大黃魚(Larimichthys crocea)幼魚同生群內(nèi)不同增重性能子群間的形質(zhì)差異*
2023-10-17 07:12:38李有志蔣宏雷劉哲宇斯烈鋼王志錚
海洋與湖沼 2023年5期
關(guān)鍵詞:能力
李有志 蔣宏雷 劉哲宇 曲 濤 江 柳 斯烈鋼, 王志錚①
(1.浙江海洋大學(xué)水產(chǎn)學(xué)院 浙江舟山 316022; 2.寧波市海洋與漁業(yè)研究院 浙江寧波 315103)
大黃魚(Larimichthys crocea)隸屬于硬骨魚綱(Osteichthyes)、鱸形目(Perciformes)、石首魚科(Sciaenidae), 與小黃魚(Larimichthys polyactis)、帶魚(Trichiurus lepturus)、曼氏無針烏賊(Sepiella japonica)并稱舟山漁場四大傳統(tǒng)知名漁產(chǎn), 肉鮮味美, 營養(yǎng)經(jīng)濟(jì)價值高, 素有“海水國魚”之美譽。自20 世紀(jì)90年代初以來, 隨著規(guī)模化全人工養(yǎng)殖關(guān)鍵技術(shù)的不斷突破與相關(guān)配套裝備的不斷提升, 我國大黃魚增養(yǎng)殖產(chǎn)業(yè)獲得了蓬勃發(fā)展, 現(xiàn)已成為當(dāng)前我國最為重要的海水魚類養(yǎng)殖對象和近海增殖漁業(yè)的當(dāng)家魚種(江柳等, 2021)。據(jù)《2023 中國漁業(yè)統(tǒng)計年鑒》(農(nóng)業(yè)農(nóng)村部漁業(yè)漁政管理局等, 2023)報道, 我國2022年大黃魚養(yǎng)殖產(chǎn)量達(dá)25.768 3 萬t, 居全國主要海水養(yǎng)殖魚類首位, 捕撈產(chǎn)量達(dá)3.709 8 萬t, 放流效果顯著。
種內(nèi)個體差異是自然界中普遍存在的現(xiàn)象, 尤以個體大小為代表的表型差異最為直觀且常見。表型受遺傳因子和環(huán)境因子的共同影響(Mayret al, 1953),作為物種在種質(zhì)可塑性范圍內(nèi)對特定環(huán)境的形態(tài)修飾(Robinsonet al, 1996; Wagneret al, 2001; Killenet al, 2011), 是劃分并闡釋水產(chǎn)動物同生群內(nèi)不同抗逆等級子群間生存對策差異的重要生物學(xué)依據(jù)(王志錚等, 2016a, 2016b, 2017, 2018)。故, 大黃魚同生群仔魚的個體大小差異有隨養(yǎng)殖時間推移而逐漸放大的趨勢(竺俊全等, 2004), 不同生長性能大黃魚養(yǎng)殖群體間在腸道菌群種類組成、致病菌含量和肌肉生長調(diào)控基因表達(dá)上均存在顯著差異(李英英等, 2017; 張波等,2023), 以及形態(tài)表型在辨析不同地理居群、養(yǎng)殖條件、遺傳特征和不同性別大黃魚上具重要指示作用(黃良敏等, 2006; 丁文超等, 2009; 諶微等, 2014; 王映等, 2016; 沈偉良等, 2017)等的研究結(jié)果, 均無疑為探究大黃魚同生群內(nèi)不同增重性能子群間形質(zhì)特征差異的內(nèi)在邏輯, 進(jìn)而揭示造成各子群間生存適應(yīng)對策分化的主因提供了重要線索和啟示。鑒于此,基于苗種質(zhì)量是決定水產(chǎn)增養(yǎng)殖產(chǎn)業(yè)健康可持續(xù)發(fā)展的關(guān)鍵, 而國內(nèi)外迄今尚未見有關(guān)大黃魚苗種質(zhì)量等級劃分與評價方法研究的系統(tǒng)報道之實際, 本研究團(tuán)隊于2019 年8 月以寧波市象山西滬港海域板式網(wǎng)箱內(nèi)養(yǎng)殖的同生群大黃魚幼魚為研究對象, 較系統(tǒng)開展了不同增重性能子群間形質(zhì)差異的研究,以期為精選大黃魚優(yōu)質(zhì)增殖群體和指導(dǎo)大黃魚幼魚階段的科學(xué)高效養(yǎng)殖提供參考。
1 材料與方法
1.1 實驗用魚
寧波象山港灣水產(chǎn)苗種有限公司育苗車間出池后,在象山西滬港海域的板式網(wǎng)箱(規(guī)格: 5 m×3 m×6 m)內(nèi)常規(guī)養(yǎng)殖3 個月的大黃魚同生群幼魚。隨機(jī)撈取形體完整、無病無傷的健壯個體3 000 ind., 立即充氣并停食暫養(yǎng)于若干直徑1 m、高1.5 m 的白色塑料桶(實際水位1.3 m)內(nèi)。2 d 后, 用BSA223S 電子天平(賽多利斯科學(xué)儀器有限公司, 精度1 mg)逐尾稱量體質(zhì)量,并根據(jù)農(nóng)業(yè)試驗研究中的小概率標(biāo)準(zhǔn)(P=0.05) (蓋鈞鎰, 2000), 按其出現(xiàn)率將劃分為A、B、C 三個增重性能子群(圖1)。
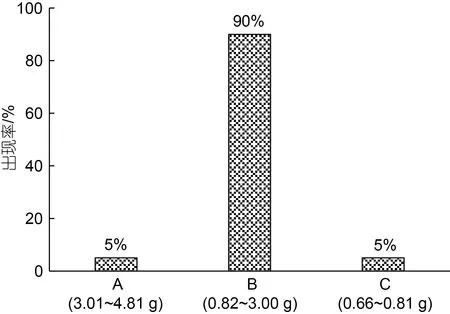
圖1 實驗魚各子群的出現(xiàn)率及其體質(zhì)量范圍Fig.1 The occurrence rate and body weight range of each subgroup of experimental fish
1.2 生物學(xué)性狀的測定
每一子群各隨機(jī)選取30 ind.作為生物學(xué)測定樣本。吸水紙抹凈體表水分后, 借助數(shù)顯游標(biāo)卡尺(精度0.01 mm, 桂林廣陸數(shù)字測控股份有限公司)和BSA223S電子天平逐尾測定體長(X1)、體寬(X2)、體高(X3)、肛長(X4)、頭長(X5)、頭寬(X6)、頭高(X7)、眼后頭長(X8)、眼徑(X9)、鰓蓋高(X10)、背鰭部體高(X11)、腹鰭間距(X12)、側(cè)線長(X13)、尾柄長(X14)、尾柄高(X15)等15項體尺性狀(具體測量方法見圖2 和表1)和體質(zhì)量(BW)、凈體質(zhì)量(NW, 去除鰓和內(nèi)臟后的體質(zhì)量)、內(nèi)臟質(zhì)量(W1)、鰓質(zhì)量(W2)等4 項質(zhì)量性狀。

表1 實驗魚體尺測量性狀的定義Tab.1 Definition of body size measurement traits of experimental fish
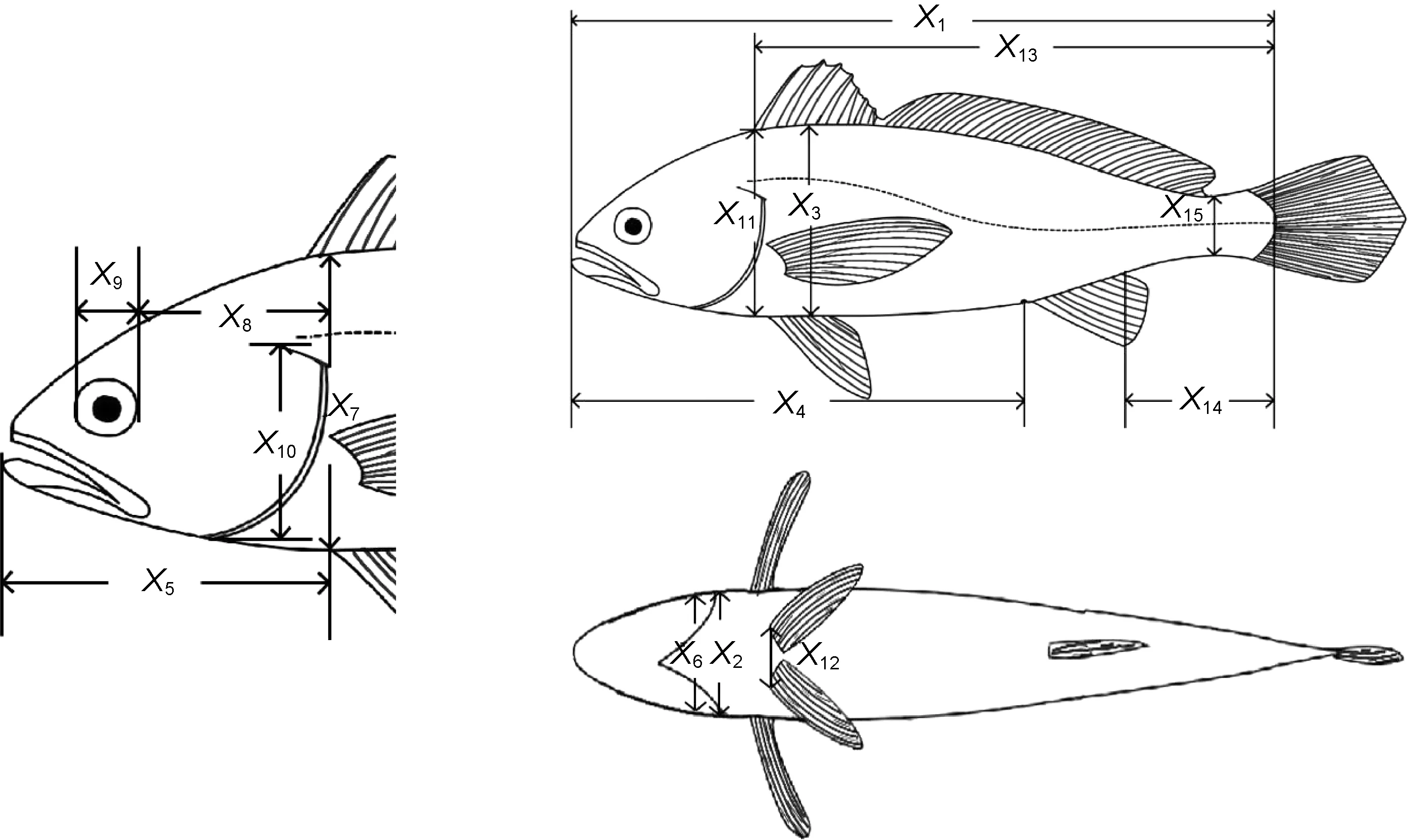
圖2 實驗魚體尺性狀測量示意圖Fig.2 Pictures showing the measurements of the body size traits of experimental fish
1.3 數(shù)據(jù)處理
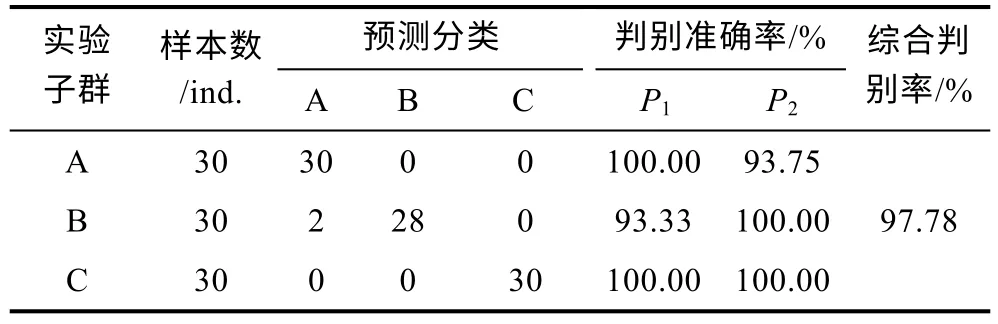
根據(jù)所得測定數(shù)據(jù), 采用SPSS24.0 軟件計算本研究所涉3 個實驗子群的各項生物學(xué)測定性狀和形質(zhì)評價指標(biāo)的均值和標(biāo)準(zhǔn)差, 并檢驗組間差異顯著性(視P<0.05 為顯著水平); 根據(jù)形質(zhì)評價指標(biāo)的均值,在計算3 個實驗子群間歐氏距離的基礎(chǔ)上, 進(jìn)行聚類分析; 按曲濤等(2022)的方法就形質(zhì)評價指標(biāo)進(jìn)行主成分分析和判別分析, 并參照Li 等(2005)的方法計算判別準(zhǔn)確率和綜合判別率以驗證所建Fisher分類函數(shù)方程組的可靠性。
2 結(jié)果
2.1 生物學(xué)測定性狀
由表2 可見, 除X15呈A>B≈C 外, 其余18 項性狀均呈A>B>C (P<0.05), 表明A、B、C 子群在生物學(xué)測定性狀上具良好的區(qū)分度, 即大黃魚幼魚的生物學(xué)性狀測定值有隨其增重性能增強(qiáng)而顯著增大的趨勢, 反之亦然。
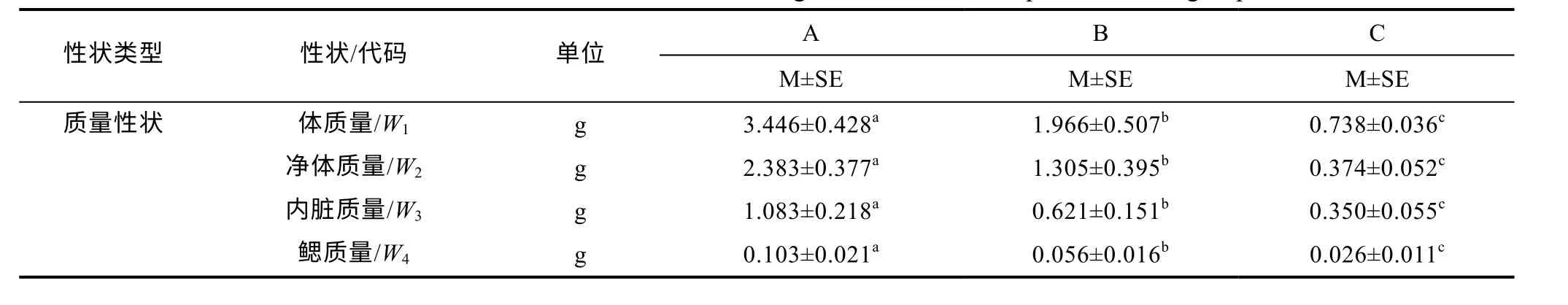
表2 各實驗子群生物學(xué)測定性狀的均值和標(biāo)準(zhǔn)差(n=30)Tab.2 Means and standard deviations of biological traits in each experimental subgroup
2.2 形質(zhì)評價指標(biāo)
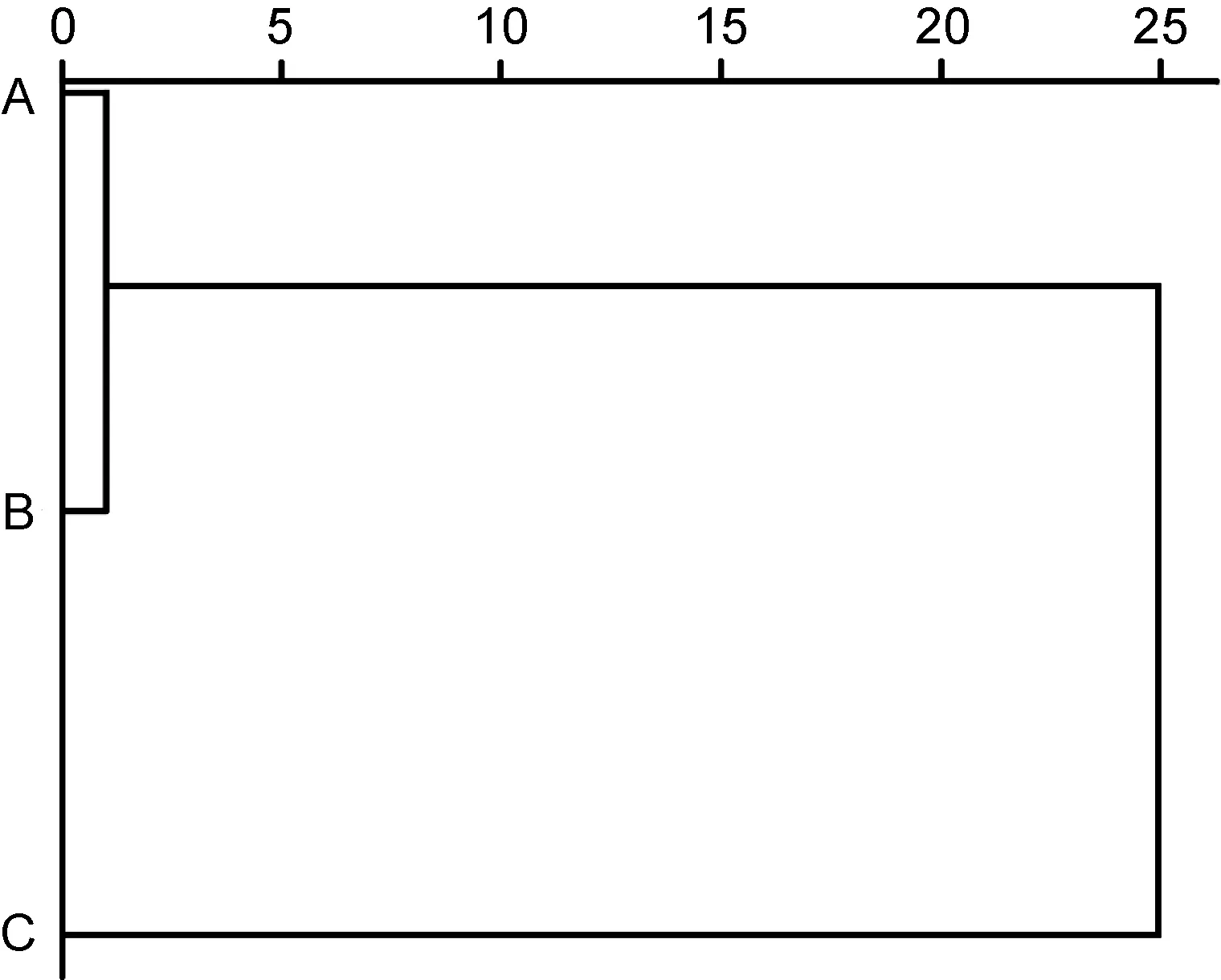
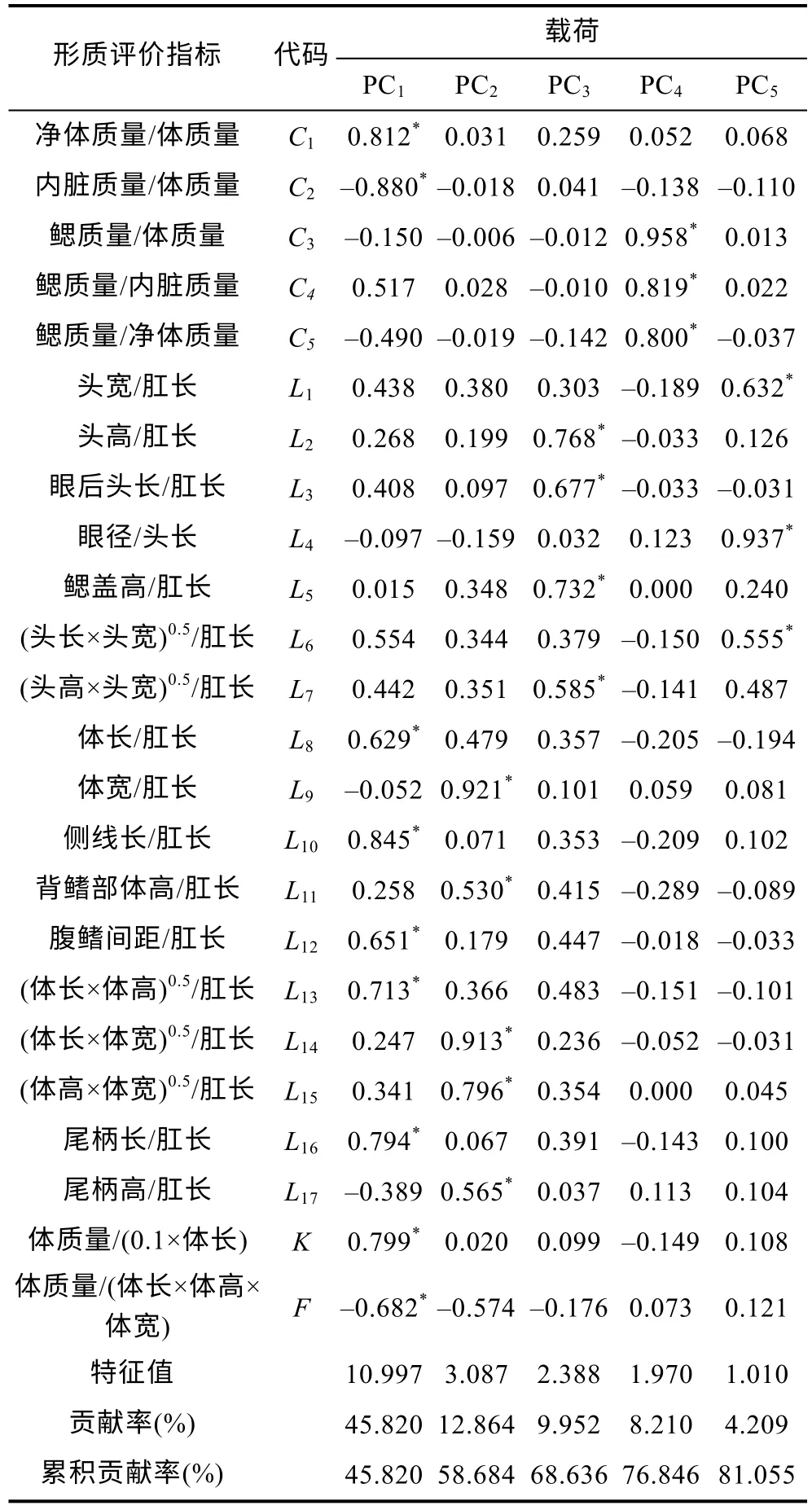
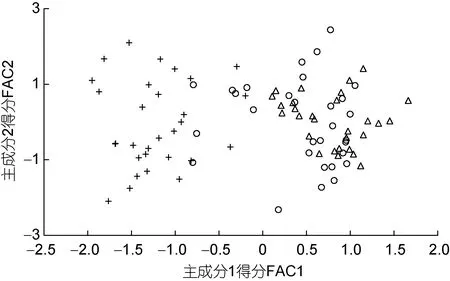
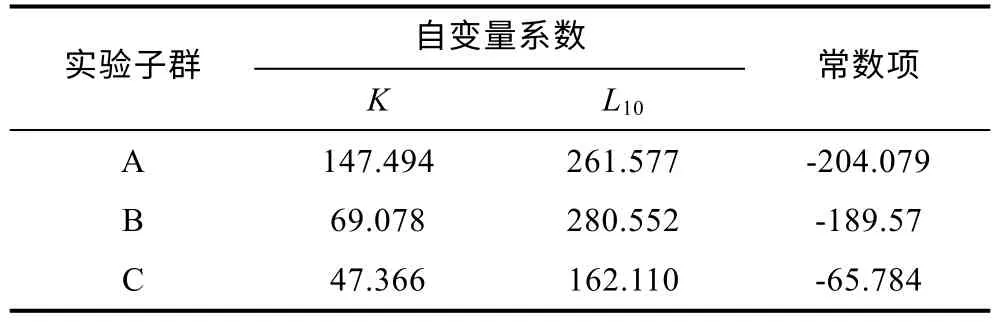
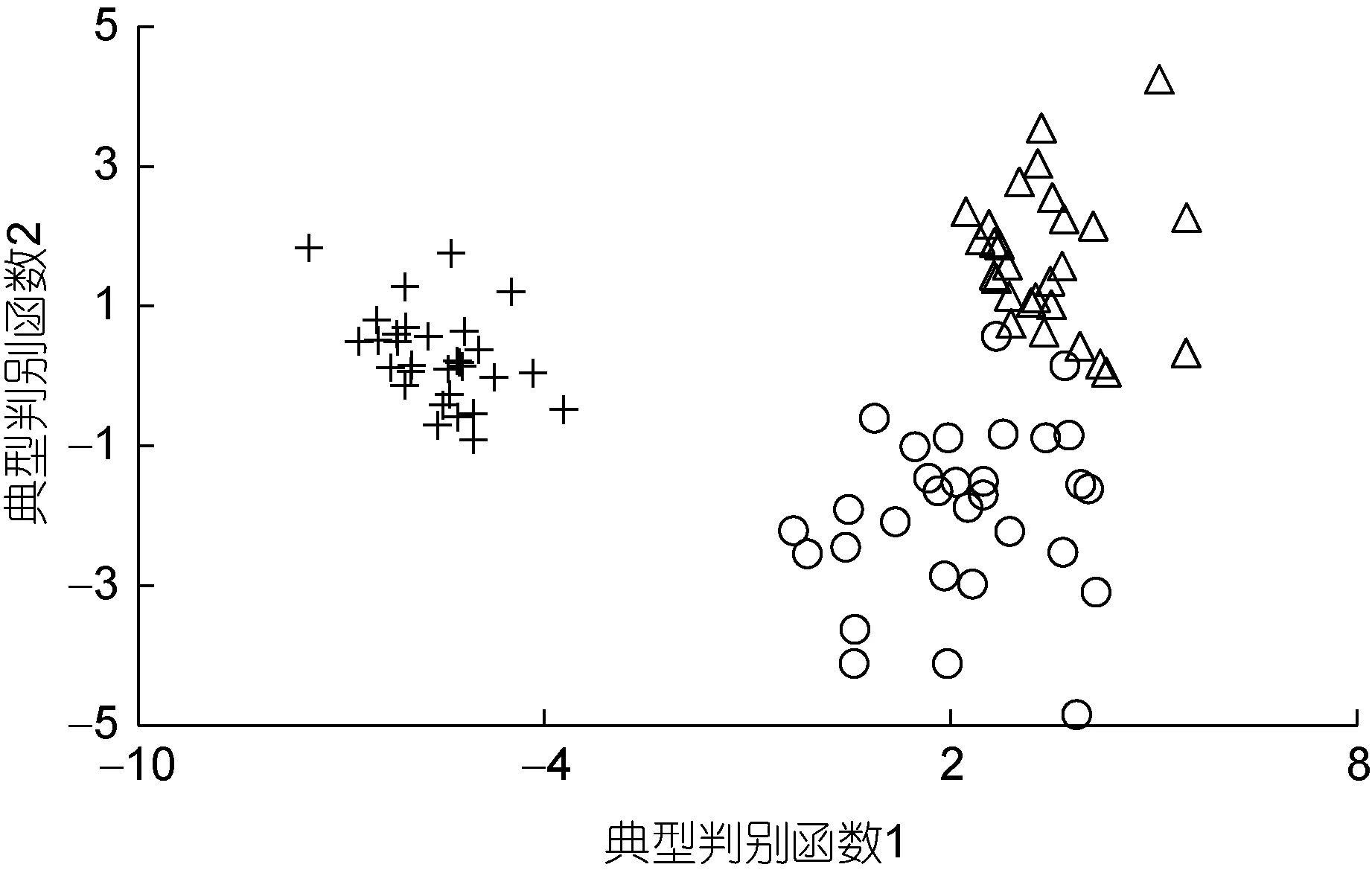
由表3 可見, 各子群間在本研究所涉形質(zhì)評價指標(biāo)上的異同表現(xiàn)為: (1) 5 項質(zhì)量比例中, 除C1、C4呈A≈B>C 外, 其余指標(biāo)均呈A≈B 表3 各實驗子群形質(zhì)評價指標(biāo)的均值和標(biāo)準(zhǔn)差(n=30)Tab.3 Means and standard deviations of the morphological and quality evaluation indicators in each experimental subgroup 由圖3 可見, A、B 子群間基于各項形質(zhì)評價指標(biāo)均值的歐式距離最小, 首先被聚為一支, 而后再與C子群聚在一起, 表明子群C 的形質(zhì)特征與A、B 子群均已出現(xiàn)明顯乖離, 即絕大多數(shù)實驗個體的形質(zhì)特征有向高增重性能等級子群趨同的傾向。 圖3 各實驗子群間的聚類圖Fig.3 Clustering diagram of each experimental subgroup 經(jīng)檢驗, 本研究所涉各項形質(zhì)評價指標(biāo)的相關(guān)系數(shù)與單位矩陣間具極顯著差異(P<0.01)且適合度尚可(KMO=0.765), 有進(jìn)行因子分析的必要。相關(guān)結(jié)果見表4。經(jīng)分析, 所獲特征根值大于1 的5 個主成分,其方差累積貢獻(xiàn)率達(dá) 81.055%, 表明它們?yōu)榭筛爬ˋ、B、C 子群間形質(zhì)特征差異的公共因子。其中, PC1特征根值10.997, 方差貢獻(xiàn)率45.820%, 主要影響變量(載荷絕對值大于0.5) 9 個, 絕對值排序為C2>L10>C1>K>L16>L13>F>L12>L8; PC2特征根值3.087, 方差貢獻(xiàn)率12.864%, 主要影響變量5 個, 排序為L9>L14>L15>L17>L11; PC3特征根值2.388, 方差貢獻(xiàn)率9.952%,主要影響變量4 個, 排序為L2>L5>L3>L7; PC4與PC5的特征根值分別為1.970 和1.010, 方差貢獻(xiàn)率分別為8.210%和4.209%, 主要影響變量均為3 個, 排序分別為C3>C4>C5和L4>L1>L6。由此可知, 特征根值、方差貢獻(xiàn)率、主要影響變量個數(shù)自PC1至PC5均表露出由急劇下降至趨于穩(wěn)定的態(tài)勢, 表明PC1在甄別A、B、C 子群間形質(zhì)差異上較其他主成分具極端重要性。繪制PC1和PC2得分散布圖, 得圖4。由圖4 可見, A、B、C 沿FAC1軸自右向左雖大體上均各自占據(jù)獨立的區(qū)域, 但B 子群與A、C 子群間均存在一定程度重疊,尤以A、B 間為甚, 這既真實反映了B 子群為A、C子群間的過渡群體, 以及B 子群形質(zhì)特征有明顯向A子群靠攏的傾向的事實, 也客觀指示了PC1中含有分辨A、B 和B、C 子群間形質(zhì)特征的干擾變量。故, 需進(jìn)一步做判別分析以進(jìn)一步提高甄別A、B、C 子群的分辨率。 表4 各實驗子群的主成分分析Tab.4 PCA of each experimental subgroup 圖4 主成分得分散布圖Fig.4 The scatter diagram of principle component 經(jīng)逐步導(dǎo)入剔除法, 篩留判別貢獻(xiàn)率較大的重長系數(shù)K和L10(側(cè)線長/肛長)進(jìn)行判別分析。F檢驗亦表明K、L10均達(dá)到極顯著水平(P<0.01), 并據(jù)此建立用于判別A、B、C 子群的Fisher分類函數(shù)方程組于表5。經(jīng)計算, A、B、C 子群的判別準(zhǔn)確率P1依次為100.00%、93.33%和 100.00%,P2依次為 93.75%、100.00%和100.00%, 綜合判別準(zhǔn)確率為97.78% (表6)。與此同時,圖5 所示典型判別函數(shù)得分散布圖, 無疑更直觀證實了上述分類判別結(jié)果的可靠性。 表5 Fisher 分類函數(shù)方程組自變量系數(shù)及常數(shù)項Tab.5 Coefficients and constant terms of independent variables of Fisher classification function equations 表6 判別函數(shù)分類結(jié)果Tab.6 Classification results of discriminant function 圖5 各實驗子群的典型判別函數(shù)散布圖Fig.5 Typical discriminant function scatter plots of each experimental subgroup 魚類網(wǎng)箱養(yǎng)殖模式具魚群擁擠度高、飼料投喂頻次少且時長短的特點, 故養(yǎng)殖于網(wǎng)箱內(nèi)的魚類常處于高烈度種內(nèi)食物競爭狀態(tài), 致使能優(yōu)先獲得食物的個體往往能在種內(nèi)競爭中處于強(qiáng)勢地位, 反之亦然。盡可能增加獲利并減少消耗, 是生物選擇并實施增重對策的底層邏輯。因此, 養(yǎng)殖于網(wǎng)箱內(nèi)的魚類同生群增重性能等級的分野勢必與其在食物競爭中所選擇的“獲利-消耗”模式有著極為密切的關(guān)系。無疑, 本研究所涉A、B、C 子群間形質(zhì)特征的差異(表3), 應(yīng)與它們針對所處網(wǎng)箱養(yǎng)殖生境選擇與自身競食能力相適配的“獲利-消耗”模式密切相關(guān)。即: (1) 重長系數(shù)K作為衡量單位體長生物量的評價指標(biāo), 是表征機(jī)體營養(yǎng)狀況和體質(zhì)強(qiáng)壯程度的重要形質(zhì)變量。感覺系統(tǒng)在魚類捕食過程中起著至關(guān)重要的作用(Evanset al, 2004; Coombset al, 2014), 側(cè)線感覺系統(tǒng)不僅能感知和定位獵物(梁旭方, 1998), 而且還具有過濾或消除嘈雜環(huán)境噪音的功能(Engelmannet al, 2000)。由此, 本研究中K為唯一呈A>B>C (P<0.05)的形質(zhì)評價指標(biāo), 以及K、L10組合成為判別A、B、C 子群的關(guān)鍵變量組合的結(jié)果(表3, 表5), 既揭示了以體質(zhì)強(qiáng)壯程度和競食靈敏度為代表的食物搶奪能力在表征大黃魚幼魚增重性能等級上的極端重要性的同時,也映射了切實提高食物保障水平是大黃魚幼魚權(quán)衡“獲利-消耗”的優(yōu)先選項; (2) “器官代謝活性假說”指出, 活性器官的臟器系數(shù)與機(jī)體代謝水平呈正相關(guān),而惰性組織與體質(zhì)量的比例則與此相反(Itazawaet al,1983; Oikawaet al, 1992)。故, 表3 所示C2(內(nèi)臟質(zhì)量/體質(zhì)量)、C3(鰓質(zhì)量/體質(zhì)量)和C5(鰓質(zhì)量/凈體質(zhì)量)均呈A≈B 游泳運動作為支持魚類捕食活動的重要生理行為, 其運動能力與運動方式勢必與所采取的攝食策略有著極為密切的關(guān)聯(lián), 并將深刻影響其增重機(jī)制。據(jù)報道, 魚類在持續(xù)運動能力與暴發(fā)運動能力之間存在權(quán)衡, 即當(dāng)其持續(xù)運動能力獲得顯著增強(qiáng)時, 暴發(fā)運動能力則可能會變?nèi)? 反之亦然; 對于海洋洄游性魚類而言, 需具較強(qiáng)的持續(xù)游泳能力(傅世建等,2008)。研究發(fā)現(xiàn), 體高的顯著增大有助于提升中華倒刺 鲃和大黃魚等紡錘體型魚類的高速游泳能力(李秀明等, 2013; 沈偉良等, 2017)。故, 陳慧等(2007)報道的網(wǎng)箱養(yǎng)殖大黃魚體高與體長間的比值普遍大于野生大黃魚, 以及王映等(2016)報道的室內(nèi)人工養(yǎng)殖大黃魚體高與體長間的比值顯著大于野生大黃魚的結(jié)果, 均指示了暴發(fā)游泳較持續(xù)運動對促進(jìn)養(yǎng)殖大黃魚生長更具重要性的事實。由此, 結(jié)合本研究分析所得A、B、C 子群的攝食策略差異, 可導(dǎo)出網(wǎng)箱養(yǎng)殖生境下大黃魚幼魚的暴發(fā)運動在降低機(jī)體代謝能耗、提高搶奪食物成功率和優(yōu)先獲得食物保障上較持續(xù)運動更具增重生理生態(tài)重要性的推論, 即暴發(fā)游泳運動能力越強(qiáng), 增重越快, 持續(xù)游泳運動能力越強(qiáng),則增重越慢, 反之亦然。故, 可判定持續(xù)運動和暴發(fā)運動在運動量上占比的多寡是導(dǎo)致A、B、C 子群增重性能差異的重要原因。 經(jīng)剖析, 本研究通過對形質(zhì)特征進(jìn)行降維處理獲得的5 個相互獨立的公共因子PC1、PC2、PC3、PC4和PC5(表4), 在揭示引起A、B、C 子群間形質(zhì)特征和攝食策略差異的真實原因的同時, 也進(jìn)一步支持了持續(xù)運動和暴發(fā)運動量占比是導(dǎo)致三者增重性能差異的重要原因的可靠性。即: (1) 魚類的游泳能力與其體長、體型以及鰭的發(fā)達(dá)程度均密切相關(guān)(Webb,1977; Peakeet al, 1998)。據(jù)報道, 絕大多數(shù)魚類游泳時主要通過魚體兩側(cè)或腹背肌肉的交替收縮, 由身體前部向尾部依次收縮各肌肉群, 使魚尾擺動來獲得向前的推動力(周應(yīng)祺, 1985), 并通過背鰭、臀鰭和腹鰭的協(xié)助來保持魚體平衡(李林春, 2009)。因此, 由指示機(jī)體代謝強(qiáng)度的C1和C2、表征暴發(fā)游泳能力的L8、L13和L16、維系魚體游泳平衡能力的L1、反映機(jī)體生物量積累水平的F, 與被篩留的判別變量組合K、L10共同組成的PC1主要影響變量(表4, 表5), 既集中反映了在網(wǎng)箱養(yǎng)殖生境下大黃魚幼魚的暴發(fā)運動能力、魚體平衡能力、食物感知與定位能力、體質(zhì)強(qiáng)壯程度和機(jī)體營養(yǎng)狀況對顯著提升競食能力的支撐作用, 也映射了提升競食有效性對降低機(jī)體代謝能耗的助益效應(yīng)。由此表明, 獲取食物的難易程度既是影響A、B、C 子群選擇不同攝食策略的關(guān)鍵因素,也是導(dǎo)致表2 所示它們在生物學(xué)測定性狀表露“馬太效應(yīng)”的主要原因。故, 可將PC1歸為與機(jī)體代謝、魚體平衡、食物感知與定位、體質(zhì)與營養(yǎng)狀況以及運動方式相關(guān)的競食能力因子; (2) 周應(yīng)祺(1985)研究發(fā)現(xiàn), 鮐魚和鱈魚等紡錘形體型魚類游泳運動時頭部的擺幅最小, 之后波幅沿著魚體從頭至尾呈指數(shù)級增大。也就是說, 魚類的體型框架對推進(jìn)其游泳運動具重要作用, 尤以頭部之后的體型為甚。由此, PC2主要影響變量中除L9呈A≈B≈C (P>0.05)外, 余下的L11、L14、L15和L17均呈A≈B>C, 以及這些主要影響變量所涵蓋的體寬、背鰭部體高、體高、體長和尾柄高等體型框架關(guān)鍵信息均呈A>B>C (P<0.05)的結(jié)果(表2, 表3, 表4), 在進(jìn)一步印證了浙江中、北部海域大黃魚幼魚養(yǎng)殖群體尚處于以長長為主的生長階段, 與其體質(zhì)量關(guān)系最密切的體尺性狀為體長和體高(許益銨, 2014; 江柳等, 2021)的可靠性的同時, 也揭示了這些體型框架變量所隱含的暴發(fā)運動能力是導(dǎo)致大黃魚幼魚同生群內(nèi)出現(xiàn)增重性能差異的主要原因。故, 可將PC2歸為與暴發(fā)運動能力相關(guān)的體型框架比例因子; (3) 大黃魚的視覺、嗅覺器官和鰓均位于頭部。研究發(fā)現(xiàn), 視覺在魚類運動過程中具補(bǔ)償運動以保持視野穩(wěn)定的作用(Collinet al, 2003)。故,眼徑呈 A>B>C (P<0.05) (表2), 和L4呈 A≈B≈C(P>0.05)所示眼徑與肛長等速生長的結(jié)果(表3), 表明尋食視野在促進(jìn)養(yǎng)殖大黃魚幼魚的生長上具重要作用。根據(jù)“器官代謝活性假說”(Itazawaet al, 1983;Oikawaet al, 1992)可知, 表3 中C3(鰓質(zhì)量/體質(zhì)量)和C5(鰓質(zhì)量/凈體質(zhì)量)均呈A≈B 綜上, 上述公共因子所涵蓋的競食能力、暴發(fā)運動能力、食物搜尋能力等一系列與增重性能密切相關(guān)的生物學(xué)功能信息, 既真實反映了暴發(fā)運動能力和食物搜尋能力對競食能力的支撐作用, 以及競食能力對暴發(fā)運動能力和食物搜尋能力的牽引作用, 更揭示了競食能力、暴發(fā)運動能力和食物搜尋能力的三維互動是導(dǎo)致A、B、C 子群增重性能產(chǎn)生顯著異的內(nèi)在本質(zhì)。由此, 建議在大黃魚網(wǎng)箱養(yǎng)殖具體生產(chǎn)實踐中, 一方面可通過適當(dāng)增加投喂頻次并延長投喂時長, 以切實防止個體表型差異過大的現(xiàn)象, 另一方面應(yīng)及時實施分箱養(yǎng)殖, 通過有效緩減種內(nèi)食物競爭烈度, 使生長嚴(yán)重受抑個體通過競爭釋放(competion release)有效釋放生長潛能。 (1) 從生物學(xué)測定性狀看, 大黃魚幼魚在增重性能上具較大的定向選育價值和潛力; 從形質(zhì)評價指標(biāo)看, 絕大多數(shù)個體在形質(zhì)特征上有向高增重性能等級子群趨同的傾向, 其中唯一可清晰分辨A、B、C子群的形質(zhì)變量為重長系數(shù)K。 (2) 大黃魚幼魚在網(wǎng)箱養(yǎng)殖生境下貫徹并實施了以優(yōu)先保障攝食運動代謝為核心的生存對策。其中,A 子群依托強(qiáng)勁的競食優(yōu)勢, 建立了優(yōu)先獲得食物保障的高收益攝食策略, C 子群為有效彌補(bǔ)在競食中所處的劣勢地位, 固化了通過顯著提高機(jī)體代謝水平以大幅增加覓食運動量的低收益攝食策略, 而B 子群則確立了通過積極爭搶以努力提高食物保障程度的攝食策略。 (3) 網(wǎng)箱養(yǎng)殖生境下, 持續(xù)運動和暴發(fā)運動在大黃魚幼魚運動量上占比的多寡是導(dǎo)致A、B、C 子群增重性能差異的重要原因。以判別貢獻(xiàn)率較大的重長系數(shù)K和L10(側(cè)線長/肛長)為自變量, 建立的Fisher分類函數(shù)方程組可較清晰區(qū)分A、B、C 子群實驗個體, 綜合判別準(zhǔn)確率達(dá)97.78%。 (4) 競食能力、暴發(fā)運動能力和食物搜尋能力的三維互動是導(dǎo)致網(wǎng)箱養(yǎng)殖生境下大黃魚幼魚同生群內(nèi)個體間增重性能產(chǎn)生顯著差異的內(nèi)在本質(zhì)。在具體網(wǎng)箱養(yǎng)殖生產(chǎn)實踐中, 一方面可通過適當(dāng)增加投喂頻次并延長投喂時長, 切實防止個體表型差異過大的現(xiàn)象, 另一方面應(yīng)及時實施分箱養(yǎng)殖, 通過有效緩減種內(nèi)食物競爭烈度, 使生長嚴(yán)重受抑個體通過競爭釋放有效釋放生長潛能。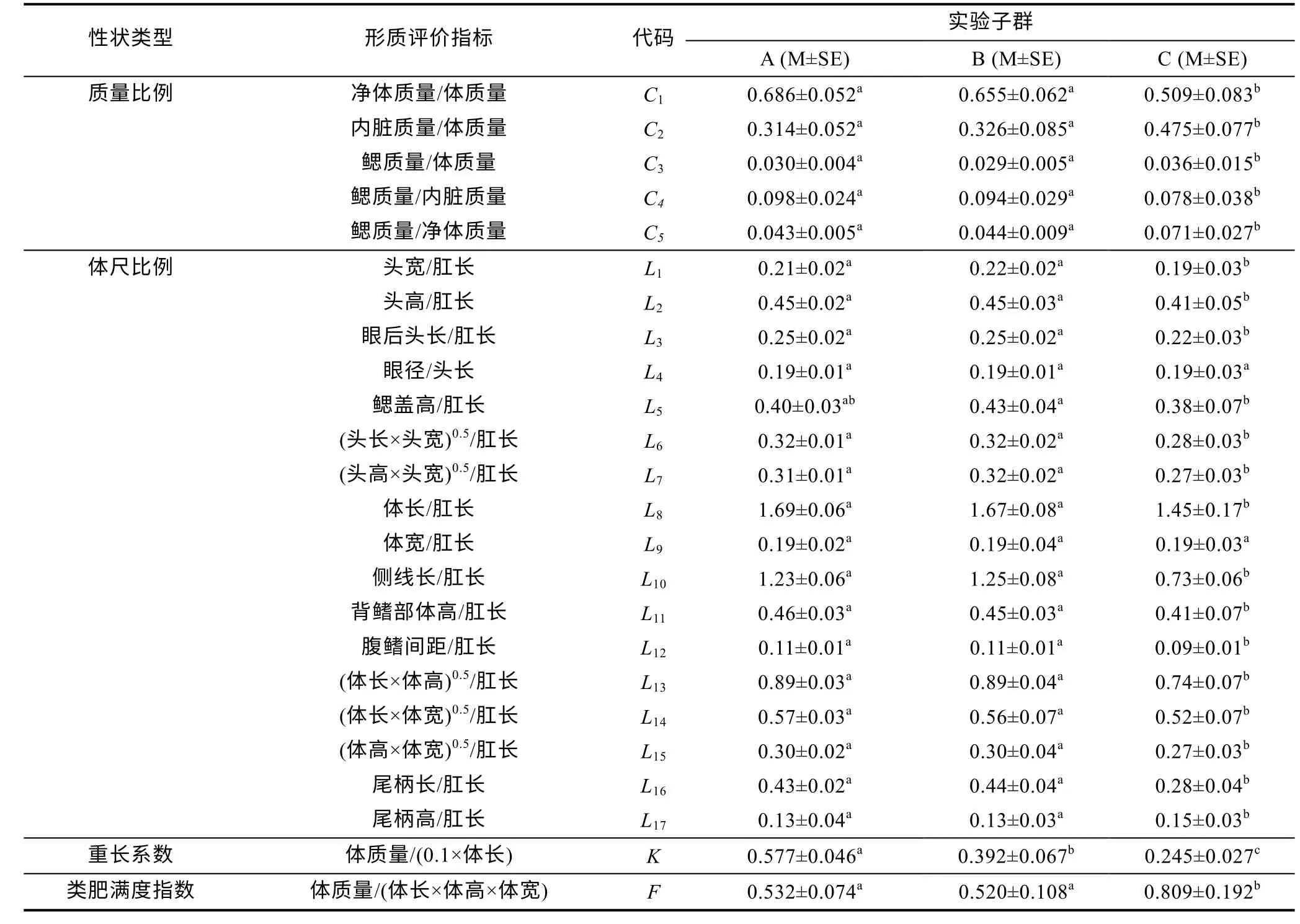
2.3 聚類分析
2.4 主成分分析
2.5 判別分析
3 討論
3.1 增重性能與攝食策略選擇間的相關(guān)性
3.2 增重性能與形質(zhì)特征間的相關(guān)性
4 結(jié)論
猜你喜歡
發(fā)明與創(chuàng)新(2022年30期)2022-10-03 08:40:56
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2022年6期)2022-06-05 06:50:58
意林(兒童繪本)(2020年2期)2021-01-07 02:12:04
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
作文成功之路·小學(xué)版(2020年5期)2020-06-11 12:48:46
意林(兒童繪本)(2019年9期)2019-10-15 08:51:46
中國生殖健康(2019年10期)2019-01-07 01:21:14
人大建設(shè)(2018年6期)2018-08-16 07:23:10
新高考(英語進(jìn)階)(2018年1期)2018-04-18 14:00:11
文理導(dǎo)航·科普童話(2017年5期)2018-02-10 19:42:14

- 海洋與湖沼的其它文章
- 海洋資料浮標(biāo)姿態(tài)信息測量技術(shù)研究現(xiàn)狀及發(fā)展趨勢*
- 瑞氏紅魴(Satyrichthys rieffeli)基因組survey分析及線粒體基因組注釋*
- 不同抗流能力大黃魚(Larimichthys crocea)肌肉轉(zhuǎn)錄組學(xué)差異分析*
- 美洲鰻鱺(Anguilla rostrata)養(yǎng)殖尾水高效處理與水循環(huán)養(yǎng)殖技術(shù)研究與應(yīng)用*
- 溫度和鹽度對虹鱒(Oncorhynchus mykiss)“水科1號”生長、消化酶活性和相關(guān)基因表達(dá)的影響*
- 深海化能極端環(huán)境中勞盆擬刺鎧蝦(Munidopsis lauensis)的全長轉(zhuǎn)錄組測序分析*