狗頭棗ACO Ⅰ基因RNAi表達載體的構建及遺傳轉化
2024-01-08 06:33:50尚秉眾田瑞康雒淑婷王真珍陳國梁
延安大學學報(自然科學版) 2023年4期
尚秉眾,田瑞康,雒淑婷,王真珍,陳國梁,2,3*
(1.延安大學 生命科學學院;2.陜西省紅棗重點實驗室;3.陜西省黃土高原菌產業生態循環發展工程技術研究中心,陜西 延安 716000)
棗樹(ZizyphusjujubeMill)是原產于我國的最具特色和優勢的果樹之一。近年來國內外市場對鮮棗的需求量日漸增加[1],但是鮮棗采摘后易失水、皺縮,果肉酒軟、褐變、腐爛,保鮮期極短,大大縮短了鮮棗市場的貨架期[2]。乙烯作為一種氣態的植物激素,在生理、生化和分子水平上參與果實成熟軟化[3-4]。乙烯生物合成途徑中有兩個關鍵反應:在第一個反應中,S-腺苷-L-甲硫氨酸(SAM)被ACC-合酶(ACS)轉化為1-氨基環丙烷-1-羧酸(ACC);在第二個反應中,ACC 被ACC 氧化酶(ACO)轉化為乙烯。其中ACO 是乙烯生物合成中的關鍵限速酶[5]。研究表明,ACO 基因為多基因家族編碼的,在番茄中擁有9 個ACS 和5 個ACO 基因,其中在果實成熟過程中參與乙烯代謝的ACO Ⅰ為主要表達基因[6-8],故采用RNAi 技術對該基因進行特異性沉默來提高其果實的耐儲藏性成為可能,而有關以ACO 基因為靶標利用RNAi 技術改變其耐貯性的報道較多。吳曉慶等[9]通過正義,反義ACO 基因轉化石竹得到了花期較長的石竹材料;胡春華等[10]通過反義技術抑制香蕉ACO基因,得到了成熟期相比于對照推遲了十天的轉基因香蕉;張巖等[11]也通過RNAi 技術對雙孢蘑菇的ACO 基因進行干擾,有效的降低了其表達量,并使ACO 酶活力降低至68%以上。目前,RNAi 技術已經應用于番茄、珠子參、柑橘、草莓、馬鈴薯等[12-16]研究中。但將RNAi 技術應用在棗屬植物研究相關基因功能還鮮有報道。基于此,本文選取狗頭棗ACO Ⅰ基因保守序列作為靶標構建RNAi表達載體[17],以木棗葉片為外植體對其進行遺傳轉化,以期獲得轉基因材料,為培育出耐貯藏的轉基因紅棗奠定基礎。
1 材料與方法
1.1 材料與試劑
ZK品系木棗試管苗,由陜西省紅棗重點實驗室提供。大腸桿菌DH5α、農桿菌LBA4404、質粒pGM-TR(含基因片段ACO Ⅰ3-4、pART27 與pKANNⅠBAL 均為本課題組保存。實驗所需的各種酶均購自TaKaRa公司。
1.2 引物與酶切位點設計
引物與酶切位點組成:
F:5′-GCGGATCCBamHⅠCTCGAGXhoⅠATGAAATT GGAGACACTAGCTGAGG-3′
R:5′-GCATCGATClaⅠGAATTCEcoRⅠAACATTAGT TTCCACTGCTTTC-3′
1.3 RNAi表達載體pART ACO I3-4的構建
采用XhoⅠ/EcoR Ⅰ雙酶切pGM-TR(含ACO Ⅰ3-4)及pKANNⅠBAL載體,分別回收靶標片段及線性載體并進行連接,獲得pKF1 載體(含正向片段);再將pGM-TR 和pKF1 載體用BamH Ⅰ/ClaⅠ雙酶切,分別回收靶標片段及線性載體pKF1 并進行連接,得到中間載體pKF(含正反向片段)。用NotⅠ分別酶切pKF 與pART 27 質粒載體,回收4 200 bp 目的片段并與pART 27線性載體連接,得到了pART ACOⅠ3-4植物表達載體(圖1)。

圖1 pART ACO Ⅰ3-4載體構建過程
1.4 木棗葉片分化及繼代培養Kan選擇壓篩選
設置了含有5、10、15、20、25、30 mg·L-1Kan 濃度梯度的葉片分化培養基及繼代培養基進行Kan選擇壓的篩選實驗。選取生長健壯生長20 d 左右的木棗組培苗將其葉片剪成大小約為0.5 cm2的小塊、將莖芽切段分別接種在相應培養基上,觀察葉片愈傷組織形成及莖芽繼代生長情況并記錄,進行Kan選擇壓的確定。
1.5 木棗遺傳轉化及篩選鑒定
將pART ACOⅠ3-4表達載體質粒導入農桿菌LBA4404 中獲得工程農桿菌,對ZK 品系木棗苗葉片進行轉化。在MS固體培養基礎上每升添加6-BA 1.0 mg,IBA 0.2 mg,TDZ 0.01 mg 作為基礎培養基,在此基礎上每升添加AgNO30.5 mg 作為共培養基,在共培養基中添加適量Carb+Kan 進行選擇抑菌培養基。將抗性篩選獲得的陽性再生木棗植株的葉片作為材料,利用DNA 提取試劑盒提取植物DNA組并進行PCR與電泳鑒定。
2 結果分析
2.1 pART ACO Ⅰ3-4表達載體的成功構建
獲得的pKF1 經XhoⅠ/EcoR Ⅰ酶切及PCR 鑒定均獲得大小約為600 bp 的片段(圖2)與預期大小一致,表明成功獲得pKF1。將pKF 用NotⅠ酶切,經電泳檢測得到大小約為4 200 bp 和3 100 bp 的片段(圖3)與預期結果一致,表明中間載體構建成功。將得到的pART ACO Ⅰ3-4分別用BamH Ⅰ/ClaⅠ雙酶切及EcoR Ⅰ單酶切,電泳共檢出大小約為1 300 bp,600 bp 和750 bp 的三條帶(圖4),與預期結果一致;再用Hind Ⅲ、XhoⅠ分別酶切pART ACO Ⅰ3-4,電泳出現約1 000 bp、2 000 bp 大小的片段(圖5)。對pART ACO Ⅰ3-4進行PCR 擴增鑒定,結果出現了一條大小約為600 bp 的條帶(圖6),說明pART ACOⅠ3-4表達載體構建成功。

圖2 pKF1酶切與PCR鑒定

圖3 pKF酶切圖
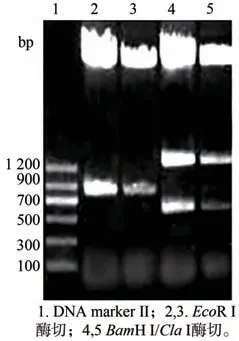
圖4 pART ACO Ⅰ3-4的酶切鑒定

圖5 pART ACO Ⅰ3-4的酶切鑒定

圖6 pART ACO Ⅰ3-4的電泳鑒定
2.2 Kan選擇壓的確定
木棗組培苗在10 mg·L-1Kan 的繼代培養基上能夠正常生長,隨著繼代培養基中Kan濃度的升高,木棗組培苗的葉子逐漸變黃,開始出現白化現象,生長態勢受阻。直到繼代培養中Kan濃度升為30 mg·L-1時,木棗組培苗葉子全部變黃、白化,并且死亡。故將30 mg·L-1Kan 濃度設為木棗遺傳轉化再生苗繼代選擇培養基的濃度上限。在Kan 濃度為5 mg·L-1時,葉片出愈率為98.3%,愈傷組織不定芽分化率為61.7%;當Kan 濃度達到10 mg·L-1時,葉片出愈率為37.7%,愈傷組織不定芽分化率11.7%,當Kan 濃度達到15 mg·L-1時,葉片出愈率為6.7%,愈傷組織不定芽1.7%。當Kan濃度達到20 mg·L-1時,愈傷組織趨于死亡,無不定芽分化(圖7),所以可將20 mg·L-1Kan濃度設為木棗葉片遺傳轉化愈傷組織及不定芽形成的Kan標記選擇壓濃度。
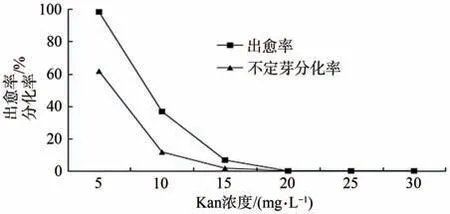
圖7 Kan選擇壓的確定
2.3 轉化植株的篩選與PCR鑒定
共侵染木棗葉塊600片,共培養之后,放入暗箱進行暗培養3 d,之后移到光下進行培養。4 周后分化出愈傷組織及并逐漸分化成苗(圖8A);而對照組葉片沒有愈傷組織形成及分化并且開始發黃(圖8B)。以分化成苗植株葉片DNA 為模板(圖9),經過PCR 與電泳檢測鑒定,陰性對照只能擴增出一個條帶,大小約為750 bp,然而轉化再生植株除可以擴增出兩條條帶,除了陰性對照組的750 bp 條帶外,還能擴增出一條大小約為600 bp 的特異性目的條帶(圖10),初步證明目的基因轉化成功。

圖8 木棗愈傷組織及不定芽的形成在分化培養基上

圖9 陽性苗在30 mg·L-1 Kan選擇壓下的生長情況

圖10 陽性植株的PCR檢測
3 討論與結論
3.1 討論
鮮棗采收后不易保鮮、不耐貯藏,其果實保鮮期的長短是影響鮮果銷售價值的關鍵品質,是影響其經濟價值的重要因素之一,嚴重制約著紅棗產業的發展[18]。目前采用RNAi 技術賦予植物抗性和調控植物代謝等領域已研發出多種具有優良性狀的作物,如已成功培育出耐貯藏轉基因番茄、桃子、哈密瓜等[19-21],表明通過調控ACO 基因的表達可達到調控乙烯的生成,緩解果實成熟衰老速度,提高果實的耐貯性。人們在棗基因組數據庫中發現有ZjACO1、ZjACO2和ZjACO3等3 個ACO 同源基因且其相對表達量與果實成熟度呈顯著正相關,其中ZjACO1在棗果實七個不同發育階段的轉錄表達模式具有極顯著差異[22],因此ZjACO1與果實成熟度相關性最高,而我們沉默的靶標基因正是ZjACO1。
棗品種繁多,呼吸類型復雜,薛夢林等[23]認為棗為非呼吸躍變型果實,由于非躍變型果實成熟衰老過程產生的乙烯遠低于躍變型果實,乙烯被認為在非躍變型果實成熟衰老中發揮的作用非常有限。然而,在對柑橘果實的研究表明,低水平的內源乙烯調控了果實的成熟衰老。近年來,隨著乙烯參與非躍變型果實成熟衰老相關研究陸續報道,研究人員發現,非躍變型果實諸多成熟相關過程也直接受乙烯調控,且非躍變型果實和躍變型果實品質變化的某些分子調控途徑非常相似[24]。越來越多的研究表明,乙烯在非躍變型果實成熟衰老的調控中扮演著重要的角色。如在香蕉、荔枝、柑橘和番茄等[25-28]果實的延遲成熟、延長保質期等方面取得成功。雖然ACO 是催化乙烯生物合成途徑的最后關鍵步驟,并作為靶向基因構建了內含子嵌入的倒置重復序列載體對其進行調控,但干擾效果如何及該基因對棗的成熟和儲藏影響如何,還有待于進一步驗證。
3.2 結論
成功構建了ihRNAi 植物表達載體,初步篩選到木棗葉片愈傷組織分化培養基,其配方為MS+6-BA 1.0 mg·L-1+IBA 0.2 mg·L-1+TDZ 0.01 mg·L-1,添 加0.5 mg·L-1Carb+20 mg·L-1Kan 可作為篩選培養基。確定了木棗葉片愈傷組織形成Kan 篩選濃度為20 mg·L-1,再生苗的Kan 篩選濃度為30 mg·L-1。采用葉盤法對木棗進行轉化,經抗生素篩選和PCR 檢測,初步得到了16株陽性木棗轉基因植株。
