溫度和鹽度對虹鱒(Oncorhynchus mykiss)“水科1號”生長、消化酶活性和相關基因表達的影響*
2023-10-17 07:12:36黃天晴劉恩慧王高超王炳謙郭福元曹學彬鄭龍華董福霖徐革鋒
海洋與湖沼 2023年5期
關鍵詞:影響
紀 凱 黃天晴 谷 偉 劉恩慧 王高超 王炳謙郭福元 曹學彬 鄭龍華 董福霖 徐革鋒①
(1.農業農村部淡水水產生物技術與遺傳育種重點實驗室 中國水產科學研究院黑龍江水產研究所 黑龍江哈爾濱 150070;2.上海海洋大學水產與生命學院 上海 201306; 3.煙臺經海海洋漁業有限公司 山東煙臺 264006)
鮭鱒是世界第三大水產養殖品種, 據聯合國糧農組織統計, 2019 年全球產量超過350 萬噸(FAO,2021)。中國漁業統計年鑒資料顯示, 2021 年我國鮭鱒總產量不足4 萬t, 同時相較于2020 年總產量約減少2.89%。鮭鱒因其肉質鮮嫩可口而深受國內外的消費者喜愛, 對其的需求量也因此逐年增加, 而我國虹鱒產量遠不能滿足國內市場的需求, 仍需從國外大量進口(戶國等, 2017)。我國淡水養殖的鮭鱒品種主要以虹鱒(Oncorhynchus mykiss)為主, 虹鱒屬于鮭形目鮭科的一種廣鹽性魚類, 是我國廣泛養殖的冷水魚品種, 適宜生長的溫度為 12~18 °C (孫大江等,2010)。受到我國淡水資源日趨緊張及虹鱒生活習性的制約, 海水養殖虹鱒成為擴大我國鮭鱒產量, 解決市場供不應求的一種養殖方式。我國水產科技工作人員自20 世紀70 年代起在大連、煙臺、青島等地的開放海域嘗試養殖鮭魚, 但因夏季海水溫度過高不適合鮭鱒生存而失敗(董雙林, 2019)。然而, 在黃海中央洼地存在一個巨大的季節性冷水團, 為鮭鱒魚在海區開展養殖創造了條件(張竹琦, 1990; Yuet al, 2006;Dong, 2015; Xinet al, 2015; Hanet al, 2016)。
鰓和腸作為魚類滲透壓調節的重要器官, 能夠相互協調發揮作用, 兩組織均通過離子交換實現機體滲透壓調節(Giffard-Menaet al, 2006; Conet al,2017)。鰓和腸道中特定的離子轉運細胞通過多種離子通道和轉運蛋白來維持滲透壓平衡, 如nka和aqp3等。Nka通過為魚類體內的許多離子轉運系統提供驅動力, 在維持魚類體內滲透壓平衡的過程中發揮著重要作用(Kültz, 2015)。Aqp3是重要的水通道, 參與調節細胞的水通量(Cutleret al, 2007)。環境鹽度的變化通過影響nka和aqp3的基因表達進而對魚類的滲透壓平衡產生影響。鹽度和溫度的協同作用會對魚類的滲透壓平衡及消化吸收產生負面作用, 引起應激現象發生。熱休克蛋白是機體在應激情況下細胞內迅速合成的一組蛋白質, 在應對外界環境變化的過程中發揮著重要的作用。據報道,hsp70在水生動物適應溫度和鹽度變化的過程中發揮著作用(Yanget al,1992)。同時, 腸道滲透壓調節功能還與消化和吸收密切相關, 因為上皮組織吸收營養由離子梯度和膜轉運蛋白驅動(Bakkeet al, 2010), 腸道滲透壓的改變通過影響魚類的消化和吸收進而影響魚類的生長和發育。gh-igf-1 軸是魚類生長發育的重要調控軸, 外界鹽度的變化可通過擾亂gh和igf-1等因子的表達, 從而影響魚類的生長發育(李文笙等, 2010; 鄭艷等,2012; 岳蒙蒙, 2017)。
虹鱒具有很強的滲透壓調節能力, 經過科學合理的鹽度馴化可以在海水中存活, 并表現出良好的生長性能(Thorarensenet al, 1996; 劉騁躍等, 2018)。早期研究主要集中于不同鹽度條件下虹鱒的馴化方式(付占斐等, 2020)、消化酶活性和氧化應激能力(楊靜雯等, 2019)對虹鱒的影響, 而關于溫度和鹽度交互作用對虹鱒生長、滲透壓調節、消化酶活性及應激影響的研究尚未見報道。本研究以虹鱒“水科1 號”為研究對象, 通過在3 種溫度(10、16 和22 °C)條件下均設置5 種鹽度梯度處理(鹽度分別為0、8、16、24、32), 進行為期21 d 的養殖試驗, 比較探究溫度和鹽度交互作用對虹鱒生長、消化酶活性和生長、應激和滲透壓調節相關基因表達的影響, 以期為虹鱒的海水馴化和海水養殖提供科學依據。
1 材料與方法
1.1 實驗魚來源及暫養
虹鱒“水科1 號”來自中國水產科學研究院黑龍江水產研究所渤海冷水性魚類實驗站。在循環控溫水族箱(180 cm×60 cm×50 cm)中暫養14 d 后進行實驗, 暫養期間的光暗比為(12L : 12D), 水溫控制在(16±0.5) °C。暫養期間保持連續供氧, 實測溶解氧濃度為 7.8~10.0 mg/L, pH 7.1~7.5。暫養期間每日按照魚體重的2%投喂配合飼料(北京漢業科技有限公司), 實驗前一天停止投餌。挑選健康個體作為實驗對象備用, 規格為初始體長(14.16±1.03) cm、初始體重(40.00± 2.36) g。
1.2 實驗設計
實驗共設3 種溫度梯度(10、16 和22 °C), 同時每種溫度下設置5 個鹽度梯度處理(鹽度分別為0、8、16、24、32)。10 °C 條件下鹽度梯度分別以A1~A5表示; 16 °C 條件下分別以B1~B5表示; 22 °C條件下分別以C1~C5表示。以16 °C 且鹽度0 組作為基礎, 采用日升(降)溫度1 °C 的同時日升鹽度2 的方法, 直至達到目的溫度和鹽度, 暫養1 d 后開展為期21 d 的養殖試驗, 實驗設計見表1。每種鹽度處理設置三組平行實驗組, 每個平行組放置8 尾魚。鹽度采用曝氣自來水與海水素(海之絢海洋生物有限公司, 青島, 中國)調配, 鹽度變化通過手持鹽度計(AZ-8371)測定。溫度通過循環控溫水族箱控制溫度, 溫差控制在±0.2 °C。實驗期間管理與暫養期間一致, 每日投喂商業飼料兩次至視覺飽足并做好記錄。實驗期間及時撈出死魚及糞便等。
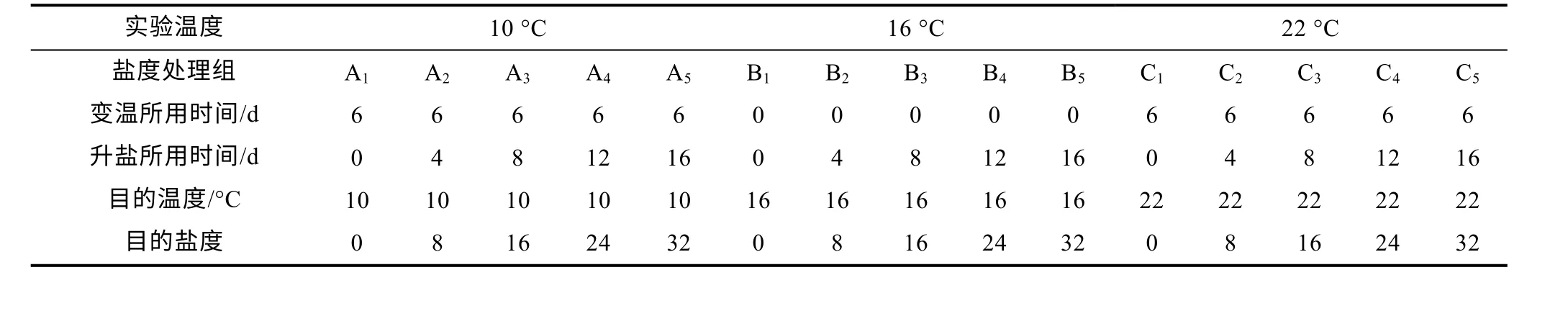
表1 實驗設計表Tab.1 The experimental design
1.3 樣品采集
生長實驗結束后停食1 d, 將實驗魚經MS-222麻醉后用電子天平(HC-C, 0.01 g)測量終末體重。隨后,分別從每種實驗條件下的三組平行組中各隨機選取6尾魚分別用于消化酶活性測定以及RNA 提取, 在無菌冰盤上解剖取得鰓和腸道組織(中腸), 剔除內容物和脂肪, 經液氮迅速冷凍后于–80 °C 超低溫冰箱中保存, 用于后續實驗測定。
1.4 腸組織酶活性測定
淀粉酶(AMS)活性、脂肪酶(LPS)活性及酶液蛋白含量測定均采用南京建成生物工程研究所生產的試劑盒進行測定。相關酶活性及蛋白含量的測定均按照相應的試劑盒說明書進行。
1.5 相關基因表達分析
將鰓和腸組織解凍后, 使用Simply P 總RNA 提取試劑盒(BioFlux, 杭州, 中國)對RNA 進行提取。通過分光光度計在260 和280 nm 處測定吸光度值來檢測所提取RNA 的純度, 1%瓊脂糖凝膠電泳評估RNA的完整性。使用PrimeScript RT 試劑盒(TaKaRa, 大連,中國)合成cDNA。采用Roche 公司(瑞士) FastStart Universal SYBR?Green Master Mix (ROX)進行實時定量PCR。qPCR 反應體系為10.0 μL, PCR 反應條件為: 95 °C 10 min, 95 °C 下變性15 s, 40 個循環60 °C下退火30 s。以β-actin作為內參基因。使用CFX96 C1000 TOUCH 熒光定量儀(BIO-RAD, 美國)檢測Ct值, 通過2–ΔΔCt方法計算基因的相對表達水平, 每個樣本重復三次。實驗中所用到的引物序列見表2。
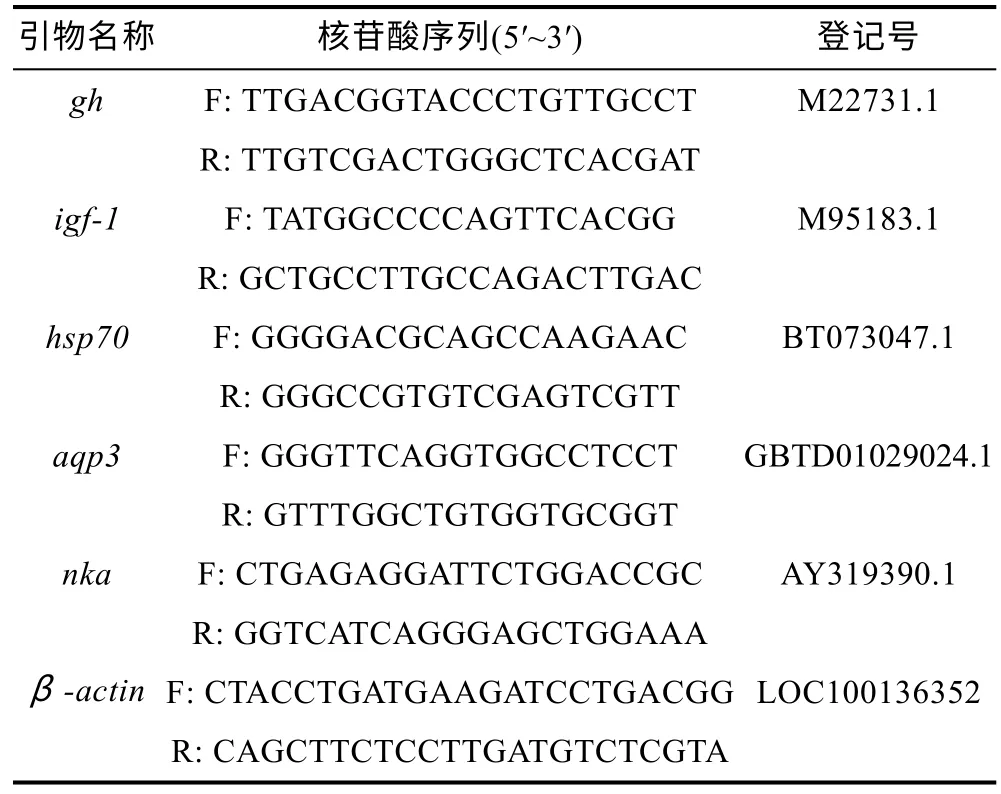
表2 PCR 引物的核苷酸序列Tab.2 The nucleotide sequences of PCR primers
1.6 計算公式與統計分析
餌料轉化率FCE (feed conversion efficiency)、增重率 WGR (weight gain rate)和特定生長率 SGR(specific growth rate)的計算公式如下:
式中,W0和Wt分別表示實驗魚初始體重和終末體重(g);Cw表示實驗魚在實驗期間總攝食量(g);t表示實驗時間(d)。
實驗數據均使用SPSS 25.0 版(SPSS Inc, Chicago,USA)進行統計分析, 以溫度和鹽度作為自變量, 進行雙因素方差分析(Two-Way ANOVA), 并采用Duncan 多重比較法檢驗不同實驗組平均值間是否存在差異(P<0.05)。統計結果均以平均值±標準差(Mean±SD)表示, 以P<0.05 作為差異顯著水平。使用GraphPad Prism 5.0 (San Diego, CA, USA) 可視化所有統計數據。
2 結果
2.1 不同溫度和鹽度對虹鱒“水科1 號”生長性能的影響
溫度和鹽度交互處理后虹鱒的FCE、WGR 和SGR 及存活率見表3。從表3 中可以看出, 10 °C 時, 隨著鹽度增大FCE、WGR 和SGR 三項指標均呈下降趨勢; 除A1組FCE 與B1(淡水對照組)無顯著差異外,其余鹽度組FCE、WGR、SGR 三項指標均低于B1(淡水對照組)。16 °C 時, 隨著鹽度增大FCE、WGR 和SGR 均呈先升高后下降的趨勢; 除B2組FCE、WGR和SGR 均高于B1(淡水對照組), 其余實驗組三項指標均較B1(淡水對照組)下降。22 °C 時, 隨著鹽度增大FCE、WGR 和SGR 三項指標均呈下降趨勢, 均顯著低于B1(淡水對照組)。此外, 養殖實驗過程中, 三種不同溫度條件下均在32 鹽度組中出現實驗魚死亡現象, 其中16 °C 時三組平行組共死亡1 尾魚, 10 °C時三組平行組共死亡3 尾魚, 22 °C 時三組平行組共死亡4 尾魚。不同溫度和鹽度處理后虹鱒的FCE、WGR 和SGR 雙因素方差分析結果如表4 所示。從表4 中可得出, 虹鱒的FCE、WGR 和SGR 均受到溫度和鹽度的交互影響顯著(P<0.05)。
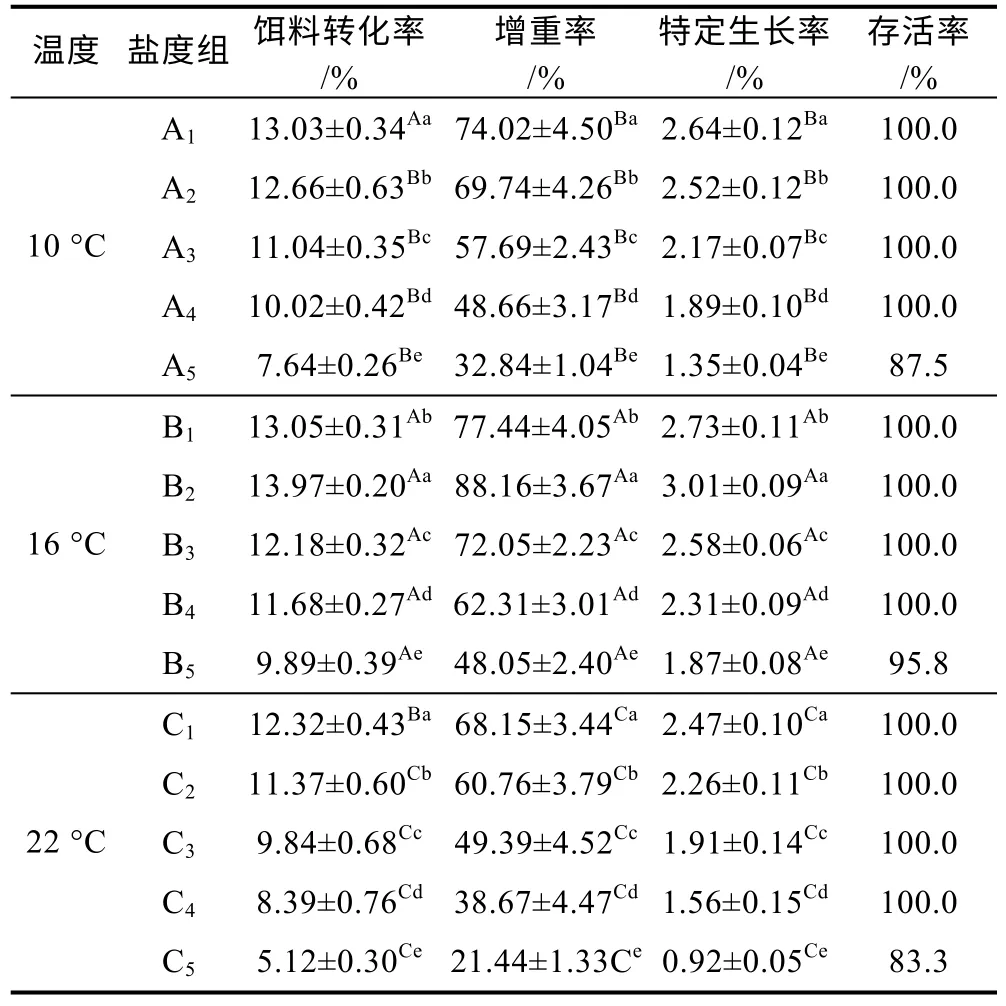
表3 溫度和鹽度交互作用對虹鱒生長性能的影響Tab.3 Effect of temperature-salinity interaction on growth performance of rainbow trout
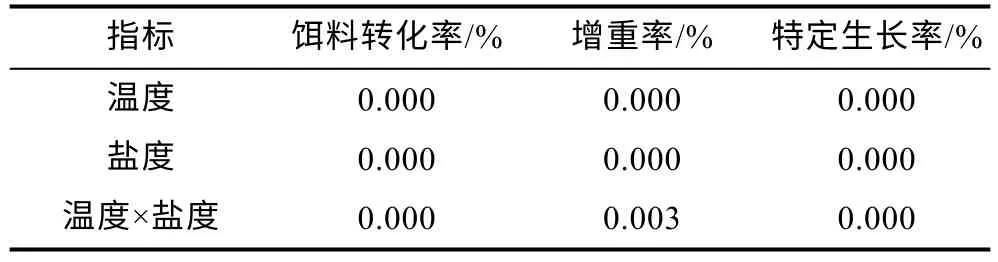
表4 溫度和鹽度及其交互作用對虹鱒生長性能影響的雙因素方差分析P 值Tab.4 Two-factor ANOVA P values for the effects of temperature, salinity, and their interaction on growth performance of rainbow trout
2.2 溫度和鹽度對虹鱒“水科1 號”腸組織中消化酶活性的影響
溫度和鹽度交互作用下虹鱒腸道淀粉酶活性和脂肪酶活性變化見圖1。由圖1 可知, 10 °C 時, 腸道中淀粉酶和脂肪酶活性均隨著鹽度增大呈降低趨勢,各鹽度組淀粉酶活性分別較對照組活性顯著降低約15%、16%、29%、53%和75% (P<0.05); 脂肪酶活性分別較對照組顯著下降約12%、21%、30%、39%和46% (P<0.05)。16 °C 時, 淀粉酶和脂肪酶活性隨著鹽度增大均呈先升高后下降的趨勢, 淀粉酶活性除B2組較對照組顯著升高約23%, 其余鹽度組淀粉酶活性分別較對照組顯著下降約25%、45%和49%; 脂肪酶活性除B2組較對照組顯著升高約8%, 其余鹽度組脂肪酶活性分別較對照組顯著下降約12%、24%和31%。22 °C 時, 腸道中淀粉酶活性和脂肪酶活性隨著鹽度的增大呈下降趨勢, 各鹽度組淀粉酶活性分別較對照組顯著下降約18%、32%、42%、51%和63%;脂肪酶活性分別較對照組顯著下降約10%、16%、19%、34%和47%。此外, 在同一鹽度條件下, 腸道中淀粉酶和脂肪酶活性均隨著溫度的升高呈先升高后下降的趨勢, 16 °C 實驗組顯著高于10 和22 °C 實驗組(P<0.05)。溫度和鹽度交互處理后虹鱒腸道中淀粉酶活性和脂肪酶活性雙因素方差分析結果如表5所示。從表5 中可以看出, 虹鱒腸道中淀粉酶活性和脂肪酶活性均受到溫度和鹽度的交互影響顯著(P<0.05)。
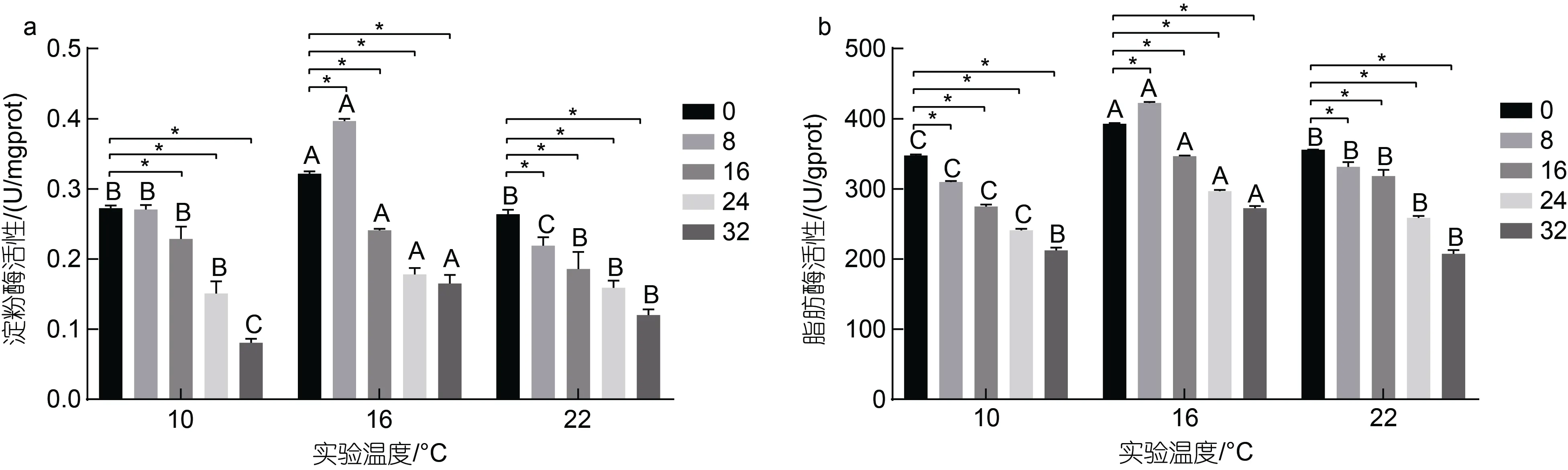
圖1 溫度和鹽度交互作用對虹鱒腸道消化酶活性的影響Fig.1 Effect of temperature- salinity interaction on intestinal digestive enzyme activity of rainbow trout
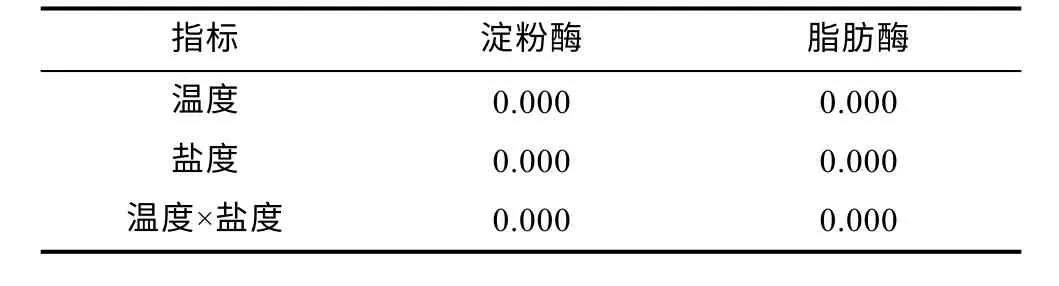
表5 溫度和鹽度及其交互作用對虹鱒腸道消化酶影響的雙因素方差分析P 值Tab.5 Two-factor ANOVA P values for the effects of temperature, salinity, and their interaction on the intestinal digestive enzyme activities of rainbow trout
2.3 溫度和鹽度交互作用對虹鱒“水科1 號”鰓和腸組織中基因表達的影響
鰓組織中gh表達量如圖2a, 結果表明, 10 °C 時,gh表達量隨著鹽度增大呈下調趨勢, 各實驗組分別較對照組顯著下調6%、10%、18%、22%和31%(P<0.05); 16 °C 時,gh表達量隨實驗組鹽度增大呈先上調后下調的趨勢, B2組和B3組表達量分別較對照組顯著上調約79%和47%, B4組較對照組無顯著差異(P>0.05), B5組較對照組顯著下調16% (P<0.05);22 °C 時,gh表達量隨著鹽度增大呈下調趨勢, 各實驗組分別較對照組顯著下調5%、7%、13%、19%和28% (P<0.05)。鰓組織中igf-1表達量如圖2b 所示, 從圖中可以觀察到,igf-1表達量的變化趨勢基本與gh一致, 10 °C 時,igf-1表達量隨著鹽度增大呈下調趨勢,各實驗組分別較對照組顯著下調1%、1%、8%、29%和49% (P<0.05); 16 °C 時,igf-1表達量隨實驗組鹽度增大呈先上調后下調的趨勢, B2組和B3組表達量分別較對照組顯著上調約48%和21%, B4組較對照組無顯著差異(P>0.05), B5組較對照組顯著下調 16%(P<0.05); 22 °C 時,igf-1表達量隨著鹽度增大呈下調趨勢, 各實驗組分別較對照組顯著下調1%、3%、7%、19%和27% (P<0.05)。鰓組織中hsp70表達量如圖2c所示, 各溫度下hsp70表達量隨鹽度增大呈上調趨勢,10 °C 時, 各鹽度組hsp70分別較對照組顯著上調約25%、98%、132%、218%和494% (P<0.05); 16 °C 時,除B2組和B3組較對照組無顯著差異外(P>0.05), 其余鹽度組分別較對照組顯著上調約 73%和 129%;22 °C 時, 各鹽度組分別較對照組顯著上調約68%、128%、118%、246%和301% (P<0.05)。鰓組織中aqp3表達量如圖2d 所示, 三種不同溫度下aqp3表達量隨鹽度增大均呈下調趨勢, 10 °C 時, 除A1組與對照組無顯著差異外(P>0.05), 其余鹽度組分別較對照組顯著下調9%、17%、25%和40% (P<0.05); 16 °C 時, 除B2組較對照組無顯著差異外(P>0.05), 其余鹽度組分別較對照組顯著下調10%、22%和33% (P<0.05);22 °C 時, 除C1 組較對照組無顯著差異外(P>0.05),其余鹽度組分別較對照組顯著下調6%、15%、25%和36% (P<0.05)。鰓組織中nka表達量如圖2e 所示,三種不同的溫度下nka表達量隨鹽度增大均呈上調趨勢, 10 °C 時, 除A1組與對照組無顯著差異外(P>0.05), 其余鹽度組分別較對照組顯著上調19%、98%、210%和390% (P<0.05); 16 °C 時, 除B2組較對照組無顯著差異外(P>0.05), 其余鹽度組分別較對照組顯著上調56%、155%和310% (P<0.05); 22 °C 時, 除C1組較對照組無顯著差異外(P>0.05), 其余鹽度組分別較對照組顯著上調9%、83%、198%和365% (P<0.05)。此外, 在鹽度相同溫度不同時, 16 °C 實驗組較10 °C和22 °C 實驗組gh和igf-1表達量顯著上調(P<0.05)、hsp70表達量顯著下調(P<0.05)、aqp3表達量(除16和32 鹽度時)無顯著差異(P>0.05)、nka表達量(除0鹽度時)顯著下調(P<0.05)。不同溫度和鹽度處理后虹鱒鰓組織中gh、igf-1、hsp70、aqp3和nka基因表達雙因素方差分析結果如表6 所示。從表6 中可以看出,虹鱒鰓組織中gh、igf-1、hsp70、aqp3和nka基因表達均受到溫度和鹽度的交互影響顯著(P<0.05)。
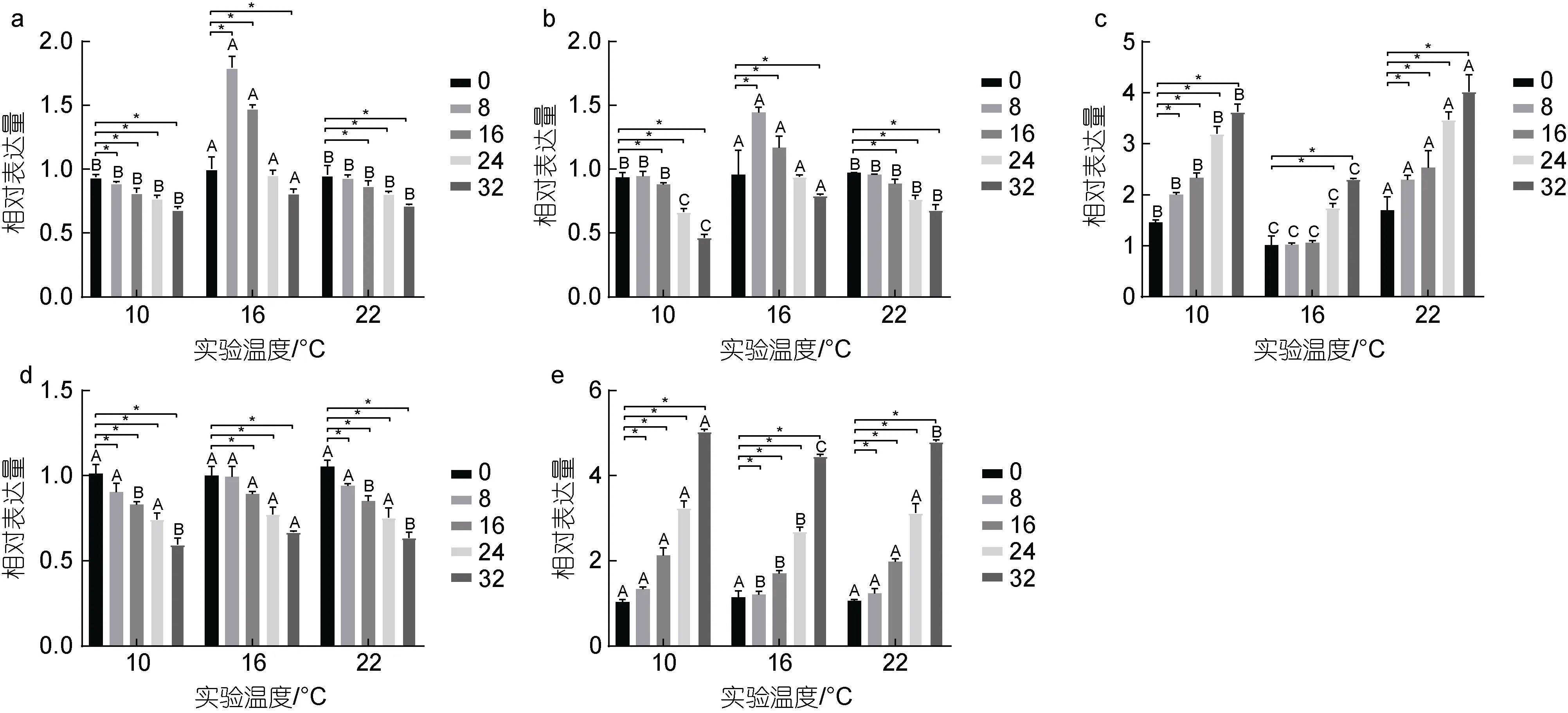
圖2 溫度和鹽度交互作用對虹鱒鰓組織中gh、igf-1、hsp70、aqp3 和nka 基因表達的影響Fig.2 Effect of temperature-salinity interaction on gh, igf-1, hsp70, aqp3, and nka gene expression in gill tissue of rainbow trout
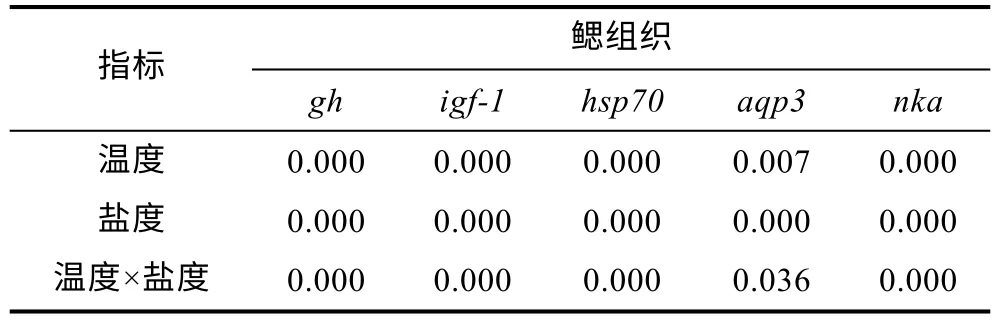
表6 溫度和鹽度及其交互作用對虹鱒鰓組織中gh、igf-1、hsp70、aqp3 和nka 影響的雙因素分析P 值Tab.6 Two-way ANOVA P values for the effects of temperature, salinity and their interaction on gh, igf-1, hsp70,aqp3, and nka in gill tissue of rainbow trout
腸組織中gh表達量如圖3a, 10 °C 時,gh表達量隨鹽度增大呈先上調后下降的趨勢, 除A2組較對照組無顯著差異外(P>0.05), 其余鹽度組分別較對照組顯著下調8%、14%、19%和25% (P<0.05); 16 °C 時gh表達量隨鹽度增大呈先上調后下調的趨勢, B2和B3組分別較對照組顯著上調77%和16% (P<0.05), 其余鹽度組分別較對照組顯著下調2%和11% (P<0.05);22 °C 時,gh表達量隨鹽度增大整體呈下降趨勢, 各鹽度組分別較對照組顯著下調10%、10%、19%、24%和31% (P<0.05)。腸組織中IGF表達量如圖3b, 10 °C時,igf-1表達量隨鹽度增大呈先上調后下調的趨勢,除A2組較對照組無顯著差異外(P>0.05), 其余鹽度組分別較對照組顯著下調 7%、11%、18%和 24%(P<0.05); 16 °C 時,igf-1表達量隨鹽度增大呈先上調后下調的趨勢, B2和B3組分別較對照組顯著上調92%和7% (P<0.05), 其余鹽度組分別較對照組顯著下調7%和13% (P<0.05); 22 °C 時,igf-1表達力隨鹽度增大呈下調的趨勢; 各實驗組分別均較對照組顯著下調9%、6%、19%、23%和30%(P<0.05)。腸組織中hsp70表達量如圖3c, 結果顯示, 不同溫度條件下hsp70表達量隨鹽度增大均呈上調趨勢; 10 °C 時,各鹽度組hsp70分別較對照組顯著上調約31%、29%、97%、159%和205% (P<0.05); 16 °C 時, 除B2組較對照組無顯著差異外(P>0.05), 其余鹽度組分別較對照組顯著上調約34%、93%和124%; 22 °C 時, 各鹽度組分別較對照組顯著上調約76%、71%、180%、217%和285% (P<0.05)。腸組織中aqp3表達量如圖3d, 不同溫度條件下aqp3表達量均隨鹽度增大呈上調趨勢,10 °C 時, 隨著鹽度組鹽度增大, 各鹽度組分別較對照組顯著下調 4%、45%、304%、316%和 497%(P<0.05); 16 °C 時, 隨著鹽度組鹽度增大, 各鹽度組分別較對照組顯著下調36%、176%、273%和418%(P<0.05); 22 °C 時, 隨著鹽度組鹽度增大, 各鹽度組分別較對照組顯著下調5%、60%、257%、291%和778% (P<0.05)。腸組織中nka表達量如圖3e, 三種不同的溫度下, 各實驗組nka表達量隨鹽度增大均呈上調趨勢, 10 °C 時, 隨著鹽度組鹽度增大, 各鹽度組分別較對照組顯著上調6%、135%、469%、595%和606%(P<0.05); 16 °C 時, 隨著鹽度組鹽度增大, 各鹽度組分別較對照組顯著上調61%、354%、597%和719%(P<0.05); 22 °C 時, 隨著鹽度組鹽度增大, 各鹽度組分別較對照組顯著上調18%、84%、350%、560%和700% (P<0.05)。此外, 在鹽度相同溫度不同時, 16 °C實驗組較10 °C 和22 °C 實驗組gh和igf-1表達量顯著上調(P<0.05)、hsp70表達量顯著下調(P<0.05)。溫度和鹽度交互處理后虹鱒鰓組織中gh、igf-1、hsp70、aqp3和nka基因表達雙因素方差分析結果如表7 所示。從表7 中可以看出, 虹鱒腸組織中gh、igf-1、hsp70、aqp3和nka基因表達均受到溫度和鹽度的交互影響顯著(P<0.05)。
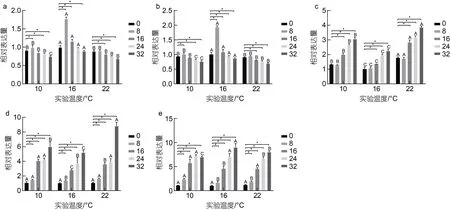
圖3 不同溫度和鹽度對虹鱒腸組織中gh、igf-1、hsp70、aqp3 和nka 基因表達的影響Fig.3 Effects of temperature and salinity on gh, igf-1, hsp70, aqp3,and nka gene expression in intestinal tissue of rainbow trout
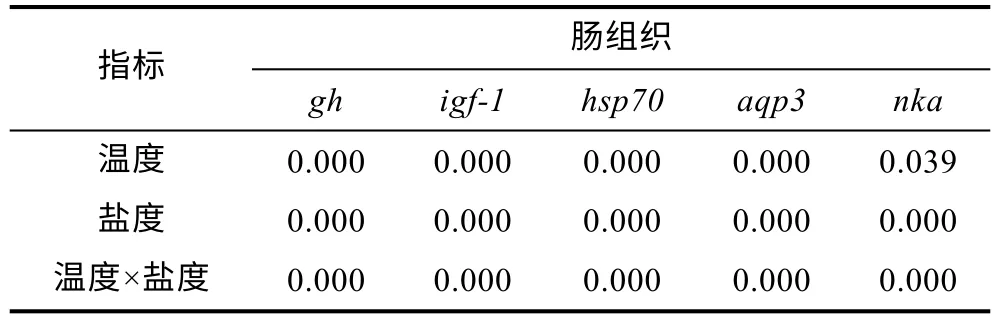
表7 溫度和鹽度及其交互作用對虹鱒腸組織中gh、igf-1、hsp70、aqp3 和nka 影響的雙因素方差分析P 值Tab.7 Two-way ANOVA P values of the effects of temperature,salinity, and their interactions on gh, igf-1, hsp70, aqp3, and nka in rainbow trout intestinal tissues
3 討論
在溫度和鹽度馴化過程的中, 不同的升(降)溫和升鹽方式會影響魚類的生長發育及存活率。有研究表明, 通過較慢的升溫速率更能真實地反映自然環境中魚類受到影響(Galbreathet al, 2004; 姜旭陽等,2019)。因此本實驗選用日升溫度或日降溫度1 °C 的方式模擬養殖過程中面臨的溫度變化。有關魚類生長發育及存活率受到鹽度馴化方式的影響已有不少報道, 鹽度馴化過程中升鹽速率過大會影響魚類的生長發育。黃國強等(2013)對褐牙鲆(Paralichthy solivaceus)鹽度馴化的過程中發現, 采用日升鹽度7的方式時, 褐牙鲆血清滲透壓需要2 d 才能恢復穩定;采用日升鹽度14 的方式血清滲透壓則需要4 d 才能回復穩定。付占斐等(2020)在研究不同鹽度馴化方式對虹鱒和硬頭鱒的影響中發現, 隨著日升鹽度的增大存活率隨之下降, 作者指出, 采用緩慢的升鹽方式即日升鹽度2 進行鹽度馴化更有利于虹鱒和硬頭鱒的生長發育。因此, 本實驗中采用日升鹽度2 的方式達到實驗目的鹽度, 在養殖試驗過程中, 三種不同溫度條件均在32 鹽度組中出現虹鱒死亡現象, 可能由于實驗魚個體間存在差異, 無法耐受當前的實驗條件。溫度和鹽度是影響水產動物生長和存活的重要環境因子, 合適的溫度范圍以及適當的鹽度環境可促進水產動物的生長發育, 反之則會產生不利影響(Buckelet al, 1995; 金方彭等, 2018; 王潤萍等, 2019;李培倫等, 2020b)。據報道, 硬骨動物血液中滲透壓約為12, 約為海水鹽度(38)的三分之一(Mylonaset al,2009)。大量研究表明, 中低鹽度下會促進廣鹽性魚類的生長, 因為此時的鹽度更接近魚類的等滲點, 降低了對滲透壓調節的需求, 可為魚類生長提供更多的能量(Martinez-Cardenaet al, 2014; Godaet al, 2019;Haideret al, 2021)。本研究結果表明, 溫度和鹽度交互影響對虹鱒的FCE、WGR 和SGR 影響顯著(P<0.05)。在16 °C 時, 鹽度為8 實驗組虹鱒的FCE、WGR 和SGR 較B1淡水對照組顯著增加, 即適當的升高鹽度提高了虹鱒的生長性能。這在不同的廣鹽魚類中也有報道, 如歐洲鱸(Dicentrarchus labrax) (Bernardinoet al,2016)、黃鰭鯛(Acanthopagrus latus)和亞洲鱸(Lates calcarifer) (Mozanzadehet al, 2021)、大西洋鱈魚(Gadus morhua) (árnasonet al, 2013)等。
魚類的消化酶活性通過影響機體對營養物質的消化和吸收進而影響魚類的生長發育, 而養殖水體中的溫度和鹽度是影響魚類消化酶活性的主要因素(歐志聯, 2018)。本研究結果顯示, 同一鹽度不同溫度條件下, 虹鱒腸道中淀粉酶和脂肪酶的活性隨著溫度增大呈先升高后下降的趨勢。劉鑒毅等(2015)在溫度對點籃子魚(Siganus guttatus)消化酶活性影響的研究中發現, 點籃子魚腸道中淀粉酶的活性隨著溫度的升高呈先升后降的趨勢。梅景良等(2004)在研究溫度和pH 對黑鯛(Sparus macrocephalus)腸等消化酶活性的影響中發現, 脂肪酶活性的變化趨勢隨著溫度的升高先上升達到峰值后下降。上述研究結果與本實驗中觀察到的結果相似。本研究中, 同一溫度不同鹽度條件下, 10 °C 和22 °C 虹鱒腸道中淀粉酶和脂肪活性整體隨著鹽度的升高而降低; 16 °C 時, 腸道中脂肪酶和淀粉酶活性隨鹽度增大先升高后下降。楊靜雯等(2021)在研究鹽度對虹鱒和硬頭鱒消化酶活性的研究中發現, 隨著鹽度的升高, 虹鱒腸道中脂肪酶活性表現為先升后降, 鹽度為5 時活性最高, 與本研究中溫度為16 °C、鹽度為8 實驗組的研究結果相近。諸多研究表明, 鹽度的變化對消化酶活性的影響有三種不同的情況, 分別為促進、抑制和無顯著影響(羅鳴鐘等, 2015; 劉永士等, 2020; 李培倫等, 2020a)。本研究中, 受到溫度和鹽度的雙重影響, 虹鱒腸道中消化酶的活性在不同實驗條件下表現出不同的變化趨勢。造成這種現象的原因有很多, 虹鱒的最適生長溫度為12~18 °C, 高出或低于此溫度范圍都會影響虹鱒正常的生長發育(夏斌鵬等, 2017; 劉騁躍等, 2019;姜旭陽等, 2021)。溫度的變化會影響魚類獲取食物的欲望, 以及在腸道中消化食物吸收營養和儲存多余能量的能力(Volkoffet al, 2020)。環境鹽度的變化可能會改變魚類的飲水量、腸道中內容物的pH 值以及離子濃度等, 進而影響魚類的消化酶活性(Liuet al,2017; Mozanzadehet al, 2021)。本研究結果表明, 在適宜的溫度條件下, 適當的增加鹽度會促進虹鱒的消化酶活性, 這與本研究中生長性能指標相印證。同時, 在美洲鯡(Alosa sapidissima) (成永洲, 2015)、斜帶石斑魚(Epinephelus coioides) (Sakamotoet al, 1993)等的研究中也證實了這一點。
生長激素(Growth Hormone,gh)和胰島素生長因子(Insulin-like Growth Factor,igf)除了參與魚類生長發育調節作用外, 還能夠通過參與滲透壓調節來促進水生生物鹽度適應的過程(劉紅云等, 2004; 董云偉等, 2008)。養殖水體溫度和鹽度的變化不僅會影響魚類的生長發育, 滲透壓調節等, 還會導致魚類產生異常的應激反應。在本研究中, 鰓和腸組織中gh和igf-1表達量在16 °C 時隨實驗組鹽度增大呈先升高后下降的趨勢, 鹽度為8 實驗組表達量較對照組顯著上調(P<0.05), 說明此鹽度下利于促進虹鱒的生長。Madsen 等(1992)在對褐鱒(Salmo trutta)和虹鱒海水適應的研究中指出, 低鹽度下gh表達量上調從而促進了魚體的生長發育。李明云等(2015)在研究低鹽脅迫對大黃魚(Pseudosciaena crocea)gh和igf-1基因表達變化的影響時指出, 低鹽環境下大黃魚鰓、肝和腸等組織中gh和igf-1的表達量升高。二者的研究結果與本研究相似。但在10 °C 和22 °C 時, 虹鱒鰓和腸組織中gh和igf-1表達量隨著鹽度的升高整體呈現下降趨勢。這可能是由于在溫度和鹽度的雙重因素影響下,虹鱒處于應激狀態而抑制了鰓和腸組織中gh和igf-1基因的表達。
熱休克蛋白70 (Heat Shock Protein,hsp70)又稱應激蛋白70, 是HSPs 家族中重要的一員, 當機體處于應激條件下(溫度、鹽度等)時會被誘導表達上調。有報道稱,hsp70參與水生動物溫度和鹽度適應的過程(Bakkeet al, 2010)。本研究結果顯示, 鰓和腸組織中hsp70的相對表達量在各溫度下均隨實驗組鹽度增大呈上調趨勢。在鹽度相同溫度不同時, 鰓和腸組織中hsp70表達量均表現為22 °C 時最高, 10 °C 次之,16 °C 最低。虹鱒鰓和腸組織中hsp70相對表達量不同程度的變化, 說明溫度和鹽度的交互影響造成虹鱒體內應激現象的產生。據報道稱, 溫度和鹽度的變化會導致細胞內蛋白質肽鏈失去原有的折疊結構, 使空間結構發生改變, 失去原有功能;hsp70會在外界因素刺激下大量表達, 修復或降解細胞內的變性蛋白質, 增加機體的抗應激能力, 從而緩解溫度和鹽度變化帶來的應激壓力(任寶波等, 2005; 辛苑茹等, 2019)。
養殖環境鹽度的改變會影響魚類滲透壓平衡,進而影響魚類的生長發育。鰓和腸作為魚類滲透壓調節的重要器官, 能夠相互協調發揮作用, 兩組織通過離子交換、吸收水分等方式實現機體滲透壓調節。在本研究中, 我們觀察到暴露于不同溫度和鹽度下虹鱒鰓和腸道中aqp3的表達存在差異。鰓組織中, 不同溫度下,aqp3表達量隨著實驗組鹽度增大呈下調趨勢。Tipsmark 等(2010)在對大西洋鮭(Salmo salar)海水馴化時滲透壓調節組織中水通道蛋白表達的研究中發現, 海水條件下會誘導大西洋鮭鰓組織中aqp3的表達下調。Cutler 等(2002)和Giffard-Mena 等(2007)分別在對歐洲鰻鱺(Anguilla anguilla)和歐洲鱸受鹽度影響的研究中指出, 海水馴化后廣鹽魚的鰓組織中aqp3表達量呈下調趨勢。上述研究與本實驗中觀察到的結果相似。有報道指出,aqp3在鰓組織中的作用可能是通過促進水流出鰓上皮到循環系統中(Cutleret al, 2000)。但這不利于處于高鹽環境中的魚類維持滲透壓的平衡, 這可能是造成鰓組織中aqp3表達量下調的原因。在本研究的腸組織中, 不同溫度下, 腸組織中aqp3的表達量隨實驗組鹽度增大呈上調趨勢。Choi 等(2013)在對紅大馬哈魚(Oncorhynchusnerka)海水馴化的研究中發現, 腸組織中aqp3表達量會增加, 與本實驗的研究結果相似。眾多的研究表明,aqp3存在于腸道的上皮細胞中, 通過吸收控制鹽分的流入來維持滲透壓的平衡(Martinezet al, 2005), 本研究腸組織aqp3表達量的變化表明其在維持滲透壓平衡中發揮著作用。在本研究中, 不同溫度下, 虹鱒鰓和腸組織中nka表達量均隨實驗組鹽度增大呈上調趨勢。這與Tang 等(2012)對經鹽度馴化的日本鰻鱺(Anguilla japonica)鰓組織中的nka和Chourasia 等(2018)對高鹽適應下羅非魚(Oreochromis mossambicus)腸道中nka的表達趨勢一致。nka對維持細胞內穩態至關重要, 通過進行離子轉運產生初始電化學梯度,為其他跨膜蛋白進行離子和水交換提供基礎, 在維持魚類滲透壓平衡中發揮著重要的作用(Zhanget al,2019; Saghafiankhoet al, 2020)。在虹鱒鰓和腸組織中觀察到aqp3和nka不同程度的表達水平, 表明aqp3和nka在到維持虹鱒滲透壓平衡的調節中發揮著重要的作用。
4 結論
綜上可得出結論, 本文研究了不同鹽度和溫度對虹鱒“水科1 號”生長指標、消化酶活性及相關基因表達的影響。結果發現, 16 °C 下, 鹽度為8 實驗組的虹鱒較淡水對照組生長表現更好, 即虹鱒在適宜生長的溫度下, 適當的提升養殖水體鹽度有利于促進虹鱒的生長。本研究中還發現, 與低鹽度組相比, 各溫度下高鹽度組整體的試驗周期雖然更長, 但從生長指標、消化酶活性及鰓和腸組織中基因表達的結果來看, 并沒有彌補鹽度升高帶來的負面影響, 盡管在最適的生長溫度下, 高鹽度仍抑制了虹鱒的生長發育。本研究為溫度和鹽度變化對虹鱒海水馴化和海水養殖的研究提供了基礎資料。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
