鮮食葡萄雜交種子萌發(fā)率與親本及親本組合的關(guān)系分析
2024-04-15 04:09:43譚敏李曉梅賀晉瑜劉政海董志剛黃麗萍荀志麗王敏馬小河趙旗峰
山西果樹(shù) 2024年2期
譚敏 李曉梅 賀晉瑜 劉政海 董志剛 黃麗萍 荀志麗 王敏 馬小河 趙旗峰

收稿日期:2023-12-26中文收稿日期
基金項(xiàng)目:現(xiàn)代農(nóng)業(yè)產(chǎn)業(yè)技術(shù)體系國(guó)家葡萄產(chǎn)業(yè)技術(shù)體系(CARS-29-yc-5);山西農(nóng)業(yè)大學(xué)生物育種工程項(xiàng)目(YZGC032)
第一作者簡(jiǎn)介:譚? 敏(1987-),女,碩士,助理研究員,主要從事葡萄栽培生理及遺傳育種研究。E-mail:min.tan@sxau.edu.cn
*通信作者:趙旗峰(1975-),男,研究員,碩士導(dǎo)師,主要從事葡萄遺傳育種及種質(zhì)資源鑒定評(píng)價(jià)研究。E-mail:gssqfzhao@163.com
摘? 要:【目的】常規(guī)雜交育種是葡萄新品種選育的重要方法,通過(guò)對(duì)15年的鮮食葡萄育種資料進(jìn)行整理匯總,分析研究不同倍性親本選擇及不同親本雜交組合配置與雜交種子萌芽率之間的關(guān)系,為鮮食葡萄育種實(shí)踐工作提供參考和借鑒。【方法】以山西農(nóng)業(yè)大學(xué)果樹(shù)研究所2003—2018年間進(jìn)行常規(guī)雜交的45個(gè)鮮食葡萄組合為研究對(duì)象,對(duì)其親本選擇、雜交組合配置和雜交種子萌發(fā)率等進(jìn)行分析。【結(jié)果】1)不同倍性親本選擇及不同親本組合配置,雜交種子萌發(fā)率不同。2)以‘瑰寶作為父本時(shí),雜交種子的萌發(fā)率高。3)45個(gè)雜交組合中,不同年份、相同父母本雜交種子萌發(fā)率不同。4)45個(gè)雜交組合中,玫瑰香和紅地球是良好的母本材料;瑰寶是良好的父本材料;美人指不是理想的親本材料。【結(jié)論】選擇二倍體品種做母本與四倍體品種雜交比用四倍體品種做母本與二倍體品種雜交時(shí)更容易獲得種子;以四倍體為父母本時(shí),很難得到雜交后代。
關(guān)鍵詞:鮮食葡萄;親本組合;雜交種子萌發(fā)率
文章編號(hào):2096-8108(2024)02-0001-06? 中圖分類(lèi)號(hào):S663.1? 文獻(xiàn)標(biāo)識(shí)碼:A
Relationship Analysis of Hybrid Seed Germination Rate of Fresh Grape with Parent and Its Combination
TAN? Min,LI? Xiaomei,HE? Jinyu,LIU? Zhenghai,DONG? Zhigang,HUANG? Liping,XUN? Zhili,WANG? Min,MA? Xiaohe,ZHAO? Qifeng*
(Pomology Institute, Shanxi Agricultural University, Taiyuan Shanxi 030031,China)
Abstract:【Objective】Conventional cross-breeding is an important method for the selection of new grape cultivars. The relationship between different ploidy parents were analyzed by the configuration of different parent hybrid combinations, and the germination rate of hybrid seeds of 15 years on table grape breeding,which provided reference for the practical work of table grape breeding. 【Methods】The 45 combinations of table grape were collected between 2003 to 2018, which grown in the breeding vineyards of Pomology Institute, Shanxi Academy of Agricultural Sciences. The selection of parents, hybrid combination configuration and hybrid seed germination rate were analyzed. 【Results】1)The germination rate of hybrid seeds was different in different ploidy parents selection and combination disposition. 2) The germination rate of hybrid seeds was high when ‘GuiBao was used as the male parent. 3)Among the 45 cross combinations, the seed germination rate of different years and the same parents were different. 4)Among the 45 cross combinations, Muscat Hamburg and Red Globe were good female materials, Guibao was a good paternal materials, and Meirenzhi was not an ideal parental materials. 【Conclusion】It is easier to obtain seeds by crossing diploid cultivars as a female parent than tetraploid cultivars as female parent with diploid cultivars. It is difficult to obtain offspring by crossing tetraploid cultivars as parents.
Keywords:table grape; parent combinations; germination rate of hybrid seeds
葡萄是我國(guó)的大宗果樹(shù)樹(shù)種之一。據(jù)國(guó)家統(tǒng)計(jì)局統(tǒng)計(jì),2021年我國(guó)葡萄栽培產(chǎn)量達(dá)到1 419.54萬(wàn)t,位居柑桔、蘋(píng)果、梨之后排第4位[1]。培育優(yōu)良品種是葡萄產(chǎn)業(yè)健康可持續(xù)發(fā)展的基礎(chǔ)。20世紀(jì)50年代末60年代初至今,我國(guó)已經(jīng)自主育成了402個(gè)葡萄新品種,其中,育種技術(shù)以傳統(tǒng)雜交育種方式為主,共有269個(gè)新品種,育成品種中鮮食葡萄占82.8%[2]。目前,我國(guó)自主選育的一系列名優(yōu)鮮食葡萄品種已應(yīng)用于生產(chǎn)并形成一定規(guī)模,如:香妃、京早晶、早黑寶、巨玫瑰、無(wú)核翠寶、戶(hù)太八號(hào)、醉金香、天工墨玉、澤香、鄭州早紅、紫甜無(wú)核、新雅和新郁等[3-5]。盡管如此,我國(guó)大面積栽培的巨峰、紅地球、夏黑、金手指、紅巴拉多、陽(yáng)光玫瑰等鮮食葡萄品種均來(lái)源于國(guó)外引種[6]。因此,我們有必要加大葡萄育種工作力度,逐步縮短與國(guó)外發(fā)達(dá)地區(qū)育種工作的差距。
截至目前,我國(guó)鮮食葡萄的育種方向主要集中在大粒、抗病、早熟、耐貯運(yùn)、無(wú)核和具有香味等方面[7]。育種方式仍以常規(guī)雜交為主,并且結(jié)合葡萄特定育種目標(biāo)輔以化學(xué)誘變、胚挽救和分子標(biāo)記輔助育種等技術(shù)手段。陳俊等[8]將二倍體瑰寶與二倍體早玫瑰的雜交種子用秋水仙素進(jìn)行誘變處理,通過(guò)連續(xù)多年細(xì)胞學(xué)顯微鏡鏡檢和田間選擇育成了歐亞種四倍體葡萄新品種早黑寶。蔣愛(ài)麗等[9]以二倍體無(wú)核品種喜樂(lè)為母本與四倍體品種巨峰雜交,經(jīng)胚離體挽救培養(yǎng)育成滬培1號(hào)。潘學(xué)軍、紀(jì)薇、王剛和路鳳珍等[10-13]對(duì)葡萄胚挽救苗移栽技術(shù)進(jìn)行了研究,發(fā)現(xiàn)適齡健壯胚挽救苗蘸取適量生根粉并移栽至配比合適的營(yíng)養(yǎng)基質(zhì)中煉苗能提高葡萄胚挽救苗成活率。郭海江等[14]利用葡萄無(wú)核、抗黑痘病和抗炭疽病基因標(biāo)記,對(duì)多個(gè)胚挽救雜交后代進(jìn)行了抗病性輔助選擇。張劍俠等[15]利用無(wú)核分子標(biāo)記對(duì)胚挽救雜交后代進(jìn)行早期鑒定,從中篩選出了無(wú)核雜種。前人關(guān)于鮮食葡萄育種方式和育種技術(shù)手段等作了大量研究,而關(guān)于雜交種子的形成及萌發(fā)等方面的研究比較少。以有性繁殖為主的植物,其種子發(fā)育、成熟以及由種子轉(zhuǎn)化成幼苗的過(guò)程是雜交育種工程中最關(guān)鍵的環(huán)節(jié),直接決定了種群能否實(shí)現(xiàn)更新和壯大[16]。通過(guò)對(duì)15年的鮮食葡萄育種資料進(jìn)行整理匯總,分析研究不同倍性親本選擇及不同親本雜交組合配置與雜交種子萌芽率之間的關(guān)系,為今后進(jìn)一步有效開(kāi)展鮮食葡萄育種實(shí)踐工作提供參考和借鑒,以便提高育種效率,加速育種進(jìn)程。
1? 材料與方法
1.1? 試驗(yàn)材料
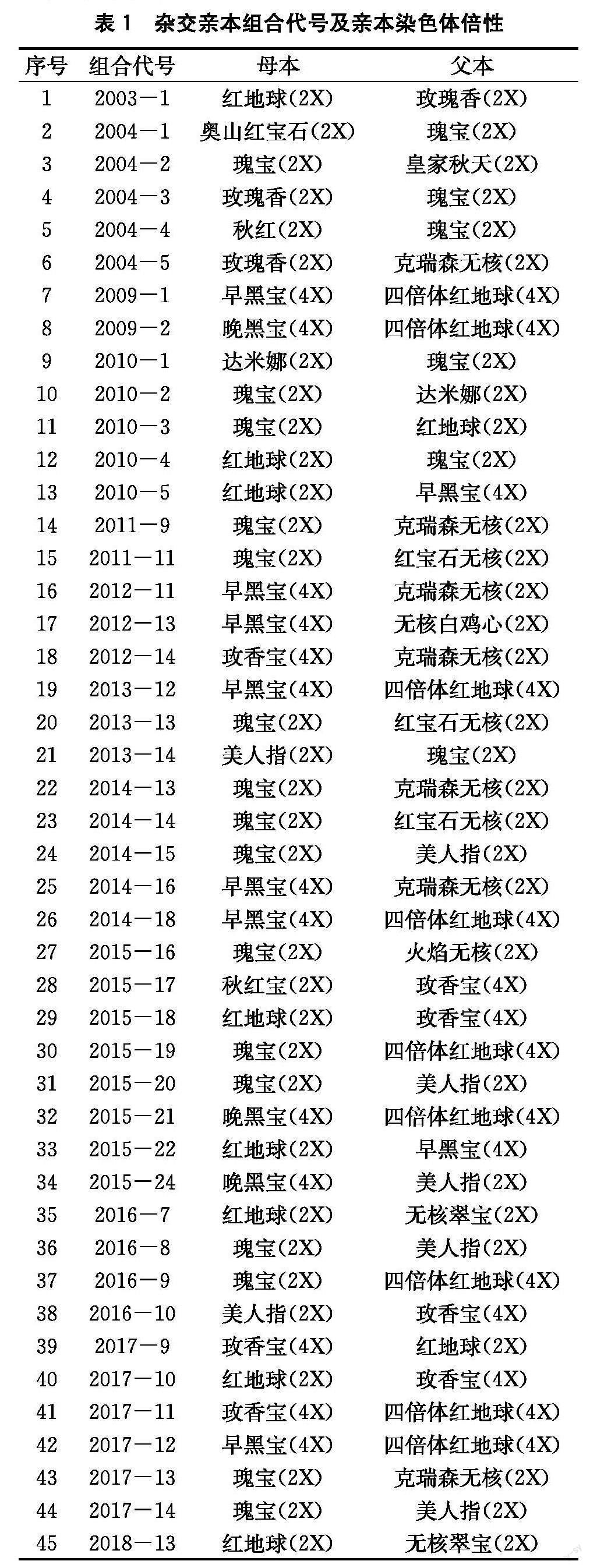
試驗(yàn)材料選自山西農(nóng)業(yè)大學(xué)果樹(shù)研究所葡萄種質(zhì)創(chuàng)制與利用創(chuàng)新團(tuán)隊(duì)育種圃。選取了2003—2018年的45個(gè)鮮食葡萄雜交組合種子萌發(fā)率進(jìn)行研究。其中,雜交親本組合代號(hào)為年份加當(dāng)年雜交組合排列序號(hào)(見(jiàn)表1)。
1.2? 試驗(yàn)方法
1.2.1? 種子層積處理
試驗(yàn)數(shù)據(jù)采集于2003年—2018年,在當(dāng)年10月份于葡萄果實(shí)成熟期采下果實(shí),放于陰涼干燥處貯存后熟;11月份取出浸泡,人工搓掉其果肉、果皮,將種子表面褐膜洗脫后晾干,用清水漂洗數(shù)次,沖洗干凈后于室內(nèi)自然風(fēng)干;12月底進(jìn)行層積處理。
1.2.2? 種子催萌
每年3月下旬取出沙藏層積的種子,清水沖洗干凈后,在溫度25℃、濕度50%的氣候箱中進(jìn)行催芽。催芽期間,每天用自來(lái)水沖洗一次,以防止種子發(fā)霉和保濕,以種子開(kāi)裂露白或長(zhǎng)根為萌芽標(biāo)準(zhǔn)記下不同雜交組合當(dāng)天發(fā)芽的種子數(shù),最終根據(jù)累計(jì)發(fā)芽種子數(shù),計(jì)算種子萌芽率。
1.3? 統(tǒng)計(jì)分析
利用Excel 2019進(jìn)行數(shù)據(jù)分析。
雜交種子萌發(fā)率=發(fā)芽種子粒數(shù)÷種子總粒數(shù)×100%
2? 結(jié)果與分析
2.1? 二倍體與二倍體葡萄雜交其雜交種子萌芽情況
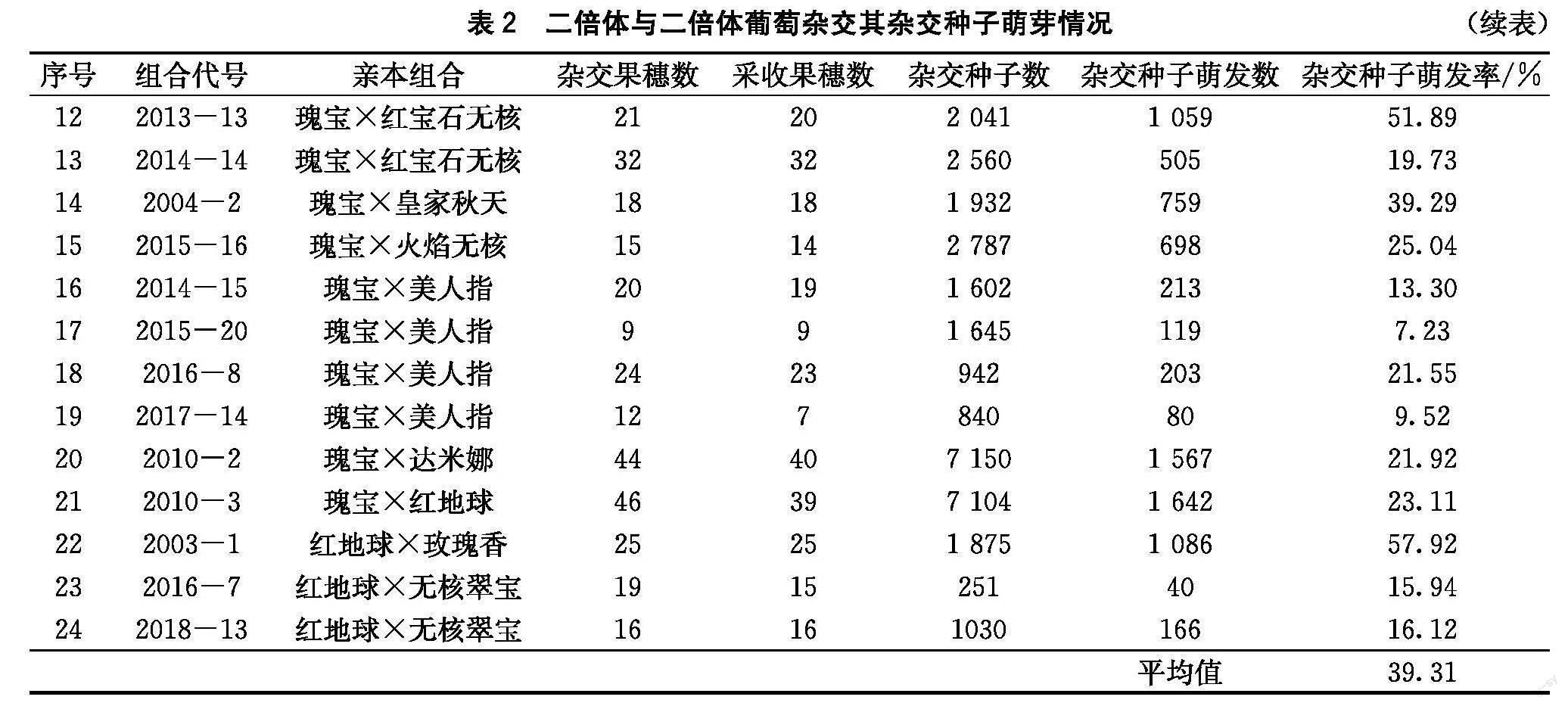
24個(gè)二倍體雜交組合雜交種子萌發(fā)情況如表2所示。不同年份不同親本組合的雜交種子萌發(fā)率為5.89%~91.86%,平均值為39.31%。以瑰寶為父本的6個(gè)雜交組合種子萌發(fā)率為32%~91.86%,平均值為73.07%;其中,美人指×瑰寶(2013-14)雜交種子萌發(fā)率最低為32%。以瑰寶為母本的14個(gè)雜交組合種子萌發(fā)率為5.89%~51.89%,平均值為23.13%,較以瑰寶為父本時(shí)的雜交種子萌發(fā)率低;其中,以無(wú)核葡萄品種為父本時(shí),8個(gè)雜交組合種子萌發(fā)率平均值為28.40%;以有核葡萄品種為父本時(shí),6個(gè)雜交組合種子萌發(fā)率平均值為19.31%,較以無(wú)核葡萄為父本時(shí)種子萌發(fā)率低,其中又以瑰寶×美人指的雜交種子萌發(fā)率分最低,平均值為12.90%。以克瑞森無(wú)核為父本的4個(gè)雜交組合:玫瑰香×克瑞森無(wú)核(2004-5)雜交種子萌發(fā)率高達(dá)91.23%、而以瑰寶為母本,克瑞森無(wú)核為父本的3年雜交種子萌發(fā)率平均值僅為20.40%。以玫瑰香為母本的2個(gè)雜交組合,玫瑰香×瑰寶(2004-3)雜交種子萌發(fā)率為86.20%;玫瑰香×克瑞森無(wú)核(2004-5)雜交種子萌發(fā)率為91.23%。由此說(shuō)明,玫瑰香是理想母本。以紅地球?yàn)槟副镜?個(gè)雜交組合,以紅地球×瑰寶(2010-4)的雜交種子萌發(fā)率65.34%最高,紅地球×玫瑰香(2003-1)的雜交種子萌發(fā)率也達(dá)到了57.92%。而紅地球×無(wú)核翠寶(2016-7、2018-13)的雜交種子萌發(fā)率分別只有15.94%、16.12%,并無(wú)大的差異;綜上,以紅地球?yàn)槟副緯r(shí),無(wú)核翠寶并不是理想父本。
在葡萄育種中,正反交產(chǎn)生的不同細(xì)胞質(zhì)遺傳效應(yīng),也是提高葡萄育種效率的有效方法。比較瑰寶×達(dá)米娜(2010-2)與達(dá)米娜×瑰寶(2010-1),瑰寶×紅地球(2010-3)與紅地球×瑰寶(2010-4),瑰寶×美人指(2014-15、2015-20、2016-8、2017-14)與美人指×瑰寶(2013-14)3組正反交組合發(fā)現(xiàn):3組正反交組合,均以瑰寶作為父本時(shí),雜交種子的萌發(fā)率高。
依據(jù)以上相關(guān)分析結(jié)果,可以發(fā)現(xiàn)瑰寶是良好的父本材料;玫瑰香、紅地球是良好的母本材料;瑰寶×美人指與美人指×瑰寶、紅地球×無(wú)核翠寶都不是理想的雜交組合。另外,不同年份、相同父母本種子萌發(fā)率不同。
2.2? 二倍體與四倍體葡萄雜交其雜交種子萌芽情況
表3、4分別是利用二倍體做母本、四倍體做父本的8個(gè)雜交組合及四倍體做母本、二倍體做父本的6個(gè)雜交組合所獲得的雜交種子萌發(fā)情況。以二倍體葡萄為母本,四倍體葡萄為父本的8個(gè)組合雜交種子萌發(fā)率為1.06%~53.41%,平均值為19.89%(見(jiàn)表3);以四倍體葡萄為母本,二倍體葡萄為父本的6個(gè)組合雜交種子萌發(fā)率為0~9.49%,均未超過(guò)10%,平均值為2.27%(見(jiàn)表4)。以玫香寶為母本的2個(gè)組合:玫香寶×紅地球(2017-9)種子萌發(fā)率為0,玫香寶×克瑞森無(wú)核(2012-14)僅萌發(fā)了1粒種子,種子萌發(fā)率為0.11%(見(jiàn)表4);而以玫香寶為父本的4個(gè)雜交組合種子萌發(fā)率均高于做母本時(shí)的種子萌發(fā)率,平均值為20.80%,說(shuō)明玫香寶適宜做父本(見(jiàn)表3)。以紅地球?yàn)槟副镜?個(gè)組合雜交種子萌發(fā)率均超過(guò)10%,平均值為27.40%(見(jiàn)表3);而以紅地球做父本時(shí),玫香寶×紅地球(2017-9)種子萌發(fā)率為0(見(jiàn)表4),說(shuō)明紅地球適宜做母本。
瑰寶×四倍體紅地球連續(xù)2年雜交的種子萌發(fā)率2015-19為3.12%、2016-9為6.90%,均未超過(guò)10%,說(shuō)明這兩個(gè)組合并不是理想的雜交組合(見(jiàn)表3)。美人指×玫香寶(2016-10)、晚黑寶×美人指(2015-24)這兩個(gè)組合的雜交種子萌發(fā)率分別為1.06%、1.43%,說(shuō)明美人指亦不是二倍體和四倍體雜交組合理想的親本選擇(見(jiàn)表3)。以早黑寶為母本、二倍體無(wú)核葡萄品種做父本的2個(gè)雜交組合中,早黑寶×無(wú)核白雞心(2012-13)雜交種子萌發(fā)率為9.49%;早黑寶×克瑞森無(wú)核(2012-11)雜交種子萌發(fā)率為2.60%;這2個(gè)雜交組合可做進(jìn)一步研究(見(jiàn)表4)。
依據(jù)以上相關(guān)分析結(jié)果,可知以二倍體葡萄為母本,四倍體葡萄為父本時(shí)易獲得雜交種子。紅地球是良好的母本材料;玫香寶是良好的父本材料;美人指不是理想的親本選擇材料。在表3、4中同樣,可以發(fā)現(xiàn),不同年份、相同父母本種子萌發(fā)率不同。
2.3? 四倍體與四倍體葡萄進(jìn)行雜交其雜交種子萌芽情況
表5是以四倍體玫瑰香型葡萄品種早黑寶(歐亞種早熟四倍體)、玫香寶(歐美雜交種早熟四倍體)、晚黑寶(歐亞種晚熟四倍體)為母本,以四倍體紅地球(歐亞種晚熟四倍體)為父本的7個(gè)四倍體雜交組合種子萌發(fā)情況。7個(gè)組合雜交種子萌發(fā)率均極低,為0~2.63%,平均值僅為0.78%。同時(shí)從表5中可以看到,在不同年份相同父母本種子萌發(fā)率亦不同。
3? 討論與結(jié)論
本文通過(guò)研究45個(gè)鮮食雜交組合,4種倍性雜交組合(二倍體×二倍體、二倍體×四倍體、四倍體×二倍體、四倍體×四倍體)的雜交種子萌發(fā)率,對(duì)親本選擇、雜交組合配置與雜交種子萌發(fā)率的關(guān)系進(jìn)行分析。結(jié)果表明,葡萄不同倍性品種間雜交,雜交后代種子獲得及雜交種子萌發(fā)率難易程度不同。以二倍體葡萄為父母本的24個(gè)雜交組合雜交種子萌發(fā)率為5.89%~91.86%,平均值最高,為39.31%;以二倍體葡萄為母本,四倍體葡萄為父本的8個(gè)雜交組合雜交種子萌發(fā)率為1.06%~53.41%,平均值為19.89%;以四倍體葡萄為母本,二倍體葡萄為父本的6個(gè)雜交組合雜交種子萌發(fā)率為0~9.49%,平均值為2.27%;以四倍體為父母本的7個(gè)雜交組合雜交種子萌發(fā)率極低,為0~2.63%,平均值為0.78%。從以上研究結(jié)果可以看出,一是選擇二倍體品種做母本與四倍體品種雜交比用四倍體品種做母本與二倍體品種雜交時(shí)更容易獲得種子,這與徐海英、郭印山等[17-18]研究結(jié)果相同。二是以四倍體為父母本時(shí),很難得到雜交后代。這可能一方面與四倍體本身存在胚 、胚乳敗育和胚珠退化現(xiàn)象,得不到成熟種子有關(guān);另一方面可能與四倍體種子相對(duì)較大,種皮厚且堅(jiān)硬,水分不容易進(jìn)入,不易萌發(fā)有關(guān)[19]。關(guān)于不同倍性親本間雜交親和性問(wèn)題,根據(jù)Endosperm Balance Number(EBN)假說(shuō),在雜種的胚乳中,只有當(dāng)父本與母本的基因組成比例為2∶1時(shí),胚乳才能正常發(fā)育,形成健全的種子,否則父母本染色體組成比例發(fā)生紊亂,胚乳發(fā)育中途停滯,造成胚的敗育[20]。由此看來(lái),多倍體常規(guī)雜交育種方法的最大障礙是雜交親本親和力差、坐果率低、雜種胚早期敗育或胚乳解體,獲得的雜交種子生活力低,以至于很難得到雜交后代。在今后工作中,我們將就如何合理選擇親本、配置雜交組合和運(yùn)用胚培養(yǎng)技術(shù)等育種工作繼續(xù)進(jìn)行研究。
品種資源是葡萄新品種選育的關(guān)鍵,豐富的資源可以提高培育新品種的機(jī)率,降低成本。本文中,利用頻次最高的雜交親本是瑰寶,其主要被作為父本材料。瑰寶[21]是我所自主選育的晚熟優(yōu)良鮮食品種,肉質(zhì)甜脆并具濃郁的玫瑰香味,品質(zhì)極佳。目前,我們以瑰寶為親本,已自主選育了包括3個(gè)四倍體葡萄新品種(早黑寶、秋黑寶、晚黑寶)[8,22-23],6個(gè)二倍體葡萄新品種(秋紅寶、早康寶、麗紅寶、無(wú)核翠寶、晶紅寶、翠香寶)等在內(nèi)的9個(gè)鮮食葡萄新品種[24-29]。因此,如何進(jìn)一步發(fā)掘我國(guó)自育品種資源的價(jià)值,提高育種材料利用率,培育出更多的優(yōu)良品種也是育種工作的重要任務(wù)。同時(shí),在以后的雜交工作中應(yīng)該擴(kuò)大親本選擇范圍,集合更多的優(yōu)異葡萄種質(zhì)資源進(jìn)行雜交組合配置,從而豐富鮮食葡萄種質(zhì)資源。
本文中相同父母本不同年份的雜交組合雜交種子形成與萌發(fā)率不同,相同父母本正反交雜交種子形成與萌發(fā)率也不相同,這與前人研究結(jié)果一致[30]。這說(shuō)明雜交種子的形成與萌發(fā)率受到了親本倍性、正反交、外界環(huán)境條件等的影響,所以雜交親本的選擇與種子層積處理、萌發(fā)溫濕度環(huán)境因子控制等同樣至關(guān)重要。
中文致謝
參考文獻(xiàn)
[1]? 中華人民共和國(guó)國(guó)家統(tǒng)計(jì)局年度數(shù)據(jù)[EB]: https://data.stats.gov.cn/adv.htm?m=advquery&cn=C01., 2021. 10.13414/j.cnki.zwpp.2022.01.003.
[2]? 諸葛雅賢,徐衛(wèi)東,李紹星,等. 中國(guó)葡萄育種單位及其育種情況的分析[J]. 落葉果樹(shù), 2023, 55(1): 31-35.
[3]? 姜建福,孫海生,劉崇懷,等. 2000年以來(lái)中國(guó)葡萄育種研究進(jìn)展[J]. 中外葡萄與葡萄酒, 2010(3): 60-65, 69.
[4]? 王勇,李玉玲,孫鋒,等. 2010年以來(lái)中國(guó)葡萄育種研究進(jìn)展[J]. 中外葡萄與葡萄酒, 2021(6): 90-97.
[5]? 孫鋒,駱強(qiáng)偉,廖康,等. 新疆葡萄育種方向與研究進(jìn)展[J]. 農(nóng)業(yè)科技通訊, 2014(12): 141-142, 145.
[6]? 楊亞蒙,姜建福,樊秀彩,等. 2000年以來(lái)我國(guó)葡萄國(guó)外引種概況[J]. 中外葡萄與葡萄酒, 2018(2): 54-59.
[7]? 秦子禹,李上云,王晨,等. 河北省葡萄育種成果及現(xiàn)狀分析[J]. 中外葡萄與葡萄酒, 2019(5): 81-84.
[8]? 陳俊. 早熟大粒優(yōu)質(zhì)葡萄新品種——早黑寶[J]. 柑桔與亞熱帶果樹(shù)信息, 2001(9): 18.
[9]? 蔣愛(ài)麗,李世誠(chéng),金佩芳,等. 胚培無(wú)核葡萄新品種——滬培1號(hào)的選育[J]. 果樹(shù)學(xué)報(bào), 2007(3): 402-403, 256.
[10]? 潘學(xué)軍,王躍進(jìn),張劍俠,等. 葡萄胚挽救苗移栽技術(shù)的研究[J]. 西北植物學(xué)報(bào), 2004(6): 1077-1082.
[11]? 紀(jì)薇,高美英,張鵬飛,等. 無(wú)核葡萄胚挽救苗的馴化及移栽技術(shù)[J]. 山西農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版), 2015, 35(2): 147-150.
[12]? 王剛,李桂榮,王躍進(jìn),等. 無(wú)核葡萄胚挽救試管苗馴化移栽體系優(yōu)化[J]. 西北農(nóng)林科技大學(xué)學(xué)報(bào)(自然科學(xué)版), 2015, 43(8): 153-158.
[13]? 路鳳珍,溫曉敏,張娜,等. 三倍體葡萄胚挽救苗馴化及移栽技術(shù)研究[J]. 天津農(nóng)業(yè)科學(xué), 2016, 22(10): 92-95.
[14]? 郭海江,王躍進(jìn),張劍俠,等. 葡萄抗病無(wú)核胚挽救育種及分子標(biāo)記輔助選擇[J]. 西北植物學(xué)報(bào), 2005(12): 2395-2401.
[15]? 張劍俠. 中國(guó)野生葡萄抗病基因標(biāo)記及輔助育種應(yīng)用研究[D]. 西北農(nóng)林科技大學(xué), 2006.
[16]? 譚廷鴻,王卓,吳禮佳,等. 瀕危植物梵凈山冷杉的球果與種子性狀研究[J]. 西北植物學(xué)報(bào), 2022, 42(10): 1671-1681.
[17]? 徐海英,閻愛(ài)玲,張國(guó)軍. 葡萄二倍體與四倍體品種間雜交胚珠的離體培養(yǎng)[J]. 果樹(shù)學(xué)報(bào), 2001(6): 317-320.
[18]? 郭印山,郭修武,張海娥,等. 利用胚挽救技術(shù)獲得三倍體葡萄植株研究[J]. 沈陽(yáng)農(nóng)業(yè)大學(xué)學(xué)報(bào), 2005(5): 606-608.
[19]? 檀英霞. 四倍體葡萄及其雜交后代有性過(guò)程的研究[D]. 河北農(nóng)業(yè)大學(xué), 2001.
[20]? 孫興民. 葡萄三倍體新株系的獲得[D]. 南京農(nóng)業(yè)大學(xué), 2011.
[21]? 陳俊. 瑰寶葡萄優(yōu)質(zhì)豐產(chǎn)栽培技術(shù)[J]. 山西果樹(shù), 1994(1): 15-16.
[22]? 馬小河,唐曉萍,陳俊,等. 優(yōu)質(zhì)中熟葡萄新品種‘秋黑寶[J]. 園藝學(xué)報(bào), 2010, 37(11): 1875-1876.
[23]? 董志剛,李曉梅,譚偉,等. 晚熟葡萄新品種晚黑寶優(yōu)質(zhì)豐產(chǎn)栽培技術(shù)[J]. 中國(guó)果樹(shù), 2015(6): 71-73, 86.
[24]? 陳俊. 早熟無(wú)核葡萄新品種——早康寶的選育[J]. 中國(guó)果業(yè)信息, 2009, 26(12): 53.
[25]? 唐曉萍,陳俊,馬小河,等. 鮮食無(wú)核葡萄新品種——‘晶紅寶的選育[J]. 果樹(shù)學(xué)報(bào), 2014, 31(1): 159-160.
[26]? 陳俊,唐曉萍,馬小河,等. 優(yōu)質(zhì)中熟無(wú)核葡萄新品種‘麗紅寶[J]. 園藝學(xué)報(bào), 2011, 38(3): 595-596.
[27]? 唐曉萍,陳俊,馬小河,等. 早熟無(wú)核葡萄新品種‘無(wú)核翠寶[J]. 園藝學(xué)報(bào), 2012, 39(11): 2307-2308.
[28]? 唐曉萍,馬小河,董志剛,等. 晚熟鮮食葡萄新品種‘翠香寶[J]. 園藝學(xué)報(bào), 2020, 47(S2): 2900-2901.
[29]? 陳俊,唐曉萍,馬小河,等. 中晚熟葡萄新品種——秋紅寶的選育[J]. 果樹(shù)學(xué)報(bào), 2007(6): 867-868, 728.
[30]? 李莎莎. 葡萄種子敗育因子與無(wú)核抗病葡萄種質(zhì)創(chuàng)制研究[D]. 西北農(nóng)林科技大學(xué), 2021.
