灰氈毛忍冬ERF 基因家族鑒定及其在衰老過程中的表達分析
2024-09-29 00:00:00劉燕妮曹正艷吳佩蔭李清清陳澤雄唐寧
經濟林研究 2024年2期
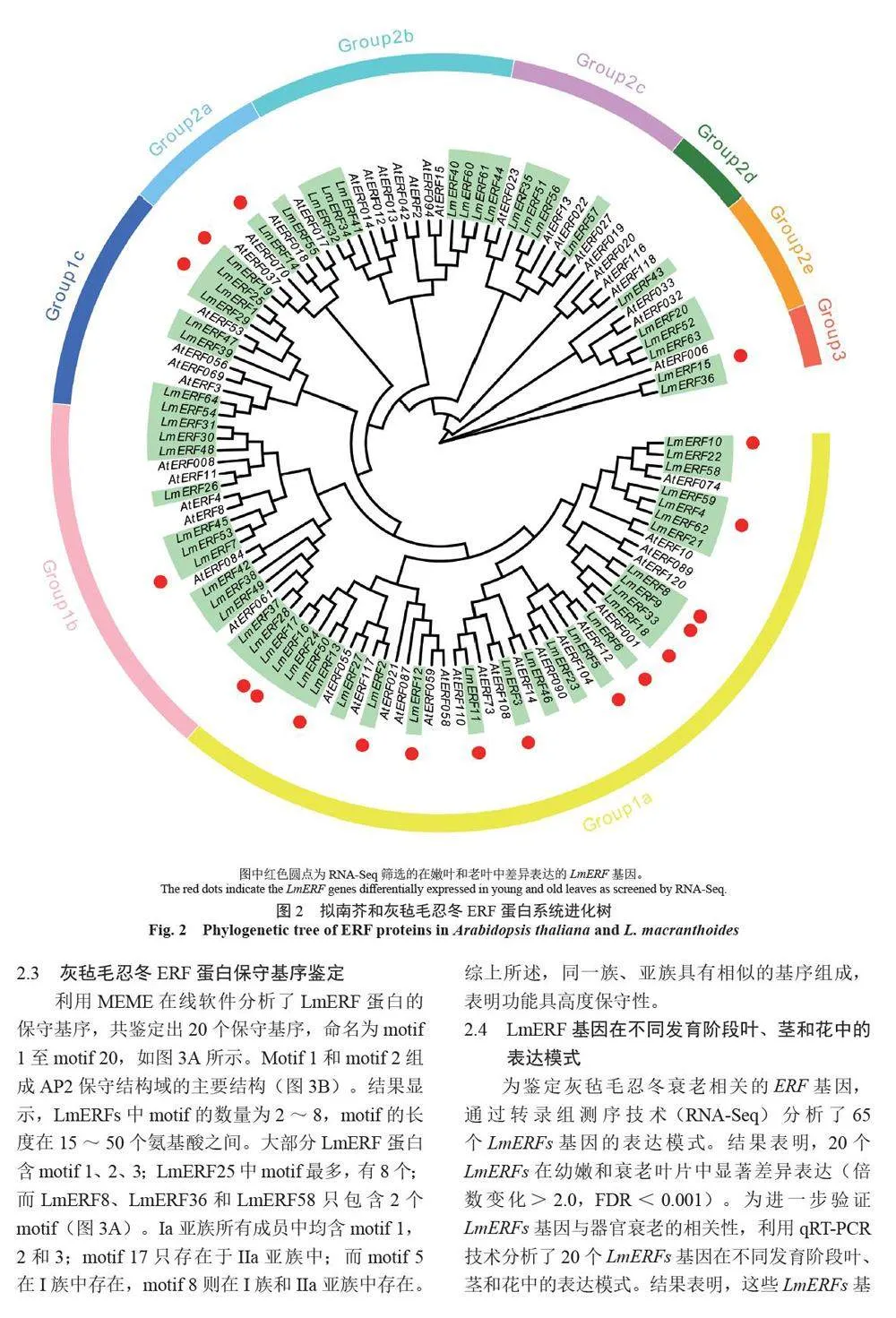
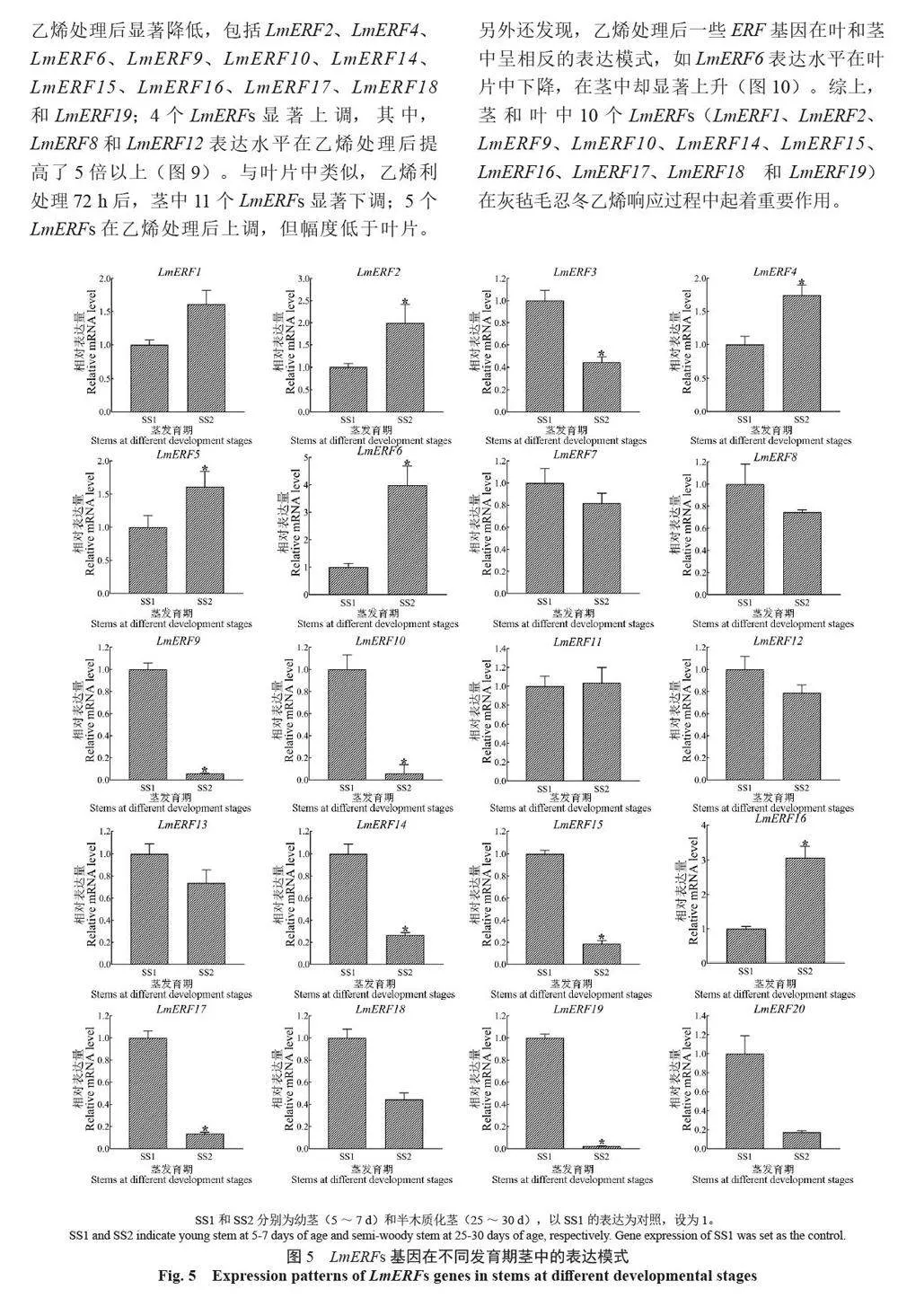
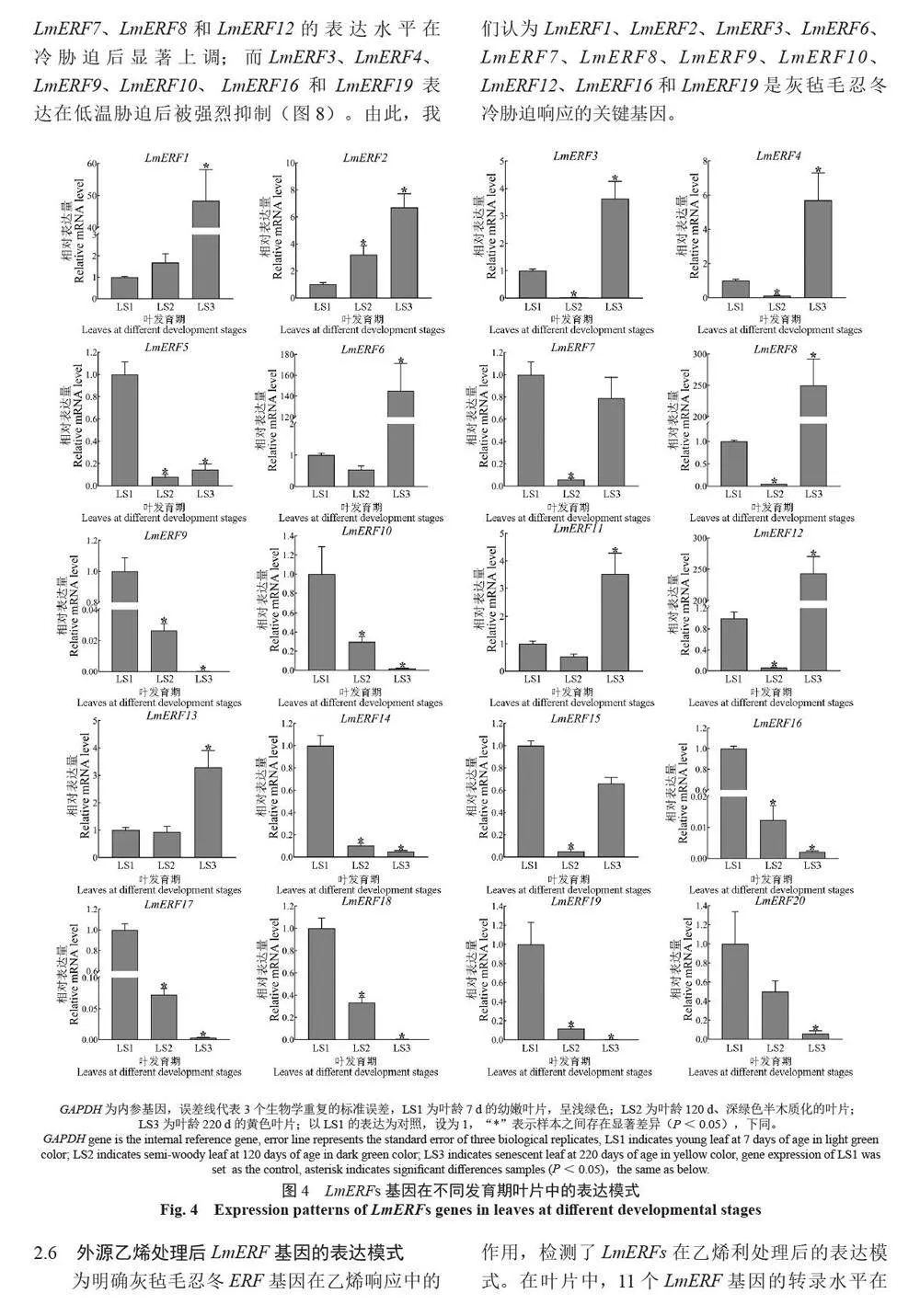
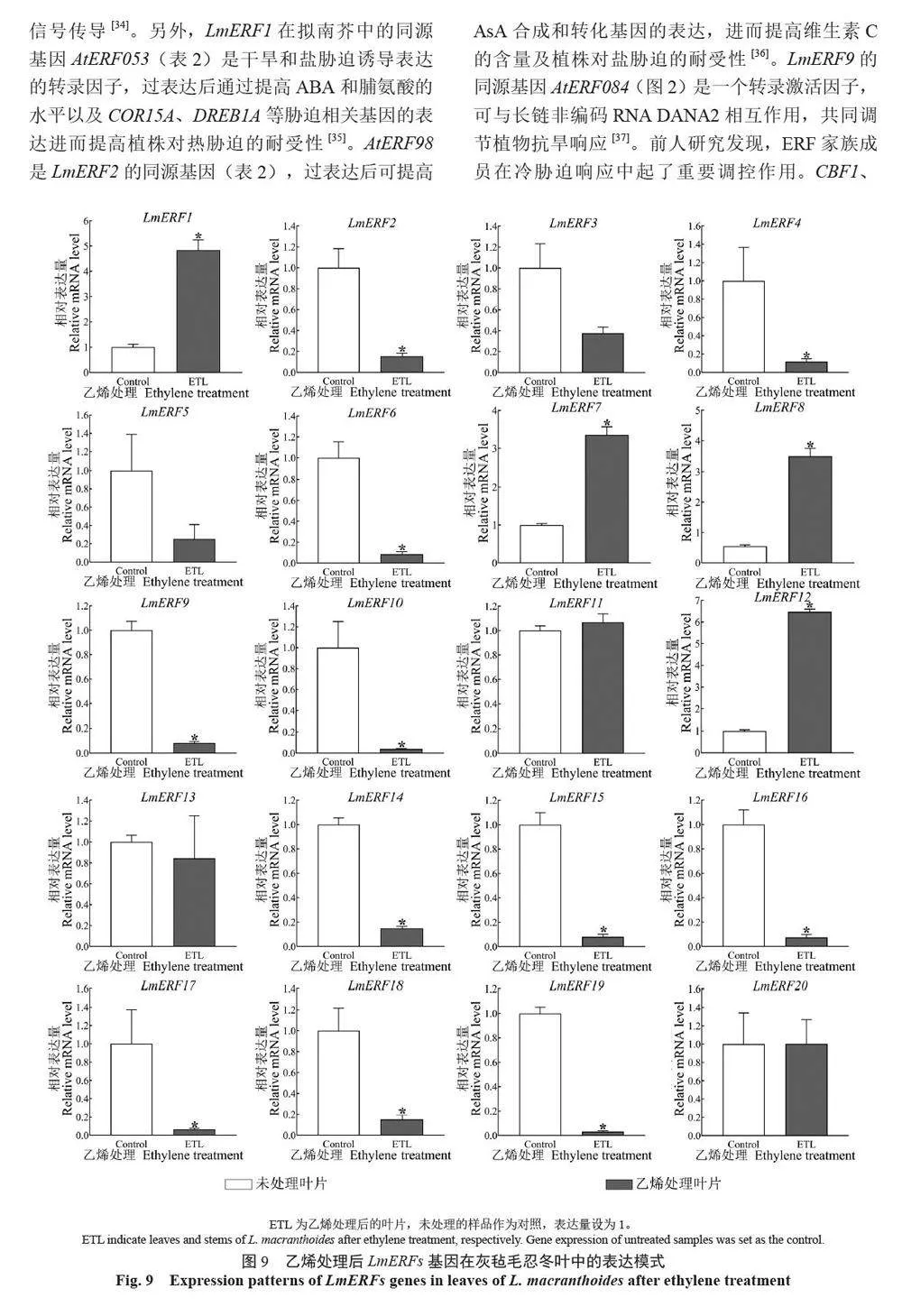
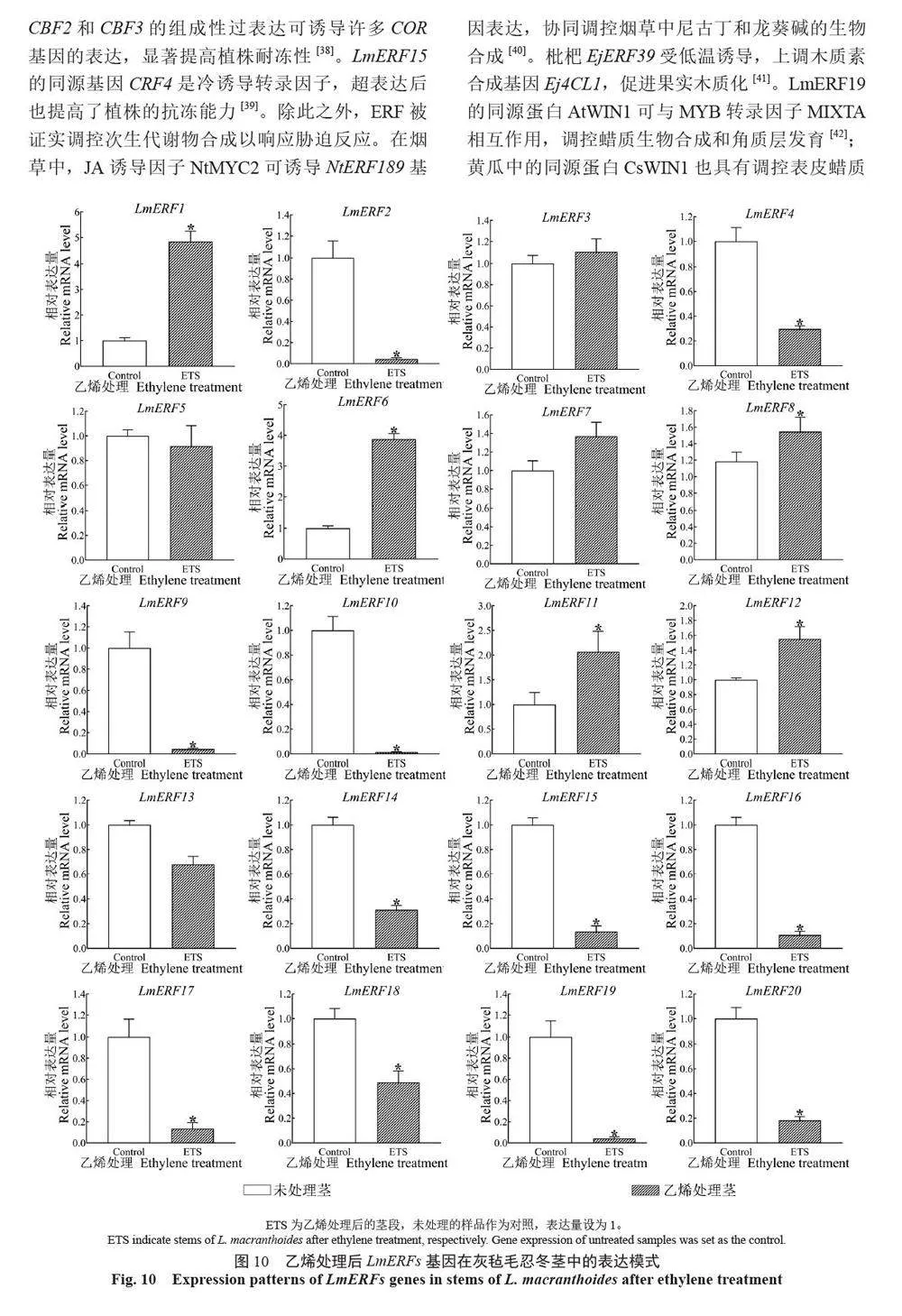
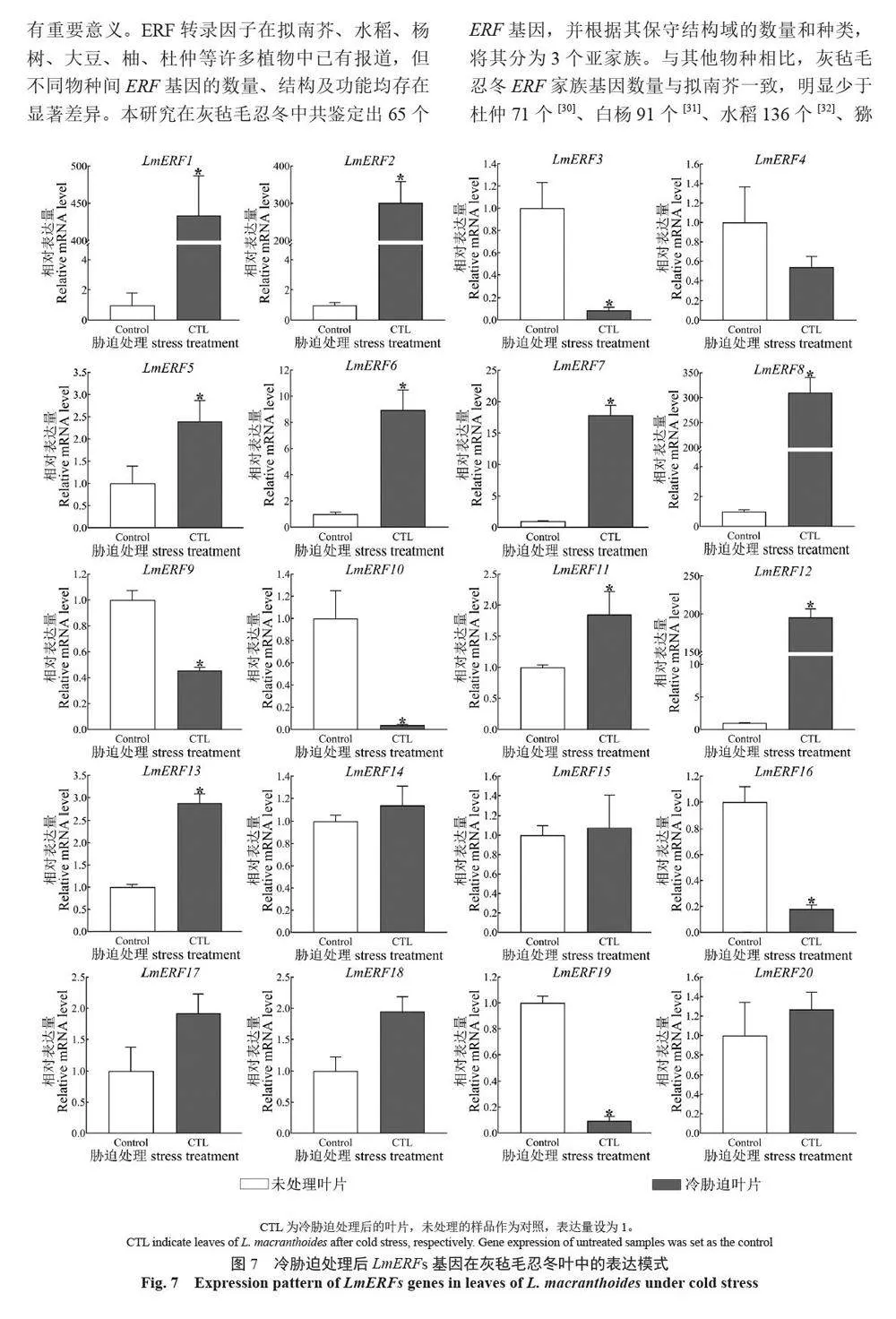
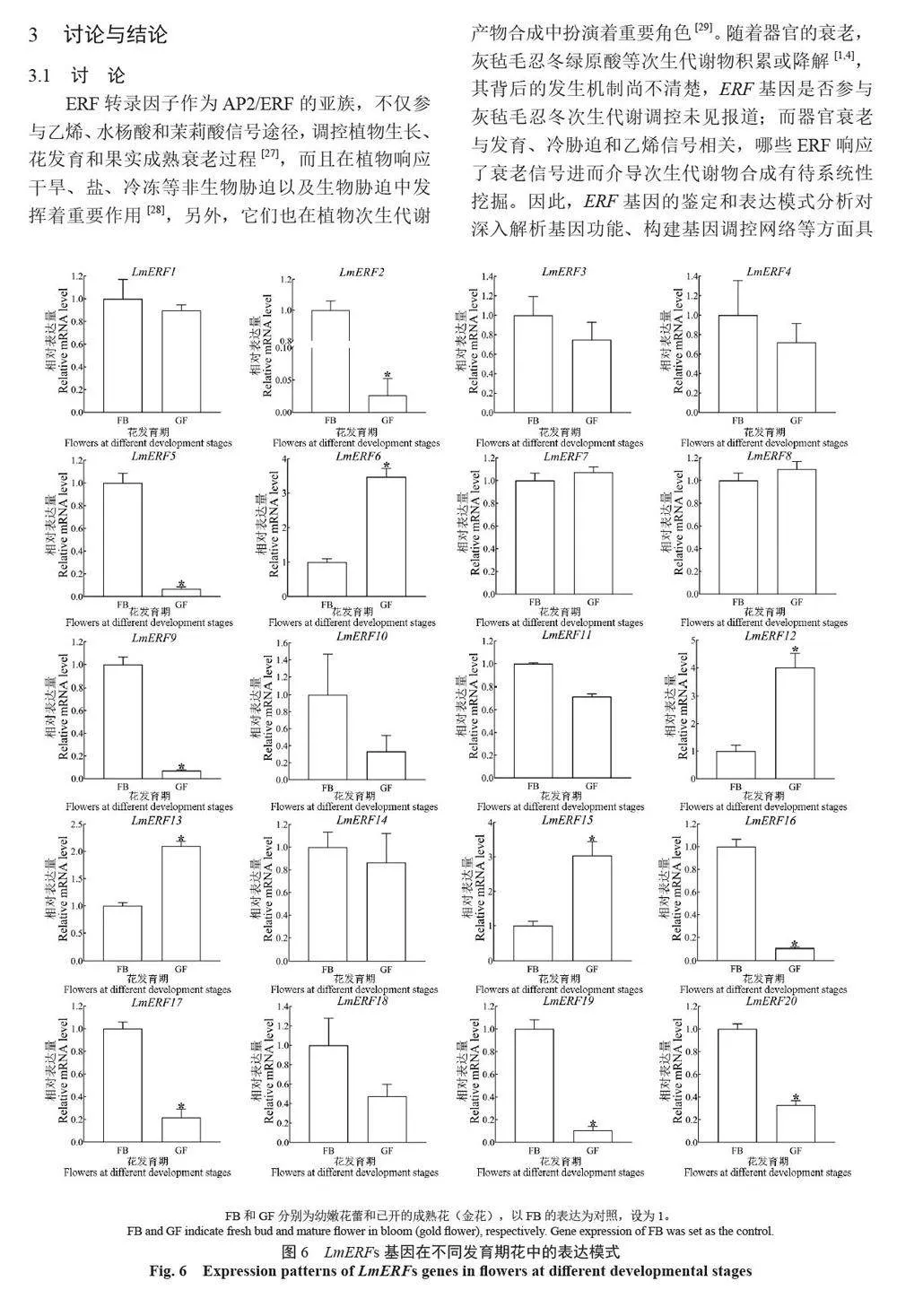
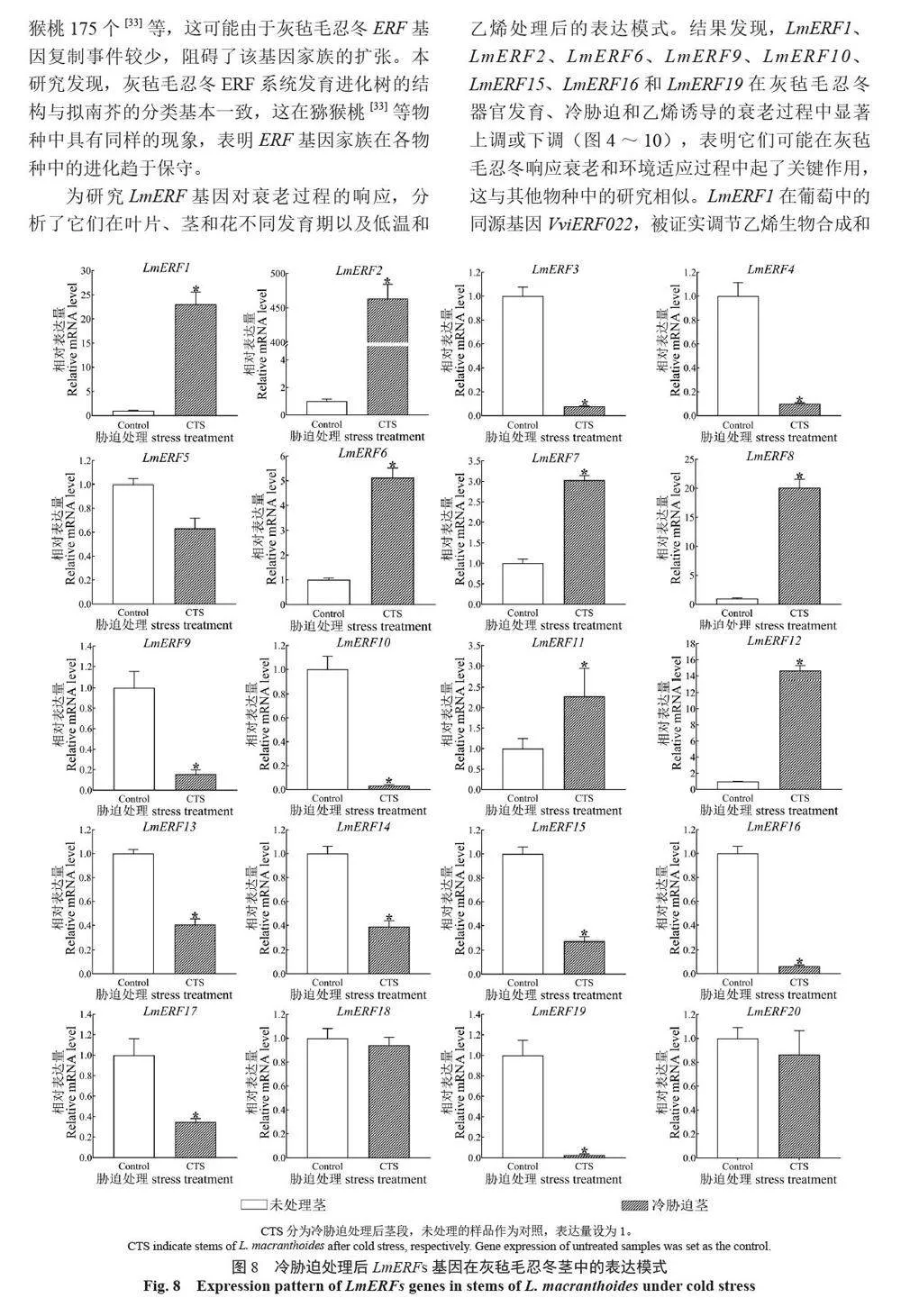


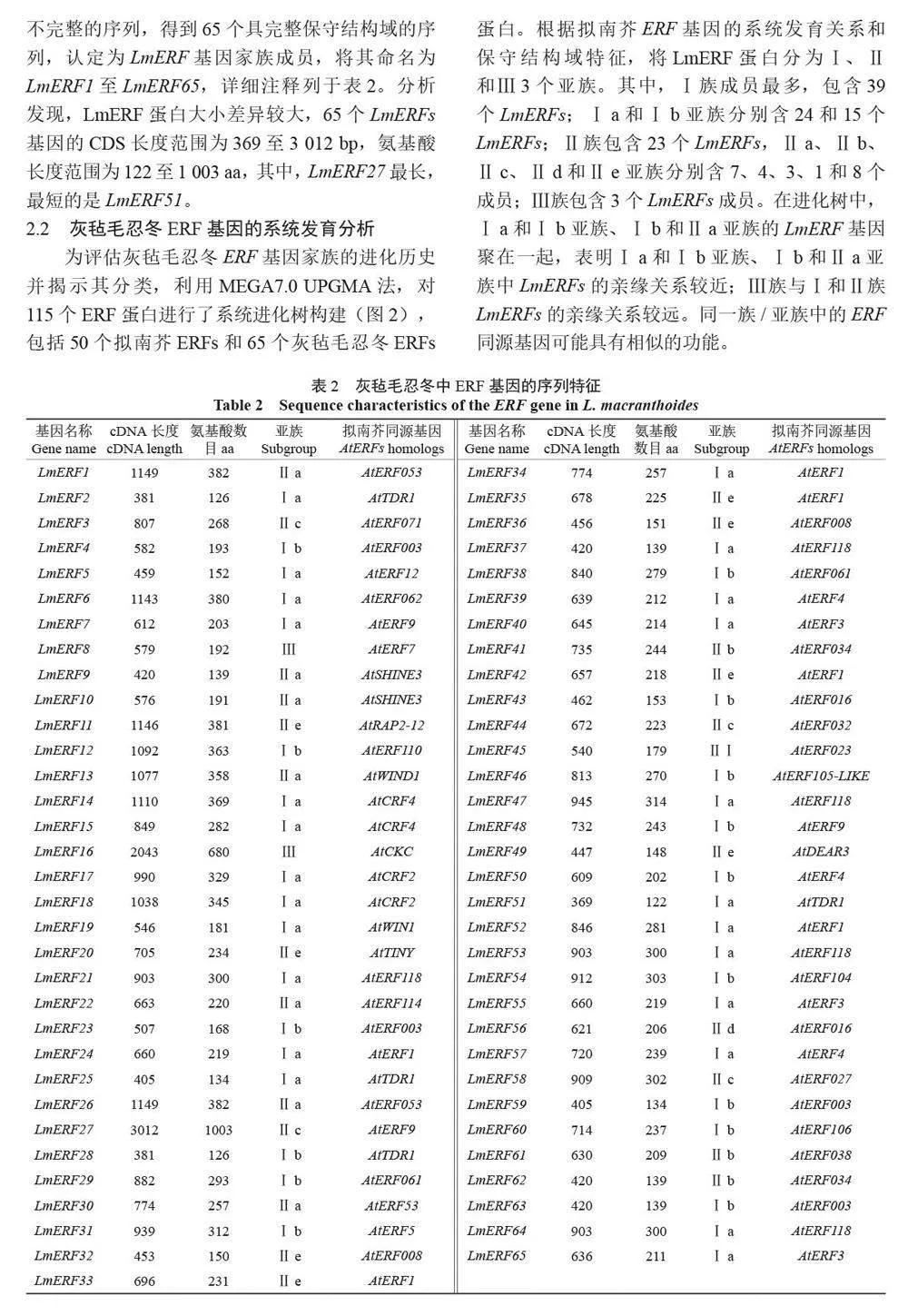
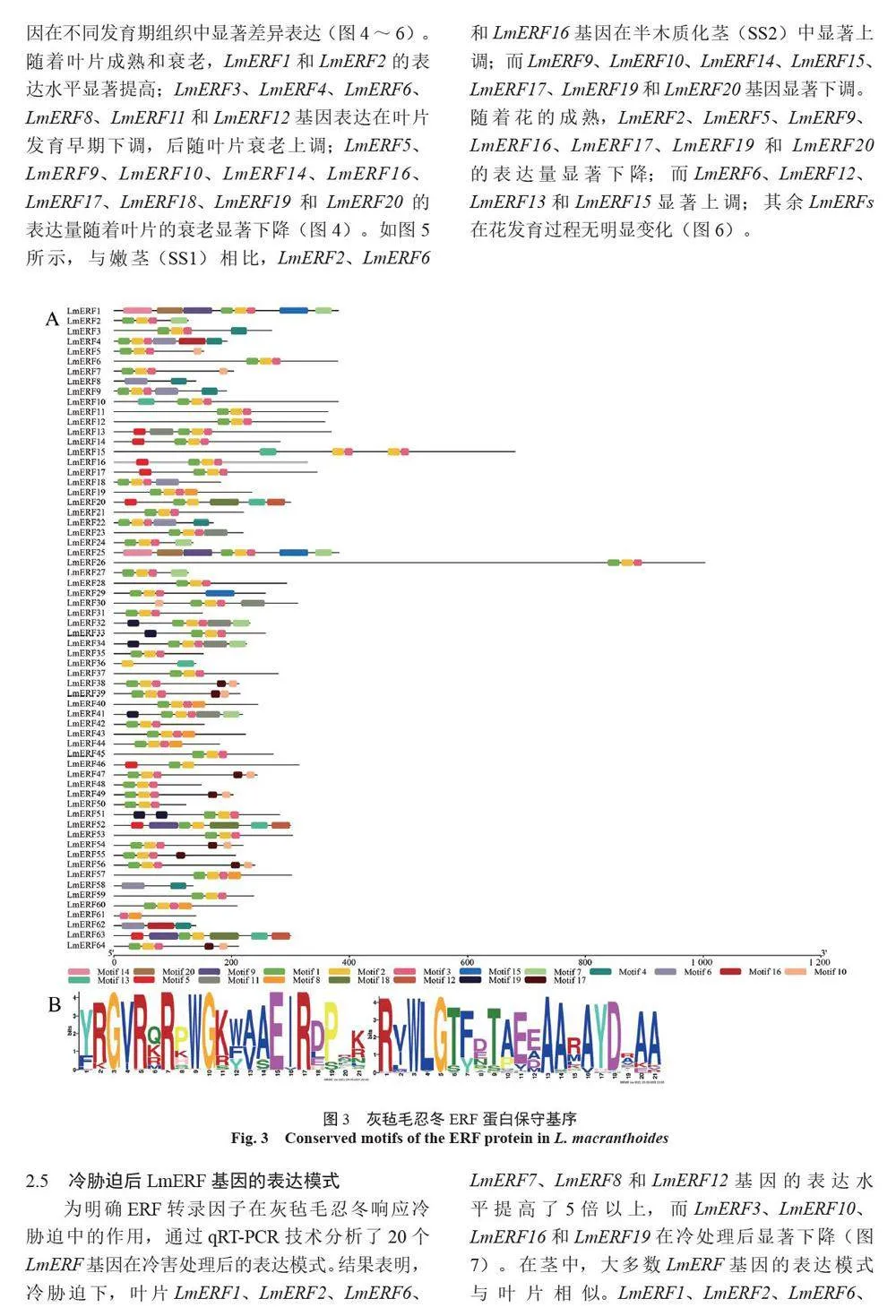

摘 要:【目的】灰氈毛忍冬Lonicera macranthoides 為我國南方地區大宗道地中藥材和重要經濟植物,綠原酸和木犀草苷等藥效物質在器官衰老過程中積累且受轉錄因子調控。乙烯信號轉導相關ERF 基因家族廣泛參與植物的衰老、脅迫響應和次生代謝。鑒定灰氈毛忍冬ERF 家族成員并探究其表達模式,可為該類基因調控次生代謝的功能研究提供參考。【方法】基于轉錄組數據鑒定灰氈毛忍冬ERF 轉錄因子家族,通過系統發育樹分析其進化關系,利用MEME 在線網站分析序列的結構基序。QRT-PCR 分析LmERFs 基因在不同器官衰老過程以及冷脅迫和乙烯處理后的表達模式。【結果】在灰氈毛忍冬中共鑒定出65 個具AP2 保守結構域的LmERFs 基因,可將其分為3 個亞組。利用RNA-Seq 技術篩選了在幼嫩和衰老葉片之間差異表達的20 個LmERFs。進一步qRT-PCR 分析顯示,分別有18 個、13 個和11 個LmERFs 基因在葉片、莖和花的衰老過程顯著差異表達,其中LmERF2、LmERF9、LmERF16、LmERF17 和LmERF19 基因表達在不同器官衰老過程中均具顯著差異。冷脅迫后,葉和莖中7 個LmERFs(LmERF1、LmERF2、LmERF6、LmERF7、LmERF8、LmERF11 和LmERF12) 顯著上調,LmERF3、LmERF9、LmERF16 和LmERF19 顯著下調;而乙烯處理誘導了LmERF1、LmERF8 和LmERF12基因的表達,抑制了LmERF2、LmERF4、LmERF9、LmERF10、LmERF14、LmERF15、LmERF16、LmERF17、LmERF18 和LmERF19 基因的表達。【結論】8 個LmERFs(LmERF1、LmERF2、LmERF6、LmERF9、LmERF10、LmERF15、LmERF16 和LmERF19)響應了器官發育、冷脅迫和乙烯誘導的衰老過程,這為后續挖掘高效調控灰氈毛忍冬次生代謝的靶點LmERFs 提供了數據支持。
關鍵詞:灰氈毛忍冬;ERF 基因家族;器官衰老;乙烯;冷害脅迫
中圖分類號:S567.79 文獻標志碼:A 文章編號:1003—8981(2024)02—0013—16
灰氈毛忍冬Lonicera macranthoides Hand-Mazz又名大金銀花,忍冬科忍冬屬植物,為我國西南地區大宗道地中藥材和特色經濟植物,其次生代謝產物如綠原酸和木犀草苷等核心藥效成分具有抗病毒、抗氧化、增強免疫力等多種功能[1]。目前,灰氈毛忍冬已被廣泛應用于制藥、保健品及化妝品等行業,市場巨大[2]。然而,灰氈毛忍冬存在品種混雜、藥效成分含量參差不齊等問題,已成為制藥、提取、加工等下游產業發展的瓶頸,挖掘調控次生代謝產物生物合成的關鍵因子是解決這一難題的重要途徑。
研究表明,植物通過產生次生代謝物響應衰老信號、適應環境壓力[3]。擬南芥葉片衰老過程中,花青素和吲哚衍生的甲基葡萄糖苷顯著增加;而黃酮醇苷和芥子酸共軛物持續減少。隨著器官的發育和衰老,灰氈毛忍冬綠原酸、綠原酸前體及異構體等苯丙烷類物質和木犀草苷等黃酮類物質含量發生顯著變化[1,4]。Rezaie 等[5] 發現,冷脅迫顯著影響了羅勒中苯丙素類化合物的含量。糖和次生代謝在柑橘類植株響應冷脅迫中發揮著重要作用[6-7]。乙烯可促進植物次生代謝物的產生,如長春花中酚酸、黃酮醇、萜類和吲哚類生物堿、烏龍茶中兒茶素和甜橙中瓦倫西亞橘烯[8]。由此可知,植物發育、激素或脅迫信號顯著影響次生代謝物合成,探究灰氈毛忍冬響應衰老和脅迫機制有助于挖掘次生代謝物合成的關鍵基因,為利用基因工程手段培育新品種提供理論依據。
轉錄因子通過介導次生代謝物合成參與生物和非生物應激反應[9]。AP2/ERF 轉錄因子是植物最大的轉錄因子家族之一,成員含有1 個或2 個由60 ~ 70 個氨基酸構成的AP2 保守結構域,根據結構域數量和識別序列的不同可將其分為AP2、ERF、DREB、RAV 和Soloist5 個亞家族[10],廣泛參與植物生長發育、初生和次生代謝調控、生物或非生物脅迫響應[11] 以及乙烯等植物激素信號傳導途徑[12-13]。其中ERF(ethylene response factor) 轉錄因子亞家族是乙烯信號通路下游的關鍵調控因子[14-15],只含有一個AP2 結構域,第14 位和第19 位氨基酸分別是丙氨酸和天冬氨酸,區別于其他亞家族成員。ERF 亞家族成員可特異性結合基因啟動子區的GCC-box 元件(AGCCGCC),從而參與調控乙烯應答、生物及非生物脅迫和次生代謝[13,16]。胡俊東等[17] 研究表明,PheERF5 基因參與毛竹抗生物和非生物脅迫過程;小麥TaERF1基因在擬南芥中過表達可提高轉基因植株對高鹽和低溫的耐受性[18];ERF- Ⅵ亞族成員CRF 具有正向調節植物滲透脅迫耐受性和抗凍性作用[18-19]。Lyu 等[20] 發現樺木中BpERF13 過表達可通過結合并上調冷脅迫關鍵轉錄因子CBF 的基因表達來增強耐寒性。擬南芥ERF 亞族轉錄因子ORA59基因受茉莉酸和乙烯的協同誘導,通過正向調控PDF1.2 的表達增加植物灰霉抗性,在茉莉酸與乙烯信號的交叉對話中起了重要的調控作用[21-22]。煙草中過表達NtERF19 可顯著增加生物堿,特別是新煙堿的含量[23]。目前,鮮見忍冬屬藥用植物ERF 轉錄因子的研究,在灰氈毛忍冬中尚未見ERF 參與脅迫響應的研究報道。
