波形蛋白糖基化突變質粒對PC12細胞分化的影響研究*
2024-05-09 09:07:36邵曉婷戴玉璇應玲靜陳美仙
重慶醫學 2024年8期
邵曉婷,戴玉璇,應玲靜,陳美仙
(浙江大學醫學院附屬金華醫院/金華市中心醫院兒科,浙江金華 321000)
蛋白糖基化是將聚糖基團添加到蛋白分子中[1],通過調節蛋白分子的結構、穩定性及功能等,使其在生物體內多種生理及病理過程中發揮重要作用[2-3]。糖基化與機體內許多生物學作用及多種疾病息息相關[4-5],糖基化修飾可以影響細胞的許多生理特性,例如分子間相互作用、識別及后續的生物調節功能等。外源性寡糖化合物(Cyclo-ManN propanyl perac,Cyclo-ManN pro)可以使糖復合物發生糖基化修飾。糖基化修飾在大鼠腎上腺嗜鉻細胞(PC12細胞)分化中發揮重要調節作用[6],而糖基化作用促進PC12細胞分化的具體機制尚不明確,作者前期實驗中發現波形蛋白(vimentin)是糖代謝分子作用PC12細胞的重要中間蛋白。波形蛋白是一種潛在的糖蛋白,有3個潛在糖基化位點(7:絲氨酸;33:蘇氨酸;34:絲氨酸),為了進一步研究糖基化在PC12細胞中的作用,本研究構建波形蛋白糖基化位點突變質粒,并研究其對PC12細胞分化的影響。現報道如下。
1 材料與方法
1.1 材料
PC12細胞購于中國科學院上海分院;Cyclo-ManN pro受贈于WERMER RUETTER教授;引物合成由上海生工生物工程公司完成;pcDNA3.1b購于美國Invitrogen公司;PCR試劑PrimeSTAR、dNTP購于天根生化科技(北京)有限公司,M-MLV逆轉錄酶購于普洛麥格(北京)生物技術有限公司;限制性內切酶(EcoRⅠ/XbaⅠ)、四碘甲狀腺原氨酸(T4)連接酶購于日本TaKaRa公司;大腸桿菌菌株Top10來源于復旦大學上海醫學院生化系存種;凝膠電泳試劑購于法國Biowest公司;DNA標記物購于天根生化科技(北京)有限公司;RNA抽提試劑盒購于上海星漢生物科技有限公司,膠回收試劑盒購于安徽省優晶生物工程有限公司,質粒抽提試劑盒購于天根生化科技(北京)有限公司;超凈工作臺購于上海凈化設備有限公司;倒置顯微鏡購于安徽安慶市醫療器械有限公司;熒光顯微鏡購于日本Olympus公司;核酸電泳槽購于北京六一儀器廠。
1.2 方法
1.2.1構建波形蛋白相關質粒
目的基因片段獲取:PC12細胞RNA抽提,獲取波形蛋白的mRNA,通過逆轉錄PCR(reverse transcription PCR,RT-PCR),獲取波形蛋白的cDNA,設計引物,PCR分別獲取波形蛋白正常基因片段和糖基化突變基因片段。重組質粒建成與分析:分別將pcDNA3.1b與波形蛋白雙酶切(EcoRⅠ/XbaⅠ)后連接,轉進大腸桿菌細胞,篩選氨芐青霉素陽性克隆菌,培養,抽提質粒并電泳鑒定,送于上海生工生物工程公司進行測序。RT-PCR反應體系:RNA 2 μg、5×RNA PCR Buffer 4 μL、MgCl24 μL、10 mmol/L dNTP Mix 8 μL、Oligo dT 1 μL、MML enzyme 1 μL、DEPC超純水加至20 μL。反應條件:預變性65 ℃ 5 min,逆轉錄酶37 ℃ 1 h,99 ℃ 5 min,5 ℃ 5 min,-20 ℃保存。PCR反應體系:Template 0.5 μL、Primer(sense) 0.5 μL、Primer(Antisense) 0.5 μL、10×PCR Buffer 2.5 μL、2.5 mmol/L dNTP 1 μL、PrimeSTAR 0.5 μL、超純水 19.5 μL。PCR反應條件:波形蛋白(98 ℃ 5 min,98 ℃ 30 s,67 ℃ 30 s,72 ℃ 2 min,35個循環);波形蛋白糖基化位點突變(98 ℃ 5 min,98 ℃ 30 s,66 ℃ 30 s,72 ℃ 2 min,35個循環)。
1.2.2細胞培養及分化分析
(1)采用含10%胎牛血清DMEM培養基,在37 ℃、5% CO2的條件下培養PC12細胞,細胞密度約為1×104個/cm2時進行形態觀察。(2)細胞密度達到70%~80%時分別轉染不同質粒:對照組轉染空載質粒pcDNA3.1b,Vim組轉染波形蛋白正常基因片段重組質粒,Vim mut組轉染波形蛋白糖基化位點突變基因片段重組質粒。繼續保持37 ℃、5% CO2的條件下培養,分別在0、3 d時記錄細胞分化情況。(3)細胞分化情況分析:隨機選取3個不同的視野,不少于150個細胞數量,統計分化神經突平均長度與分化細胞占比(分化細胞為擁有至少1個神經突>10 μm的細胞)。
1.3 統計學處理
2 結 果
2.1 波形蛋白糖基化位點突變目的基因情況
在Uniprot數據庫中分析發現波形蛋白含有3個潛在糖基化位點(7:絲氨酸;33:蘇氨酸;34:絲氨酸),通過前后兩個片段將這3個位點全部突變,設計引物將位點19(T-G)、97(A-G)、100(T-G)突變,從而實現氨基酸序列突變[7:絲氨酸(S)變為丙氨酸(A);33:蘇氨酸(T)變為丙氨酸(A);34:絲氨酸(S)變為丙氨酸(A)]。前108 bp片段(正向引物:5′-AAA GAA TTC ATG TCC ACC AGG TCC GTG GCC TC-3′,反向引物: 5′-AAA GCG GGT GGC TGC GGT CAC ATA G-3′),后1 346 bp片段(正向引物:5′-AAA TAT GTG ACC GCA GCC ACC CGC-3′,反向引物:5′-AAA TCT AGA TAC TGC GCC GTT GCA CTG AGC CT-3′),突變全長基因片段(正向引物:5′-AAA GAA TTC ATG TCC ACC AGG TCC GTG GCC TC-3′,反向引物:5′-AAA TCT AGA TAC TGC GCC GTT GCA CTG AGC CT-3′),含EcoR Ⅰ酶切位點及Xba Ⅰ酶切位點。目的基因片段進行PCR擴增后進行凝膠電泳分析,波形蛋白基因約1 404 bp,凝膠電泳結果分析,分別于近100 bp(圖1A)、1 000~2 000 bp(圖1B)、1 000~2 000 bp(圖1C)處顯示目的片段,見圖1。

A:前108 bp片段電泳圖;B:后1 346 bp片段電泳圖;C:全長基因片段電泳圖;箭頭示目的片段。
2.2 波形蛋白糖基化突變重組質粒的鑒定
重組質粒為包含波形蛋白突變基因片段(約1 404 bp)的重組體,在同一電泳條件下,Vim mut組電泳速率慢于對照組,見圖2。
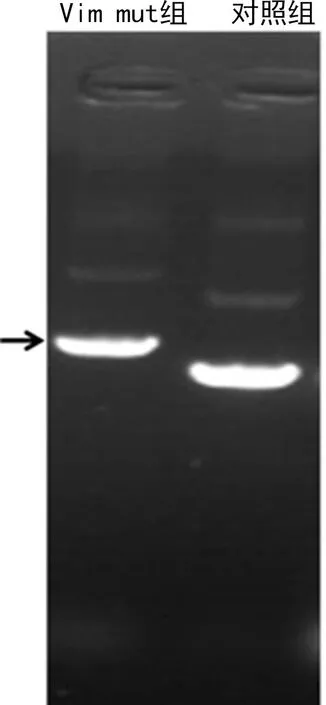
A箭頭示目的片段。
2.3 Cyclo-ManN pro處理前細胞分化分析
分別將空載質粒pcDNA3.1b、波形蛋白質粒、波形蛋白糖基化位點突變質粒轉染進PC12細胞,培養3 d觀察細胞分化情況(圖3A)。Vim組細胞平均神經突長度為(61.98±19.03)μm,高于對照組的(51.09±14.45)μm、Vim mut組的(51.49±14.78)μm,差異有統計學意義(P<0.05);Vim組分化神經突細胞百分比為(6.60±0.25)%,高于對照組的(4.27±0.18)%、Vim mut組的(4.76±0.33)%,差異有統計學意義(P<0.05),提示波形蛋白糖基化位點在PC12細胞分化中發揮重要作用,見圖3。

A:細胞形態分化圖(100×);B:細胞平均神經突長度定量結果圖;C:分化細胞百分比定量結果圖;a:P<0.05。
2.4 Cyclo-ManN pro處理后細胞分化分析
進一步觀察轉染上述3組質粒在Cyclo-ManN pro作用下對PC12細胞分化的影響。結果顯示,Cyclo-ManN pro處理后,Vim組細胞平均神經突長度為(78.01±18.31)μm,高于對照組的(69.98±12.85)μm、Vim mut組的(68.45±13.84)μm,差異有統計學意義(P<0.05);Vim組分化神經突細胞百分比為(10.62±0.25)%,高于對照組的(8.11±1.22)%、Vim mut組的(5.89±0.60)%,差異有統計學意義(P<0.05),提示Cyclo-ManN pro可能是通過波形蛋白糖基化位點作用而促進細胞分化的,將糖基化位點突變后,Cyclo-ManN pro促進神經分化的作用則會明顯降低,見圖4。
3 討 論
糖基化作用是一種重要的翻譯后修飾作用,可以參與多種生物代謝過程,其異常代謝會引起機體各種相關疾病[7-8]。翻譯后修飾可以擴大蛋白多樣性從而改變其生物功能,在許多生理和病理過程中發揮重要作用,例如細胞復制、細胞凋亡、轉錄調節、翻譯調節、信號轉導、免疫調節等[9-11]。在真核生物中有兩種主要蛋白糖基化類型:N連接型糖基化、O連接型糖基化。O連接型糖基化通常將聚糖分子連接到絲氨酸/蘇氨酸基團的羥基上[12]。波形蛋白含有3個糖基化位點(7:絲氨酸;33:蘇氨酸;34:絲氨酸),可能是一種潛在的O連接型糖基化蛋白。有研究顯示,O連接型糖基化在許多蛋白功能中發揮作用,包括蛋白細胞定位、蛋白穩定性、蛋白間相互作用等[13]。O連接型糖基化還與磷酸化修飾間有交互作用,可能是因為它們都可作用于蛋白分子的絲氨酸/蘇氨酸基團[14]。神經節苷脂的糖基化修飾可以通過影響細胞的存活與凋亡來參與早期腦發育[15]。唾液酸代謝前體物質可以使細胞中的糖復合物發生糖基化修飾作用[16-17]。
波形蛋白是一種Ⅲ型中間纖維蛋白,是最為熟知并被廣泛研究的中間纖維蛋白家族成員,有著重要的生物學功能,但其具體功能并未被充分了解。有研究顯示,波形蛋白在神經元軸突生長中發揮重要作用[18]。波形蛋白發生翻譯后修飾可以影響其生物學功能[19],在神經軸突生長中發揮重要作用[20],其中糖基化修飾作用是波形蛋白一種重要的翻譯后修飾[21],然而波形蛋白糖基化修飾的相關機制及后續對PC12細胞的生物學功能影響尚未明確。本研究中,使用Cyclo-ManN pro研究波形蛋白糖基化修飾對PC12細胞神經分化的影響。在Uniprot數據庫中對波形蛋白進行生物信息學分析,發現該蛋白有3個潛在糖基化位點,將這3個糖基化位點均進行突變,并構建相應糖基化位點突變質粒,轉染至PC12細胞來進一步研究其對神經分化的影響。結果顯示,波形蛋白糖基化修飾可以延長細胞平均神經突的長度,提高分化細胞的比例,促進PC12細胞的神經突分化與生長。在后續研究中,將設計引物,對波形蛋白的3個糖基化位點分別單獨進行突變,構建相應的糖基化位點7、33、34突變質粒,通過進一步轉染PC12細胞來分析其對神經分化的影響,從而分析具體位點突變對PC12細胞神經分化的作用。
波形蛋白是一種可能的O連接型糖基化蛋白,作用于絲氨酸/蘇氨酸基團,而絲氨酸/蘇氨酸基團同時也是磷酸化位點,可被蛋白激酶A/C磷酸化,這兩種修飾間存在交互作用。在神經細胞中,波形蛋白可被周期蛋白依賴性激酶1磷酸化,并參與整合素β1的激活及后續信號通路,從而在神經軸突生長及再生等方面中發揮作用。因此,波形蛋白糖基化可能通過磷酸化交互作用影響后續細胞神經分化,相關作用機制可能是今后的研究方向。