雜交3倍體泥鰍鰭細胞系染色體組構成研究
2024-09-29 00:00:00郭文軒周子昱馬可馨楊悅瑤李逸帆李雅娟周賀
安徽農業科學
2024年18期
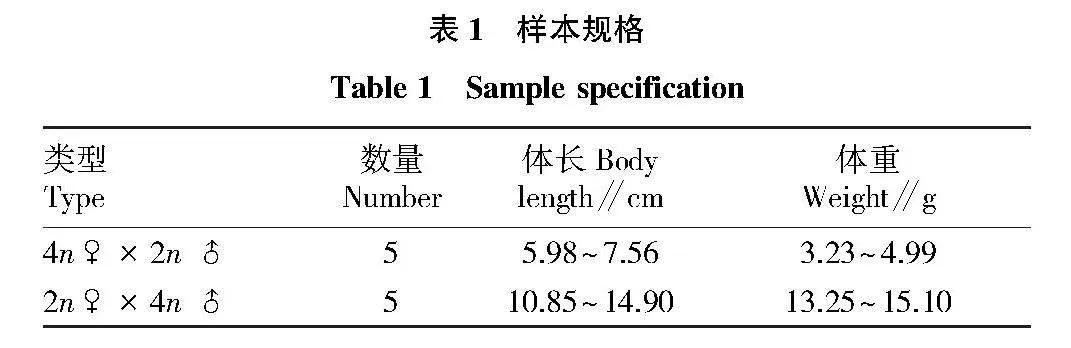
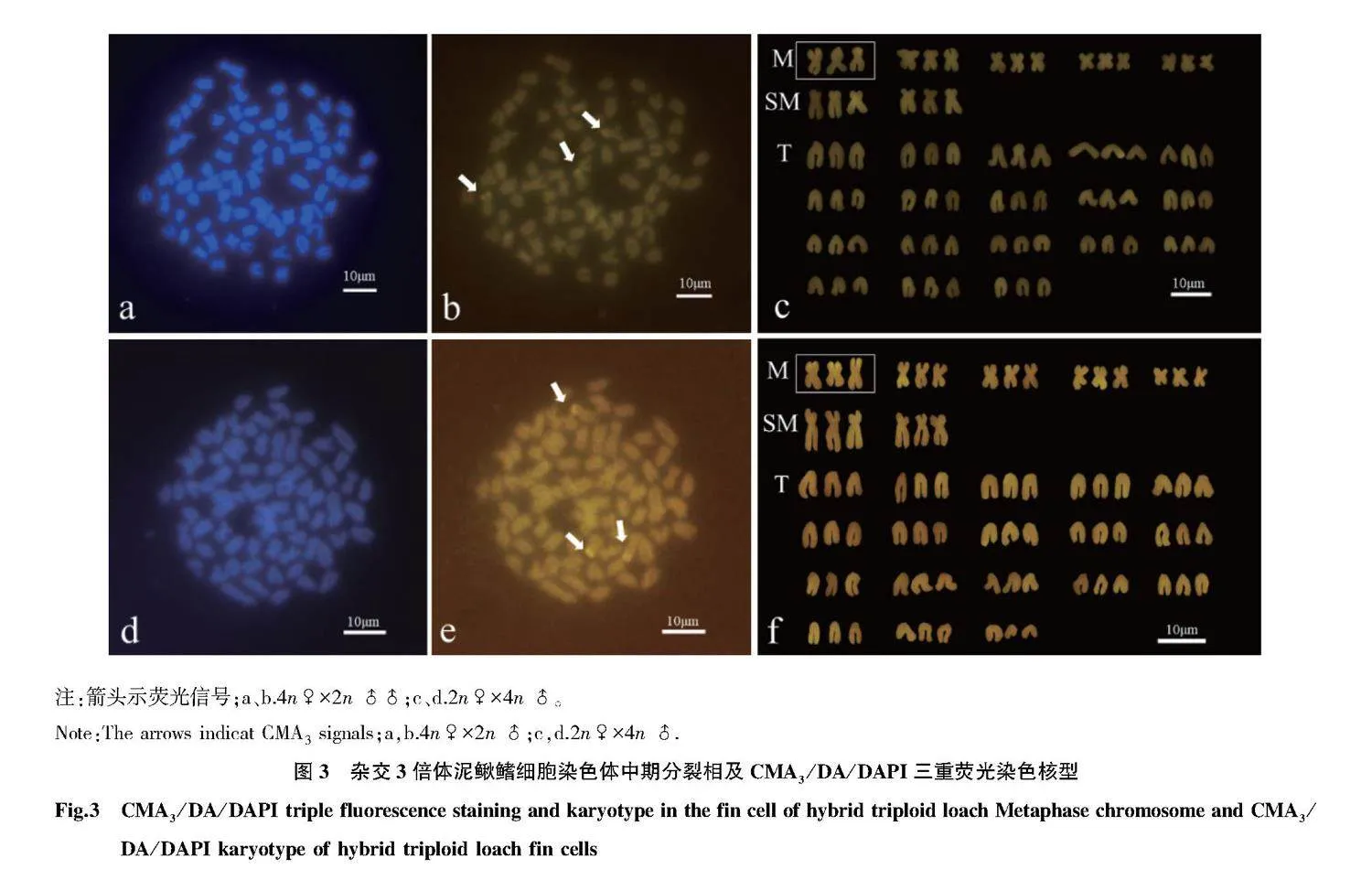
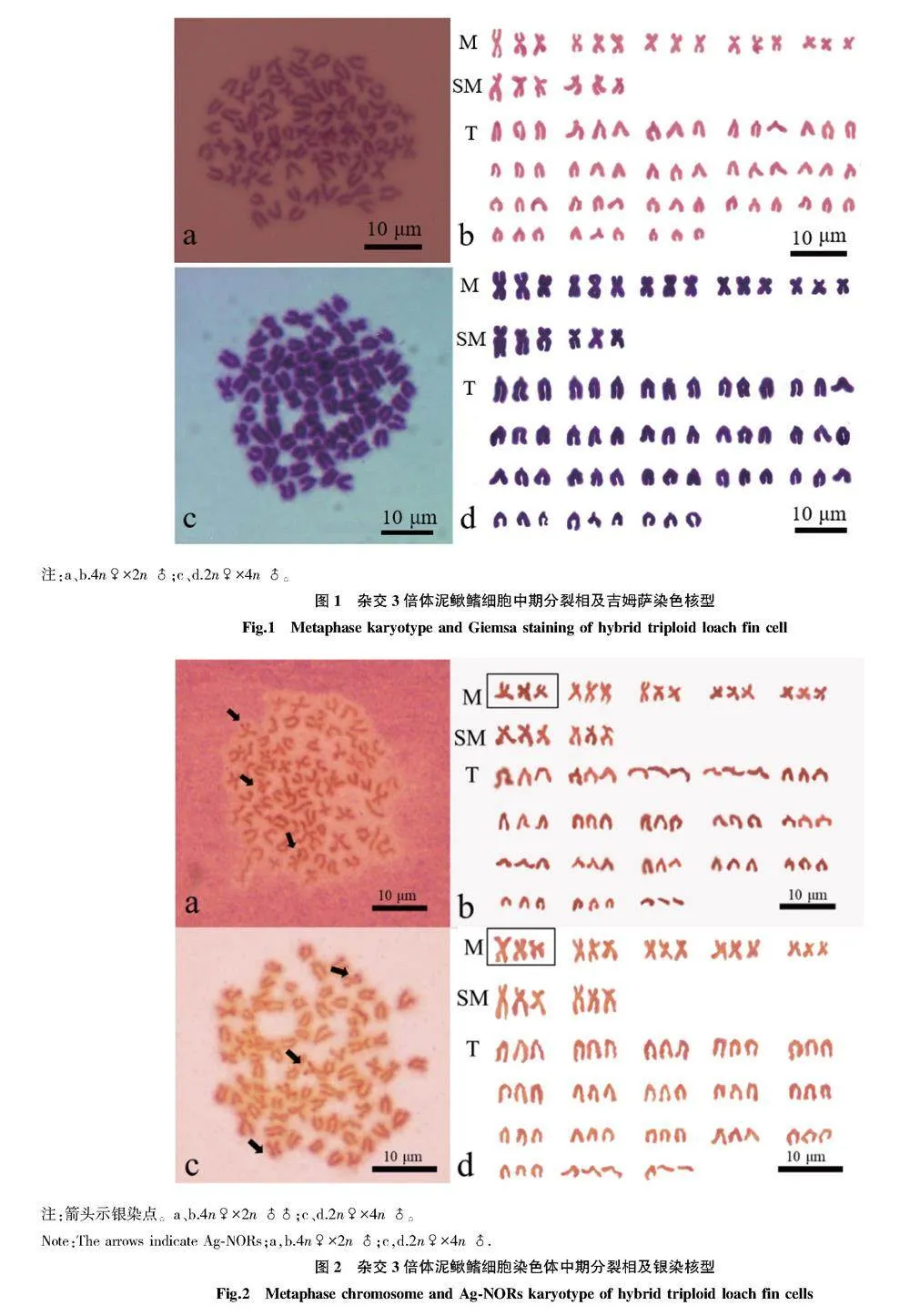
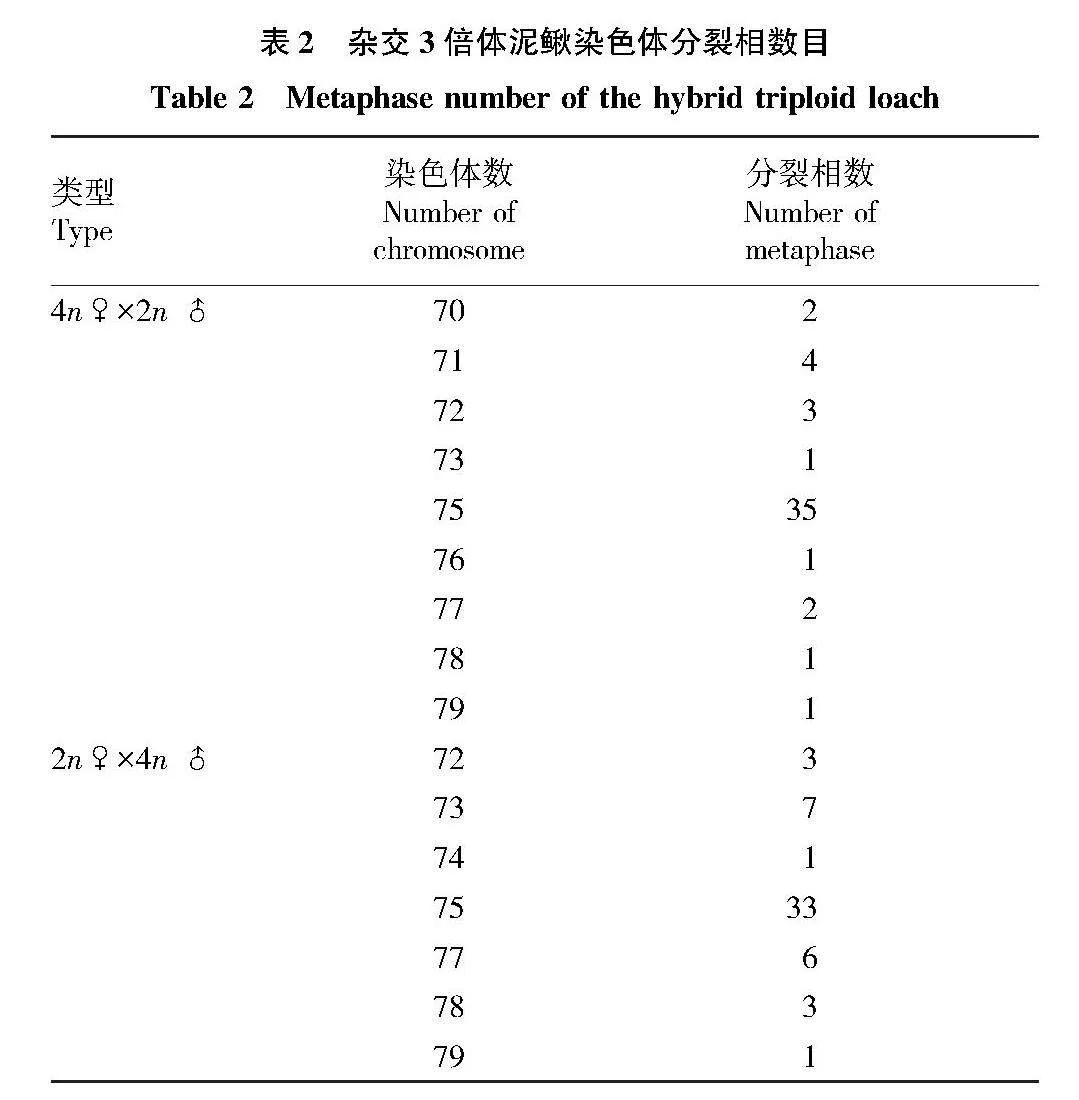
摘要 通過天然4倍體(4n=100)和2倍體(2n=50)泥鰍雜交創制不同子代雜交組合(4n♀×2n♂、2n♀×4n♂)。以傳代20次的雜交3倍體泥鰍鰭細胞系為材料,采用常規冷滴片法制備染色體標本,通過吉姆薩染色確定數目及核型;通過銀染法及CMA3/DA/DAPI三重熒光染色技術對其進行帶型分析。結果顯示:正反交3倍體鰭細胞系的染色體分裂相數目為3n=75,核型公式15m + 6sm + 54t,NF=96;正反交3倍體泥鰍鰭細胞系的染色體分裂相中顯示有3個銀染點,均位于第1組中部著絲粒染色體(M1)短臂的端部區域;正反交3倍體泥鰍鰭細胞的染色體中期分裂相中顯示有3個CMA3熒光信號點,均位于M1區域。研究表明:雜交3倍體泥鰍鰭細胞經細胞培養傳代20次后的染色體組成未發生改變,其染色體數目、核型和帶型與3倍體泥鰍其他體細胞結果一致。
關鍵詞 雜交3倍體泥鰍;鰭細胞系;核型分析;銀染;CMA3/DA/DAPI
中圖分類號 S966.4 文獻標識碼 A
文章編號 0517-6611(2024)18-0067-04
doi:10.3969/j.issn.0517-6611.2024.18.015
開放科學(資源服務)標識碼(OSID):
Study on Chromosome Composition of Hybrid Triploid Loach (Misgurnus anguillicaudatus) Fin Cell Line
GUO Wen-xuan,ZHOU Zi-yu,MA Ke-xin et al
(College of Aquatic and Life Science,Dalian Ocean University,Dalian,Liaoning 116023)
Abstract In this study,a natural tetraploid loach (4n=100) was crossed with a diploid loach (2n=50) thus creating the different offspring hybrid combinations (4n♀×2n♂,2n♀×4n♂).Chromosome samples of hybrid triploid loach fin cell passed for 20 times were prepared via conventional cold drop method.The chromosome number and karyotype were determined via Giemsa staining; Banding analysis were conducted via Ag-NORs and CMA3/DA/DAPI staining.The results showed that the metaphase chromosome number of the fin cells of the hybrid triploid loach was 3n=75,and the karyotype formula was 15m+6sm+54t,NF=96;three silver staining signal points were observed in the chromosome metaphase of hybrid triploid loach fin cells,located at the end region of the first pair of central centromeric chromosomes (M1);there were three CMA3 signal points in the chromosome metaphase of hybrid triploid loach fin cells,which were located at the region of chromosomes (M1).The study showed that the chromosome composition of hybrid triploid loach fin cells remained unchanged after 20 times of cell culture passage,and the chromosome number,karyotype and band type were consistent with other somatic chromosomes of triploid loach.The results laid the foundation for the cytogenetics of hybrid triploid loach and provided a theoretical basis for cytogenetics of triploid loaches.
Key words Hybrid triploid loach;Fin cell line;Karyotype analysis;Ag-NORs;CMA3/DA/DAPI
基金項目 國家自然科學基金項目(31272650)。
作者簡介 郭文軒(1999—),女,遼寧葫蘆島人,碩士研究生,研究方向:水產遺傳育種與繁殖。*通信作者,副教授,博士,碩士生導師,從事水產動物繁殖育種研究。
收稿日期 2023-10-24
泥鰍(Misgurnus anguillicaudatus)隸屬于鯉形目(Cypriniformes)鰍科(Cobitidae)泥鰍屬(Misgurnus),是我國水產養殖行業的潛力產品[1]。據報道,泥鰍存在自然多倍化現象,在我國的天然水域中已發現5種不同倍性的泥鰍種類,包括2倍體(2n=50)、3倍體(3n=75)、4倍體(4n=100)、5倍體(5n=125)和6倍體(6n=150)[2-4]。多倍體泥鰍普遍具有個體大、生長快及適應性強等優良的經濟性狀,具有很大的育種潛力[5]。通過化學或物理等誘導方法培育3倍體存在價格較貴,成功率較低等問題,而4倍體與2倍體泥鰍雜交的方式可以穩定高效地獲得3倍體[6-8]。……
登錄APP查看全文
