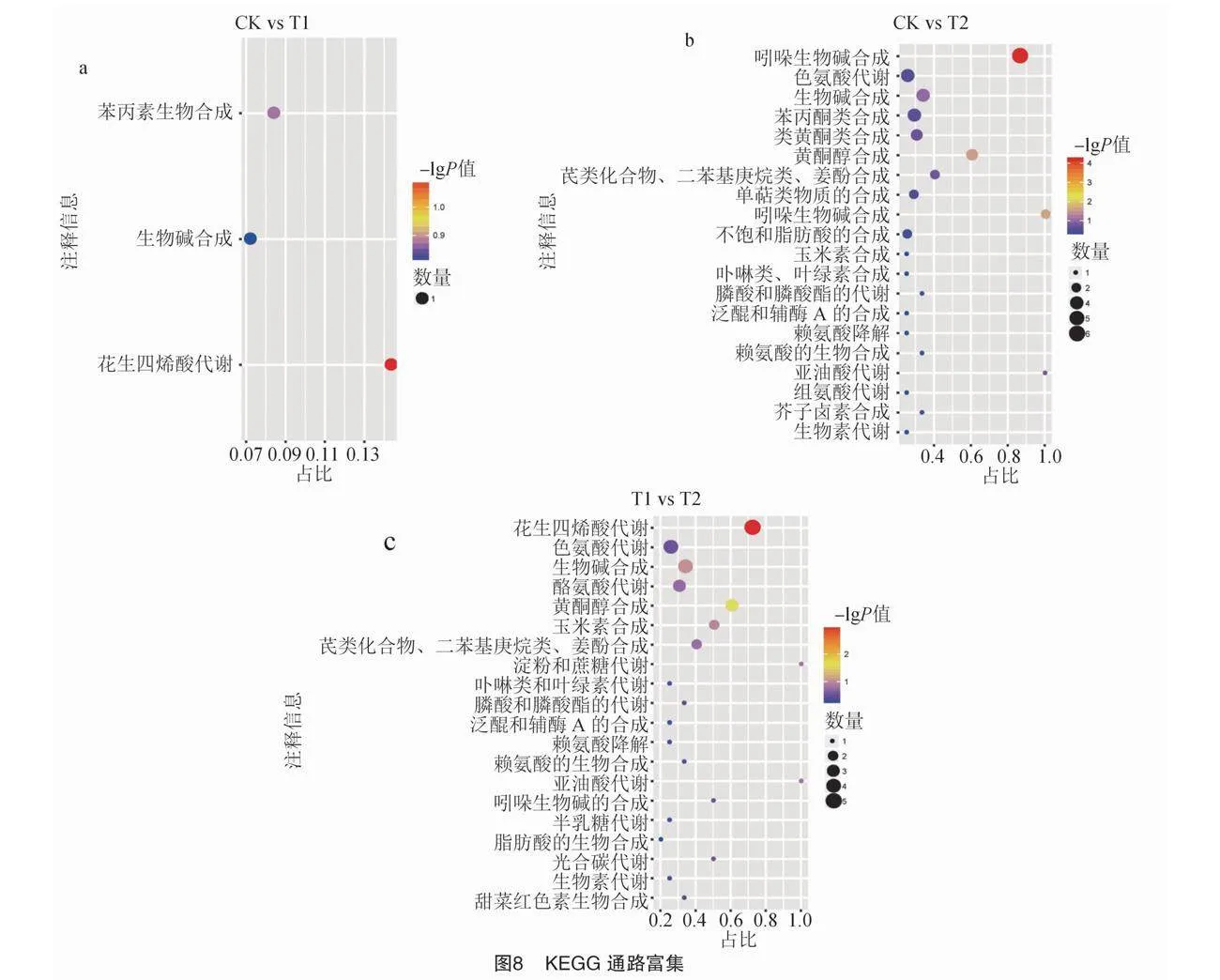
2.3.3 KEGG通路富集
根據KEGG通路富集分析(圖8),結果顯示T1、T2、CK組間比較,圖8共注釋到P<0.05的誘導煙草根系分泌物變化的代謝途徑。與CK相比,T1共注釋到3條代謝途徑,分別為花生四烯酸代謝、生物堿合成、苯丙素類生物合成途徑(圖8-a);與CK相比,T2注釋到的代謝通路共20條,涵蓋吲哚生物堿合成、色氨酸代謝、生物堿合成、苯丙酮類合成、類黃酮類合成等相關的代謝通路(圖8-b);與T1相比,T2注釋到的代謝通路最多,共22條,其中有20條代謝通路呈顯著富集,包括花生四烯酸代謝、色氨酸代謝、生物堿合成、酪氨酸代謝等相關的代謝通路(圖8-c)。
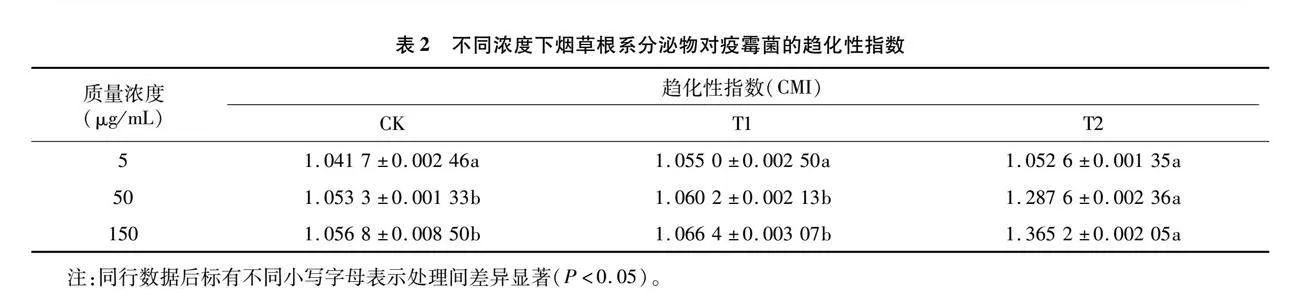
2.4 不同根損傷后的根系分泌物對疫霉菌趨化性的影響
趨化試驗結果(表2)表明,低、中和高濃度下CK和T1組無顯著差異,未表現出正趨化性,而T2組中、高濃度下的趨化指數均顯著高于CK、T1,表現出正趨化性,并且對疫霉菌的正趨化作用隨濃度升高而增大。
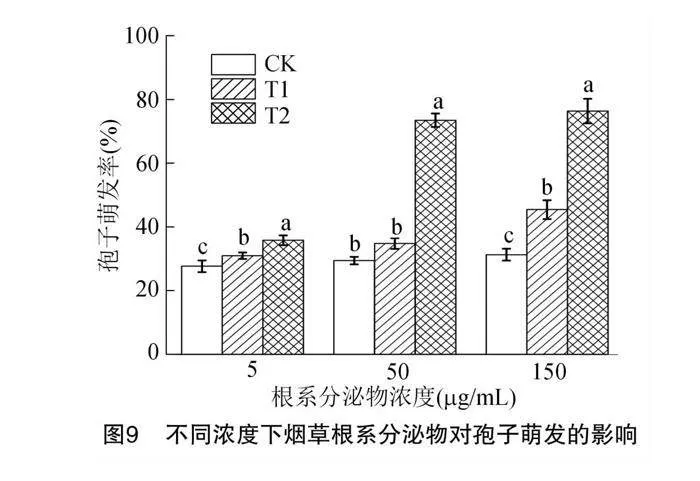
2.5 不同根損傷后的根系分泌物對孢子萌發影響
如圖9所示,試驗設CK、T1和T2各3個濃度(5、50、150 μg/mL),結果表明T2處理組在5、50、150 μg/mL 促進孢子萌發率顯著增加。以相應濃度CK組的孢子萌發率作為基線值,比較不同處理組的孢子萌發率與基線值的差異,發現T1組隨著濃度的升高孢子萌發率分別提高了3.30%、5.37%、14.17%;T2組隨著濃度的升高孢子萌發率分別上升了8.13%、44.13%、45.20%。與CK相比,T2組表現出了顯著的促進萌發作用。
3 討論
自然界中,植物、害蟲和病原菌之間構成了錯綜復雜的生態系統[25]。害蟲和病原菌會對煙草的生長和發育造成危害,害蟲咬食煙草組織,并從中吸取營養或分泌毒素,導致煙草受到更嚴重的損傷。Shi等研究發現,害蟲活動會促使植物產生一些次生代謝物質,而這些物質對微生物具有一定的化感作用,可進一步影響土壤微生物群落行為[26]。
本研究中,筆者所在課題組觀察到長期小地老虎取食會改變煙草根部的防御酶活性,誘導煙草代謝物含量變化,為疫霉菌侵害的發生創造更有利的環境。與高佳敏等研究結果[27]相似,發現取食處理對誘導植物生理代謝物質量分數的變化效果更加顯著。本研究發現小地老虎的取食會誘導煙草根部苯丙氨酸解氨酶的高表達,并調節煙草根部的次生代謝產物水平。重要的是,與機械損傷的影響相比,害蟲取食造成的影響要嚴重得多。然而苯丙氨酸途徑在產生各種中間產物方面起著重要作用,如反式肉桂酸、香豆酸、麥角酸和芥子酸[28]。這些酸可以進一步轉化為香豆素、綠原酸和輔酶A酯,最終導致木質素和其他植物生理代謝產物的合成,而這與構建病害發生的適宜環境密切相關。
根系分泌物在病原菌的生長和侵害中起著至關重要的作用[29-30]。通過代謝組學分析,發現機械損傷對煙草根系分泌物的影響較小,而昆蟲取食明顯改變了煙草根系分泌物,誘導了271種差異代謝產物,包括235種上調和36種下調的化合物。此外,還鑒定出了22個誘導途徑,包括氨基酸、生物堿、脂質、生物素、單萜、酚酸和黃酮類物質的生物合成途徑。蛋白質、可溶性糖等物質含量的測定證實了昆蟲取食直接導致煙草根系分泌物的變化,這會影響植物滲透壓、根際土壤微環境,改變植物抗逆能力,同時根系分泌物作為微生物的營養源之一,也會促進微生物的生長和移動[31]。黃玉茜等的研究表明,花生根系分泌物濃度通過影響土壤中的碳源,達到重塑土壤微生物群落結構的作用[32]。化感試驗表明,根系分泌物對疫霉菌具有正趨化效應,通過吸引疫霉菌生長的趨向性,增大疫霉菌接觸并侵染煙草的機會,從而提高煙草黑脛病發生的可能性。Li等研究發現,煙草根系分泌物中的化感自毒物質肉桂酸、肉豆蔻酸等加快了煙草枯萎病的致病能力[29]。
綜上所述,地下害蟲咬食煙草的根莖部會通過調節煙草的防御性酶活性,誘導或改變煙草根系的分泌物質,影響對疫霉菌的化感作用,導致黑脛病發生加重。本試驗僅初步探究了不同處理下根系分泌物的組件差異,關于具有化感作用的化合物分析及其對土壤微生物群落的影響還需要深入探討。
4 結論
害蟲咬食區別于單一的機械損傷,它誘導了地下部苯丙氨酸解氨酶的高表達,調節了煙草根部的CAT、POD、SOD活性和可溶性糖、有機酸次生代謝產物含量。害蟲咬食的根系分泌物對疫霉菌具有正趨化作用,這也會進一步影響病害的發生。本研究為蟲害加重病害發生途徑提供了新思路,為進一步指導如何通過調控根系分泌物的改變來更好地防治煙草土傳病蟲害的發生提供了理論指導。
參考文獻:
[1]高玉林,徐 進,劉 寧,等. 我國馬鈴薯病蟲害發生現狀與防控策略[J]. 植物保護,2019,45(5):106-111.
[2]謝銀燕,王 松,吳春銀,等. 木麻黃病蟲害及其防治的最新進展[J]. 江蘇農業科學,2019,47(20):36-41.
[3]董玉妹,張美倩,沈 慧,等. 植食性昆蟲唾液效應子和激發子的研究進展[J]. 昆蟲學報,2021,64(8):982-997.
[4]王 政,孟倩倩,鐘國華. 植食性昆蟲取食行為過程及機制研究[J]. 環境昆蟲學報,2014,36(4):612-619.
[5]李石力. 有機酸類根系分泌物影響煙草青枯病發生的機制研究[D]. 重慶:西南大學,2017:14.
[6]羅麗芬,江冰冰,鄧琳梅,等. 三七根系分泌物中幾種成分對根腐病原菌生長的影響[J]. 南方農業學報,2020,51(12):2952-2961.
[7]劉 暢. 甜瓜不同抗性品種根系分泌物對枯萎病菌的化感作用研究[D]. 沈陽:沈陽農業大學,2018:9.
[8]劉艷霞,沈 宏,李 想,等. 煙草青枯病勞爾氏菌與拮抗菌對根系分泌物的競爭作用[J]. 微生物學報,2020,60(2):333-348.
[9]秦秋菊,高希武. 昆蟲取食誘導的植物防御反應[J]. 昆蟲學報,2005,48(1):125-134.
[10]余源嬋,楊茂發,商勝華,等. 不同昆蟲取食對煙草信號分子和防御酶的影響[J]. 環境昆蟲學報,2021,43(4):967-977.
[11]胡淑霞. 茶樹病蟲害發生的相互關系[J]. 茶葉通訊,1998,25(3):26-27.
[12]介曉磊,劉世亮,李有田,等. 不同濃度鈣營養液對煙草礦質營養吸收與積累的影響[J]. 土壤通報,2005,36(4):560-563.
[13]唐世凱,劉麗芳,李永梅. 水培條件下烤煙間作草木樨對煙株氮磷鉀含量的影響[J]. 云南農業大學學報,2009,24(3):380-384.
[14]張成省. 煙草根系分泌物介導的黑脛病抗性機制研究[D]. 北京:中國農業科學院,2020:13.
[15]伍彥容,黃瑞然,張兆霞,等. 蒜頭中SOD的提取研究[J]. 山東化工,2015,44(12):29-31.
[16]張志清,叢 軍,林 虹,等. 麥苗中超氧化物歧化酶抗氧化活性研究[J]. 中國糧油學報,2014,29(7):12-17.
[17]王孝濤,李淑芹,許景鋼,等. 生物肥對大豆根際過氧化氫酶和脲酶活性的影響[J]. 東北農業大學學報,2012,43(5):96-99.
[18]趙龍飛,徐亞軍,賴心河,等. 內生細菌252和254對鹽脅迫下小麥幼苗過氧化物酶和過氧化氫酶活性的影響[J]. 應用生態學報,2017,28(9):2984-2992.
[19]Zhang Q M,Zhu L S,Wang J,et al. Oxidative stress and lipid peroxidation in the earthworm Eisenia fetida induced by low doses of fomesafen[J]. Environmental Science and Pollution Research,2013,20(1):201-208.
[20]張志良,瞿偉菁. 植物生理學實驗指導[M]. 3版.北京:高等教育出版社,2003:127-180.
[21]Dunn W B,Broadhurst D,Begley P,et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry[J]. Nature Protocols,2011,6:1060-1083.
[22]Want E J,Wilson I D,Gika H,et al. Global metabolic profiling procedures for urine using UPLC-MS[J]. Nature Protocols,2010,5:1005-1018.
[23]徐周洋. 人參根系分泌物對其病原菌的化學趨向性的影響及人參屬植物種子種苗的化感作用研究[D]. 長春:吉林農業大學,2021:8.
[24]吳 瓊,方吳云,王文杰. 機械損傷對茶樹苯丙氨酸解氨酶活性及茶多酚含量的影響[J]. 安徽農業科學,2016,44(5):11-14.
[25]Li Y D,Duan T Y,Li Y Z. Research progress in the interactions of fungal pathogens and insect pests during host plant colonization[J]. Journal of Plant Diseases and Protection,2021,128(3):633-647.
[26]Shi Y,Ma T T,Zhang Z Y,et al. Foliar herbivory affects the rhizosphere microbial assembly processes and association networks[J]. Rhizosphere,2023,25:100649.
[27]高佳敏,高素紅,高寶嘉. 昆蟲取食和機械損傷對葡萄葉片代謝物的影響[J]. 西北農業學報,2019,28(9):1543-1551.
[28]張寬朝,金 青,蔡永萍,等. 苯丙氨酸解氨酶與其在重要次生代謝產物調控中的作用研究進展[J]. 中國農學通報,2008,24(12):59-62.
[29]Li S L,Xu C,Wang J,et al. Cinnamic,myristic and fumaric acids in tobacco root exudates induce the infection of plants by Ralstonia solanacearum[J]. Plant and Soil,2017,412(1/2):381-395.
[30]董 艷,董 坤,鄭 毅,等. 不同抗性蠶豆品種根系分泌物對枯萎病菌的化感作用及根系分泌物組分分析[J]. 中國生態農業學報,2014,22(3):292-299.
[31]楊小環,王立印,李宏鑫,等. 蕎麥、高粱根系分泌物對玉米根邊緣細胞和根生長的影響[J]. 生態學報,2023,43(9):3778-3788.
[32]黃玉茜,韓曉日,楊勁峰,等. 花生根系分泌物對土壤微生物學特性及群落功能多樣性的影響[J]. 沈陽農業大學學報,2015,46(1):48-54.
