大腸桿菌胞內(nèi)L-丙氨酸響應(yīng)型啟動(dòng)子的篩選與表征
2023-11-06 09:05:36聶玉朋徐建中劉立明姜國政王深鏢田延軍
中國釀造 2023年10期
聶玉朋,徐建中,劉立明,劉 佳,徐 慧,姜國政,楊 奕,王深鏢,田延軍*
(1.齊魯工業(yè)大學(xué)(山東省科學(xué)院)山東省食品發(fā)酵工業(yè)研究設(shè)計(jì)院,山東 濟(jì)南 250014;2.江南大學(xué) 生物工程學(xué)院工業(yè)生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室,江蘇 無錫 214122;3.江南大學(xué) 食品科學(xué)與技術(shù)國家重點(diǎn)實(shí)驗(yàn)室,江蘇 無錫 214122;4.煙臺(tái)恒源生物股份有限公司,山東 煙臺(tái) 265709)
L-丙氨酸(L-alanine,L-Ala),又稱L-α-氨基丙酸,是人體血液中含量最高的中性氨基酸,被廣泛應(yīng)用于醫(yī)藥、食品和日化領(lǐng)域[1-2]。L-丙氨酸是美國食品和藥品管理局批準(zhǔn)的食品添加劑和營養(yǎng)補(bǔ)充劑[3],用于食品中可以起到增甜提鮮的作用[4];作為一種藥物成分,L-丙氨酸是維生素B6、索非布韋、醋酸格拉替雷等眾多藥品的生產(chǎn)原料,也可用于氨基酸輸液[5-6];此外,L-丙氨酸還被廣泛用作頭發(fā)和皮膚的調(diào)理劑,也被用作化妝品和一些個(gè)人護(hù)理產(chǎn)品的成分[6]。以L-丙氨酸為原料生產(chǎn)新型螯合劑甲基甘氨酸二乙酸(methylglycine-N,N-diacetic acid,MGDA),可替代洗滌劑中的含磷螯合劑,其在水中可以自然降解,避免了對(duì)環(huán)境產(chǎn)生的負(fù)面影響[7-8]。
微生物發(fā)酵法生產(chǎn)成本低、反應(yīng)條件溫和,已成為L-丙氨酸生產(chǎn)的主流方式[9-10]。目前用于L-丙氨酸發(fā)酵生產(chǎn)的主要菌株是大腸桿菌(Escherichia coli)[11-12],其次是谷氨酸棒狀桿菌(Corynebacterium glutamicum)[13-14]。相比于其他的微生物,大腸桿菌具有遺傳背景清晰、基因改造技術(shù)成熟、繁殖速度快和發(fā)酵周期短等優(yōu)點(diǎn)[15-16]。目前,高產(chǎn)L-丙氨酸大腸桿菌細(xì)胞工廠的構(gòu)建主要是通過在大腸桿菌中引入異源的丙氨酸脫氫酶(alanine dehydrogenase,ADH),丙氨酸脫氫酶可高效催化丙酮酸、氨(NH4+)和煙酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NADH)生成L-丙氨酸[17-18]。有研究發(fā)現(xiàn)[19],發(fā)酵液中L-丙氨酸的積累會(huì)降低大腸桿菌細(xì)胞的比生長速率,且異源丙氨酸脫氫酶的高效表達(dá)會(huì)對(duì)菌體造成代謝負(fù)擔(dān)。ZHOU L等[20]設(shè)計(jì)了一種溫度調(diào)節(jié)的遺傳開關(guān)來動(dòng)態(tài)控制丙氨酸脫氫酶的表達(dá),將發(fā)酵分為細(xì)胞生長和L-丙氨酸生產(chǎn)兩個(gè)階段,實(shí)現(xiàn)了發(fā)酵過程的動(dòng)態(tài)調(diào)控,發(fā)酵40 h,L-丙氨酸產(chǎn)量達(dá)120.8 g/L。目前,已有研究報(bào)道了響應(yīng)L-纈氨酸[21]和L-異亮氨酸[22]等氨基酸的生物傳感器,但尚未發(fā)現(xiàn)響應(yīng)L-丙氨酸的生物傳感器。
利用轉(zhuǎn)錄組測(cè)序技術(shù)篩選響應(yīng)天然產(chǎn)物的啟動(dòng)子已經(jīng)是一種比較成熟的啟動(dòng)子建庫策略[23-24]。LIANG G J等[25]在光滑假絲酵母中利用轉(zhuǎn)錄組分析篩選得到了響應(yīng)L-蘋果酸的Pcgr-10啟動(dòng)子元件,并以此構(gòu)建了響應(yīng)L-蘋果酸的動(dòng)態(tài)調(diào)控系統(tǒng),提高了L-蘋果酸產(chǎn)量;汪舒穎等[26]在單增李斯特氏菌(Listeria monocytogenes)中利用轉(zhuǎn)錄組分析篩選到了在厭氧條件下高效表達(dá)的Pan4啟動(dòng)子,用于調(diào)控天青蛋白和θ毒素表達(dá)。本研究對(duì)90 g/LL-丙氨酸壓力下培養(yǎng)的大腸桿菌(Escherichia coli)進(jìn)行比較轉(zhuǎn)錄組測(cè)序分析,篩選在L-丙氨酸壓力下顯著上調(diào)基因的啟動(dòng)子,利用mKate熒光蛋白進(jìn)行表征,添加10 g/L的L-丙氨酸進(jìn)行誘導(dǎo),從中篩選出響應(yīng)L-丙氨酸的啟動(dòng)子,并通過測(cè)定不同濃度L-丙氨酸和不同種類氨基酸誘導(dǎo)下啟動(dòng)子的相對(duì)熒光強(qiáng)度,評(píng)估啟動(dòng)子的靈敏性與特異性。本研究旨在從大腸桿菌中篩選出特異性響應(yīng)L-丙氨酸的啟動(dòng)子元件,為后續(xù)L-丙氨酸動(dòng)態(tài)響應(yīng)系統(tǒng)的構(gòu)建奠定基礎(chǔ)。
1 材料與方法
1.1 材料與試劑
1.1.1 菌株、質(zhì)粒和引物
大腸桿菌(E.coli)W3110、E.coliJM109、質(zhì)粒pTET:由本實(shí)驗(yàn)室保藏。本研究所用引物如表1所示。

表1 本研究所用引物Table 1 Primers in the study
1.1.2 化學(xué)試劑
限制性內(nèi)切酶、脫氧核糖核酸(deoxyribonucleic acid,DNA)Marker、Prime Star高保真酶、T4 DNA連接酶和TaqDNA聚合酶:大連寶生物工程公司;ClonExpress II One step cloning kit:南京諾唯贊生物科技有限公司;細(xì)菌基因組提取試劑盒、質(zhì)粒提取試劑盒、膠回收試劑盒、產(chǎn)物純化試劑盒、氯霉素(分析純):上海生工生物工程技術(shù)有限公司;L-丙氨酸(分析純):上海阿拉丁生化科技股份有限公司;酵母粉、蛋白胨(均為生化試劑):英國Oxoid公司;其他試劑來自國藥集團(tuán)化學(xué)試劑有限公司。
1.1.3 培養(yǎng)基
LB培養(yǎng)基:酵母粉5.0g/L,蛋白胨10.0g/L,NaCl10.0g/L,pH自然,121 ℃條件下高壓滅菌20 min。
LB固體培養(yǎng)基:酵母粉5.0 g/L,蛋白胨10.0 g/L,NaCl 10.0 g/L,瓊脂粉20 g/L,pH自然,121 ℃條件下高壓滅菌20 min。
氯霉素抗性平板:酵母粉5.0 g/L,蛋白胨10.0 g/L,NaCl 10.0 g/L,瓊脂粉20 g/L,pH自然,121 ℃條件高壓滅菌20 min后,添加終質(zhì)量濃度為30 μg/mL的氯霉素。
1.2 儀器與設(shè)備
CFX96聚合酶鏈?zhǔn)椒磻?yīng)(polymerasechainreaction,PCR)儀、Gel Doc凝膠成像儀:美國Bio-Rad公司;BEP-600核酸電泳儀:北京伯蘭特生物科技有限公司;ZQZY-AF8恒溫調(diào)速搖床:上海知楚儀器有限公司;5804R冷凍離心機(jī):德國Eppendorf公司;SpectraMaxRM3多功能酶標(biāo)儀酶標(biāo)儀:美國Molecular Devices公司。
1.3 方法
1.3.1 質(zhì)粒和基因組的提取
細(xì)菌質(zhì)粒DNA提取方法參照上海生工生物工程技術(shù)有限公司細(xì)菌質(zhì)粒提取試劑盒相關(guān)操作說明,基因組提取參照上海生工生物工程技術(shù)有限公司細(xì)菌基因組提取試劑盒相關(guān)操作說明。
1.3.2 轉(zhuǎn)錄組測(cè)序
將培養(yǎng)至對(duì)數(shù)期的大腸桿菌分別在非脅迫(未添加L-丙氨酸的LB培養(yǎng)基)和脅迫(添加90 g/LL-丙氨酸的LB培養(yǎng)基)條件下接種,初始OD600nm值為0.1[27]。培養(yǎng)6 h后,4 ℃、12 000 r/min離心10 min收集細(xì)胞,然后用磷酸鹽緩沖液(phosphate buffer solution,PBS)洗滌兩次,在液氮中快速冷凍,并送到蘇州金唯智生物科技有限公司進(jìn)行轉(zhuǎn)錄組分析。
1.3.3 重組質(zhì)粒構(gòu)建
以pTET質(zhì)粒為模板,pTET-mKate-1/pTET-mKate-2為引物,PCR擴(kuò)增線性載體,PCR擴(kuò)增體系:模板DNA 2 μL,Prime Star高保真酶50 μL,引物1、引物2各2 μL,雙蒸水(ddH2O)44 μL。PCR擴(kuò)增程序:94 ℃預(yù)變性10 min,循環(huán)程序(94 ℃變性10 s,55 ℃退火30 s,72 ℃延伸1 000 bp/min),循環(huán)程序運(yùn)行29次,后延伸72 ℃,5 min。將PCR擴(kuò)增后的產(chǎn)物膠回收后得到線性化載體片段。以E.coliW3110基因組為模板,擴(kuò)增目的基因前500 bp片段,膠回收后得到目的片段產(chǎn)物。
使用同源重組酶連接線性化載體片段和目的片段,制作JM109化轉(zhuǎn)感受態(tài)細(xì)胞,將連接好的質(zhì)粒利用化學(xué)轉(zhuǎn)化法轉(zhuǎn)入JM109感受態(tài)細(xì)胞中,37 ℃、200 r/min搖床孵育1 h后,5 000 r/min離心5 min后去掉上清,用剩余少許培養(yǎng)基將菌體重懸,涂至含有氯霉素抗性的LB固體平板上。挑選氯霉素抗性平板上長出的單菌落,以引物CX-P-1/CX-P-2進(jìn)行菌落PCR驗(yàn)證,驗(yàn)證成功的單菌落送于天霖生物科技(無錫)有限公司測(cè)序,測(cè)序成功的質(zhì)粒化轉(zhuǎn)入E.coliW3110中進(jìn)行下一步試驗(yàn)。
1.3.4 24孔板培養(yǎng)
24孔板裝液量為2 mL,將培養(yǎng)至對(duì)數(shù)期的菌種以2%的接種量接入24孔板的LB培養(yǎng)基中(含不同質(zhì)量濃度的L-丙氨酸),37 ℃、600 r/min振蕩培養(yǎng)8 h。
1.3.5 相對(duì)熒光強(qiáng)度測(cè)定
取1 mL待測(cè)樣品,8 000 r/min離心1 min,用相同體積的PBS清洗兩次,然后用PBS稀釋OD600nm值在0.4~0.6左右,取200 μL稀釋后樣品加入黑色避光的酶標(biāo)板中,使用酶標(biāo)儀測(cè)量熒光強(qiáng)度,設(shè)定激發(fā)波長為588 nm,發(fā)射波長為645 nm,并測(cè)定OD600nm值,根據(jù)熒光強(qiáng)度和OD600nm值的比值計(jì)算相對(duì)熒光強(qiáng)度[27]。
1.3.6 數(shù)據(jù)處理
試驗(yàn)數(shù)據(jù)用Microsoft Excel 2016進(jìn)行整理和分析,使用Origin 2021作圖。
2 結(jié)果與分析
2.1 L-丙氨酸壓力下高轉(zhuǎn)錄啟動(dòng)子的篩選
轉(zhuǎn)錄水平在L-丙氨酸壓力下比正常條件下升高基因的啟動(dòng)子可能是由L-丙氨酸驅(qū)動(dòng)的啟動(dòng)子[28]。從測(cè)序結(jié)果給出的差異表達(dá)基因分析列表中,選擇轉(zhuǎn)錄水平較對(duì)照組提高水平靠前的10個(gè)基因(ArgK,F(xiàn)lgG,GlgP,CpxP,BetA,GlpA,KdpB,PulG,LivH和YhfX)作為候選基因,結(jié)果見圖1。
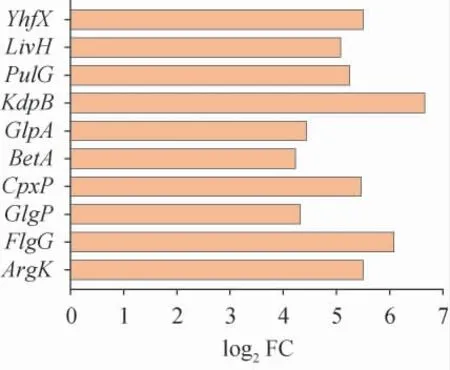
圖1 十個(gè)候選啟動(dòng)子的差異表達(dá)Fig.1 Differential expression of ten candidate promoters
由圖1可知,轉(zhuǎn)錄水平提高最多的基因?yàn)镵dpB,轉(zhuǎn)錄水平差異為6.66,其次是FlgG,轉(zhuǎn)錄水平差異為6.08。
使用蛋白質(zhì)直系同源簇?cái)?shù)據(jù)庫(cluster of orthologous groups of proteins,COG)查詢分析10個(gè)候選基因的基因功能,結(jié)果見表2。由表2可知,該10個(gè)基因的功能分別屬于細(xì)胞運(yùn)動(dòng)和分泌、氨基酸運(yùn)輸與代謝、碳水化合物運(yùn)輸和代謝、無機(jī)離子轉(zhuǎn)運(yùn)和代謝和能量產(chǎn)生和轉(zhuǎn)化,將這10個(gè)基因的啟動(dòng)子分別編號(hào)為PAla-7~PAla-10。
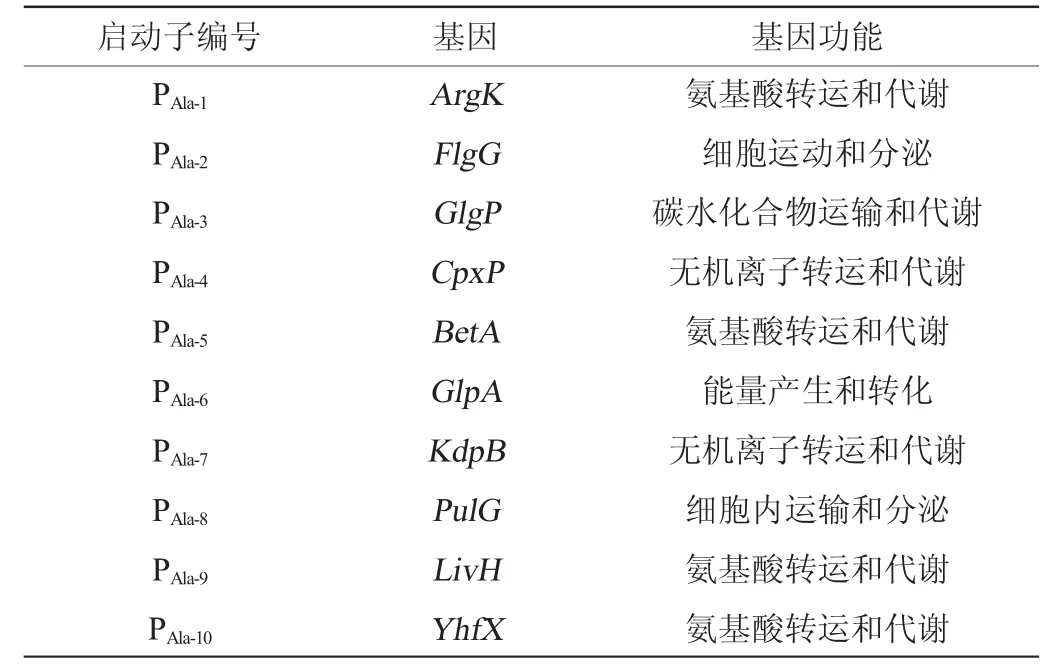
表2 10個(gè)轉(zhuǎn)錄水平提高的基因Table 2 10 genes with increased transcriptional level
2.2 熒光報(bào)告菌株的構(gòu)建
為了構(gòu)建含有紅色熒光蛋白基因的質(zhì)粒,以pTET質(zhì)粒作為骨架質(zhì)粒,以實(shí)驗(yàn)室已有的mKate基因片段為模板[29],設(shè)計(jì)含同源臂的引物mKate-1和mKate-2擴(kuò)增mKate基因片段,得到目的基因片段;以含有同源臂的引物pTET-1和pTET-2擴(kuò)增pTET質(zhì)粒,得到線性化載體片段,按方法1.3.3進(jìn)行質(zhì)粒構(gòu)建。挑選五個(gè)氯霉素抗性平板上長出的pTETmKate質(zhì)粒重組菌單菌落進(jìn)行菌落PCR驗(yàn)證,結(jié)果見圖2。由圖2可知,1號(hào)、2號(hào)、4號(hào)和5號(hào)泳道PCR擴(kuò)增產(chǎn)物堿基長度約為1 000 bp,與mKate基因大小相符,說明mKate基因成功連接到了pTET質(zhì)粒上。將菌落PCR驗(yàn)證正確的菌株送至天霖生物科技(無錫)有限公司測(cè)序,測(cè)序成功后得到pTET-mKate質(zhì)粒。
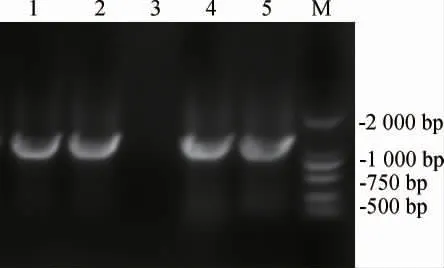
圖2 pTET-mKate質(zhì)粒重組菌菌落PCR驗(yàn)證Fig.2 PCR verification of pTET-mKate plasmid recombinant colony
基因的前500 bp可能包含潛在的啟動(dòng)子序列,為了更直觀的表征啟動(dòng)子的轉(zhuǎn)錄水平,篩選響應(yīng)L-丙氨酸濃度的啟動(dòng)子,通過京都基因與基因組百科全書數(shù)據(jù)庫(Kyoto encyclopedia of genes and genomes,KEGG)和美國國家生物技術(shù)信息中心數(shù)據(jù)庫(national center of biotechnology information,NCBI)查詢并預(yù)測(cè)10個(gè)候選基因前500 bp含有潛在啟動(dòng)子的序列。以E.coliW3110的基因組為模板,設(shè)計(jì)同源臂引物,將轉(zhuǎn)錄組分析篩選出的10個(gè)候選基因的前500 bp進(jìn)行擴(kuò)增,利用同源重組連接pTET-mKate質(zhì)粒的紅色熒光蛋白基因mKate,質(zhì)粒圖譜見圖3。由圖3可知,將10個(gè)候選基因的啟動(dòng)子序列插入到mKate基因前面,即可得到不同啟動(dòng)子控制的mKate基因表達(dá)質(zhì)粒。

圖3 熒光蛋白表達(dá)質(zhì)粒構(gòu)建Fig.3 Construction of fluorescent protein expression plasmid
挑取氯霉素抗性平板長出的單菌落進(jìn)行菌落PCR鑒定,結(jié)果見圖4。由圖4可知,PCR擴(kuò)增產(chǎn)物堿基長度約為1500bp,說明500 bp的啟動(dòng)子片段成功連接到了pTET-mKate質(zhì)粒上。將驗(yàn)證成功的菌株送至天霖生物科技(無錫)有限公司測(cè)序,測(cè)序成功后提取重組質(zhì)粒化學(xué)轉(zhuǎn)化入E.coliW3110感受態(tài)細(xì)胞中,所獲得的菌株分別命名為P1~P10。
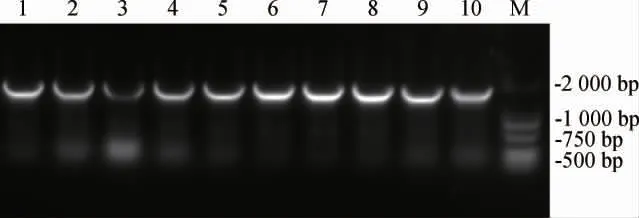
圖4 重組菌株啟動(dòng)子菌落PCR驗(yàn)證Fig.4 PCR verification of recombinant strains promoter colony
2.3 重組菌株相對(duì)熒光強(qiáng)度測(cè)定
為了比較不同菌株在L-丙氨酸壓力下的相對(duì)熒光強(qiáng)度,將培養(yǎng)至對(duì)數(shù)期的菌株接種到未添加L-丙氨酸和添加10 g/LL-丙氨酸的LB培養(yǎng)基中,培養(yǎng)8 h后取樣測(cè)定熒光強(qiáng)度和OD600nm值,通過熒光強(qiáng)度和OD600nm值的比值計(jì)算相對(duì)熒光強(qiáng)度,結(jié)果見圖5。由圖5可知,PAla-4、PAla-8和PAla-10啟動(dòng)子顯示出了較高的相對(duì)熒光強(qiáng)度,在不添加L-丙氨酸的LB培養(yǎng)基中培養(yǎng)8 h后,相對(duì)熒光強(qiáng)度分別達(dá)到1 421、1 121和1 226,但是在添加10 g/LL-丙氨酸的LB培養(yǎng)基中培養(yǎng)后,相對(duì)熒光強(qiáng)度分別下降了21.2%、18.4%和51.5%,說明PAla-4、PAla-8和PAla-10啟動(dòng)子并不會(huì)響應(yīng)L-丙氨酸濃度來提高基因的轉(zhuǎn)錄水平。PAla-7和PAla-9啟動(dòng)子的熒光報(bào)告菌株在添加10g/LL-丙氨酸壓力下的相對(duì)熒光強(qiáng)度均高于未添加L-丙氨酸,分別是未添加L-丙氨酸相對(duì)熒光強(qiáng)度的1.5倍和3.8倍。其他啟動(dòng)子菌株在添加10 g/LL-丙氨酸壓力下的相對(duì)熒光強(qiáng)度均低于未添加L-丙氨酸,可能是L-丙氨酸會(huì)抑制熒光蛋白基因的表達(dá)。因此,選擇PAla-7和PAla-9啟動(dòng)子進(jìn)行下一步的評(píng)估驗(yàn)證。

圖5 10個(gè)啟動(dòng)子菌株在10 g/L L-丙氨酸質(zhì)量濃度下相對(duì)熒光強(qiáng)度比較Fig.5 Comparison of relative fluorescence intensity of 10 promoters strains at mass concentration of 10 g/L L-alanine
2.4 啟動(dòng)子響應(yīng)L-丙氨酸濃度的靈敏性分析
將含有PAla-7和PAla-9啟動(dòng)子的菌株P(guān)7和P9分別接種到添加0~60 g/LL-丙氨酸的LB培養(yǎng)基中進(jìn)行誘導(dǎo),觀察PAla-7和PAla-9啟動(dòng)子對(duì)不同L-丙氨酸濃度響應(yīng)的靈敏性,結(jié)果見圖6。

圖6 PAla-7和PAla-9啟動(dòng)子在不同L-丙氨酸質(zhì)量濃度下相對(duì)熒光強(qiáng)度比較Fig.6 Comparison of relative fluorescence intensity of PAla-7 and PAla-9 promoters at different L-alanine mass concentrations
由圖6可知,在不同濃度L-丙氨酸的誘導(dǎo)下,PAla-9啟動(dòng)子并未隨著L-丙氨酸濃度的增加而增加。在不同濃度L-丙氨酸的誘導(dǎo)下,PAla-7啟動(dòng)子的相對(duì)熒光強(qiáng)度隨著L-丙氨酸濃度的而增加,相對(duì)于未添加L-丙氨酸,添加10 g/L、20 g/L、30 g/L、40 g/L、50 g/L和60 g/LL-丙氨酸誘導(dǎo)后的相對(duì)熒光強(qiáng)度分別增加了49%、122%、181%、275%、432%和705%,PAla-7啟動(dòng)子在0~60 g/LL-丙氨酸誘導(dǎo)下顯示出了良好的靈敏性。
2.5 啟動(dòng)子響應(yīng)L-丙氨酸濃度的特異性分析
為了分析PAla-7啟動(dòng)子是否對(duì)L-丙氨酸具有特異性響應(yīng),選擇11種常見的氨基酸(甲硫氨酸、丙氨酸、蘇氨酸、纈氨酸、脯氨酸、賴氨酸、半胱氨酸、亮氨酸、苯丙氨酸、甘氨酸、精氨酸)分別進(jìn)行誘導(dǎo)。將培養(yǎng)至對(duì)數(shù)期的菌株P(guān)7接種至含10 g/L和20 g/L的不同氨基酸的LB培養(yǎng)基中,測(cè)定誘導(dǎo)后的相對(duì)熒光強(qiáng)度,結(jié)果見圖7。由圖7可知,L-丙氨酸誘導(dǎo)質(zhì)量濃度從10 g/L增加到20 g/L時(shí),相對(duì)熒光強(qiáng)度增加了52.4%,而其他氨基酸隨著濃度的增加,相對(duì)熒光強(qiáng)度并沒有隨著有明顯的增加。因此,PAla-7啟動(dòng)子對(duì)L-丙氨酸具有特異性響應(yīng)。
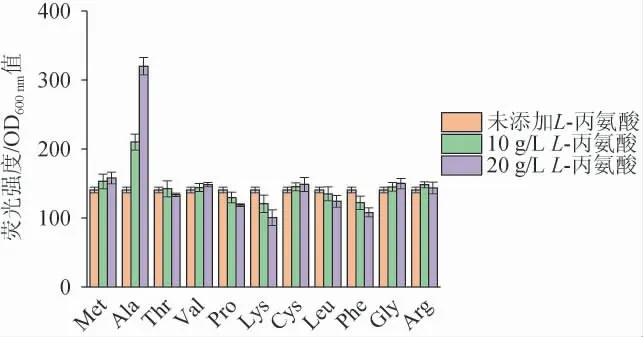
圖7 PAla-7啟動(dòng)子在不同種類氨基酸下相對(duì)熒光強(qiáng)度比較Fig.7 Comparison of relative fluorescence intensity of PAla-7 promoter under different kinds of amino acids
3 結(jié)論
通過對(duì)不同質(zhì)量濃度L-丙氨酸壓力下的大腸桿菌進(jìn)行轉(zhuǎn)錄組學(xué)分析,從中篩選了可能是L-丙氨酸誘導(dǎo)的10個(gè)候選基因的啟動(dòng)子。隨后,利用紅色熒光蛋白進(jìn)行表征,篩選得到了特異性響應(yīng)L-丙氨酸濃度的PAla-7啟動(dòng)子,該啟動(dòng)子在0~60 g/LL-丙氨酸范圍內(nèi)具有良好的靈敏性。值得注意的是,其余啟動(dòng)子雖不能響應(yīng)L-丙氨酸濃度表達(dá),但可作為組成型啟動(dòng)子,用于后續(xù)啟動(dòng)子優(yōu)化、調(diào)控基因表達(dá)水平和平衡代謝流等研究。該研究篩選得到的PAla-7啟動(dòng)子可作為生物傳感器的調(diào)控元件,也為后續(xù)L-丙氨酸動(dòng)態(tài)調(diào)控系統(tǒng)的構(gòu)建提供了理論基礎(chǔ)。
