深海化能極端環境中勞盆擬刺鎧蝦(Munidopsis lauensis)的全長轉錄組測序分析*
2023-10-17 07:12:32沙忠利
海洋與湖沼 2023年5期
閆 晗 惠 敏 沙忠利① 程 嬌①
(1.中國科學院海洋研究所海洋生物分類與系統演化實驗室 青島市海洋生物多樣性與保護重點實驗室 山東青島 266071;2.嶗山實驗室 山東青島 266237; 3.中國科學院大學 北京 100049)
自1977 年美國深海潛水器阿爾文號在加拉帕戈斯裂谷附近發現深海熱液活動和高密度的熱液生物群落以來, 深海化能合成生態系統開始展現于世人面前(Corlisset al, 1979)。隨后, 1983 年美國研究人員在廣闊的墨西哥灣3 200 m 深處首次發現了海底冷泉系統(Andersonet al, 1983)。與依賴太陽光能的光合生態系統不同, 以熱液和冷泉為代表的深海化能合成生態系統主要依靠化能自養微生物利用硫化氫(H2S)、元素硫(S)、甲烷(CH4)和氫氣(H2)等還原性物質氧化產生的能量進行固碳、合成有機物, 從而維持生態系統的運轉, 這種“黑暗食物鏈”的發現改變了人們對深海初級生產力主要來源的認知(Dubilieret al, 2008)。此外, 深海化能生態系統是認識特殊生命過程和研究生物適應性進化的天然實驗場所。深海化能環境具有黑暗、高壓、低氧、寡營養且富含高濃度硫化物、甲烷和重金屬等特征, 大型生物卻能在如此極端環境中生存并形成群落, 主要原因是它們經過長期的適應與歷史進化過程, 形成了獨特的生命特征和環境適應機制(程嬌等, 2021)。因此, 對深海化能合成生態系統中生物的環境適應機制的研究, 不僅將有助于加深對深海極端環境中特殊生命過程的理解, 也將為深海生物資源的開發利用奠定理論基礎。
近年來, 高通量測序技術的快速發展及大數據分析能力的提高, 使得在組學水平探究深海生物演化和環境適應機制成為可能。作為后基因時代應用最為廣泛的技術手段, 轉錄組測序技術可以全面快速獲得生物體在特定時空或生理條件下幾乎所有轉錄本的序列信息和表達信息, 已經在深海生物適應性演化機制研究中逐步得到應用。例如, 在雙殼綱貝類中, Zheng 等(2017)通過與淺海貽貝的比較轉錄組分析發現兩種深海偏頂蛤(Bathymodiolus platifrons和B.manusensis)中與共生菌識別相關的免疫識別受體發生收縮, 物質轉運、溶酶體活動以及細胞凋亡等相關基因受正選擇或高表達, 可能有助于共生關系的建立和維持; 在多毛類環節動物中, Zhang 等(2017)對分布于熱液區深海多鱗蟲(Branchipolynoe pettiboneae和Lepidonotopodiumsp.)與淺海近緣物種進行了比較轉錄組分析, 發現熱液多鱗蟲可能通過不同類型的血紅蛋白的快速進化或高表達等策略實現對深海低氧環境的適應; 在十足目甲殼動物中, Cheng 等(2019)對沖繩熱液與南海冷泉共有優勢物種柯氏潛鎧蝦(Shinkaia crosnieri)進行轉錄組測序和進化分析, 提出柯氏潛鎧蝦很可能是通過功能基因的適應性進化和改變基因的表達水平來適應熱液與冷泉兩種不同的深海化能環境。以上研究結果表明轉錄組學分析方法已成為探究深海極端環境中特殊生命過程的重要研究手段。然而, 由于二代測序讀取長度短, 轉錄本易發生拼接錯誤, 無法準確獲取完整轉錄本, 可變剪切和轉錄本多樣性在很大程度上仍然是未知(Zhanget al, 2020), 這限制了深海生物環境適應機制及深海基因資源開發方面的深入研究。
十足目鎧甲蝦類是深海多樣性最高的甲殼動物類群之一。在深海化能生態環境中, 鎧甲蝦是底棲生態群落中的常見種或優勢種。勞盆擬刺鎧蝦(Munidopsis lauensis)隸屬于十足目(Decapoda)、異尾下目(Anomura)、鎧甲蝦總科(Galatheoidea)、擬刺鎧蝦科(Munidopsidae)、擬刺鎧蝦屬(Munidopsis), 是深海化能生境鎧甲蝦類代表性物種, 在西太平洋勞海盆(Lau Basin)、馬努斯海盆(Manus Basin)、北斐濟海盆(North Fiji Basin)、南海東北部以及印度洋等海域的熱液、冷泉區均有分布, 分布水深在1 120~2 000 m之間(Baba, 2005; Martinet al, 2005; Cubelioet al,2007; Hwanget al, 2022)。目前對勞盆擬刺鎧蝦分子生物學方面的研究僅局限于線粒體基因組的測定(Sunet al, 2019), 以及微衛星分子標記的篩選(Boyleet al, 2013)。針對勞盆擬刺鎧蝦深海適應性的相關研究尚未開展, 且該物種基因序列信息相對匱乏, 限制了對其適應性分子機制的深入研究。本研究利用PacBio 平臺的SMRT 測序技術開展了勞盆擬刺鎧蝦全長轉錄組測序, 對獲得的轉錄本進行注釋分析, 預測其編碼序列、轉錄因子、長鏈非編碼RNA 等, 挖掘可能與其適應深海化能極端環境相關的關鍵基因和通路。研究旨在豐富勞盆擬刺鎧蝦的基因資源, 為進一步探究深海化能生態系統中大型底棲甲殼動物的進化和環境適應分子機制提供數據支持。
1 材料與方法
1.1 樣品采集
用于本研究全長轉錄組分析的勞盆擬刺鎧蝦樣品于2021 年6 月由“科學號”(中國科學院海洋研究所)科考船上搭載的遙控潛水器Quasar MkII 從位于中 國 南 海 的 臺 西 南 冷 泉(119°17.01′E, 22°06.91′N,~1 118 m)采集。樣品收集到船上后, 通過形態學初步鑒定立即進行解剖。將肝胰腺、鰓、肌肉和腸組織分離并在液氮中速凍后保存在–80 °C 條件下, 以便用于后續RNA 提取。保留部分肌肉組織儲存于濃度為70%的乙醇中, 以備于DNA 提取與COI 序列的擴增,從而進行分子鑒定。
1.2 RNA 提取
參照使用說明書, 使用Trizol 試劑盒(Invitrogen,Carlsbad, CA, USA)提取各組織的總 RNA。使用Nanodrop2000 微分光光度計(Thermo Fisher)測定總RNA 的純度和濃度, 使用Agilent 2100 生物分析儀(Agilent Technologies, Palo Alto, CA, USA)檢測總RNA 的完整度以評估RNA 的質量。不同組織檢測合格后的總RNA 等量混合, 用于下一步全長轉錄組cDNA 文庫的構建。
1.3 文庫構建、測序與數據處理
首先用 Oligo(dT) 磁珠富集 mRNA, 再用NEBNext?Single Cell/Low Input cDNA Synthesis &Amplification Module 將mRNA 反轉錄成cDNA。然后進行PCR 周期優化, 用于確定下游大規模PCR 反應的最佳擴增周期數, 之后進行大規模PCR 富集合成cDNA。cDNA 經過DNA 損傷修復, 末端修復, 并與adapter 連接。將SMRTbell 模板與測序引物退火并與聚合酶結合形成完整的SMRTbell 文庫之后, 在PacBio Sequel II 平臺(美國太平洋生物科學公司)上進行測序。
使用SMRT Link-v8.0.0 (Gordonet al, 2015)處理下機的PacBio 序列數據。首先, 從subreads.bam 文件中提取高質量的環形一致性序列(circular consensus sequence, CCS)。通過去除序列中包含的5'端引物、3'端引物和多聚體結構, 獲得全長非嵌合體(full length non-concatemer, FLNC)序列。然后使用Minimap2 (Li,2018)對同一轉錄本的FLNC 序列進行聚類, 獲得到一致性序列。再使用Quiver 算法對序列進行校正, 獲得高質量一致轉錄本。使用CD-HIT-v4.6.7 (Cluster Database at High Identity with Tolerance) (Fuet al,2012)對獲得的一致性序列去冗余, 得到勞盆擬刺鎧蝦的全長轉錄本。最后, 基于BUSCO (Benchmarking Universal Single-Copy Orthologs)的節肢動物單拷貝直系同源基因標準集數據庫, 使用 BUSCO-v3.0.2(Sim?oet al, 2015)對勞盆擬刺鎧蝦的全長轉錄本集進行完整性評估。
1.4 轉錄本的功能注釋
為了獲得勞盆擬刺鎧蝦全長轉錄本的注釋信息,將isoform 序列與NCBI 非冗余蛋白質(Nr)數據庫(http://www.ncbi.nlm.nih.gov)、Swiss-Prot 蛋白質數據庫(http://www.expasy.ch/ sprot) 、 KEGG 數 據 庫(http://www.genome.jp/kegg)和 KOG 數據庫(http://www.ncbi.nlm.nih.gov/COG)進行比對分析, 閾值設置為 1E-5。根據 Nr 數據庫的注釋信息, 使用Blast2GO (Conesaet al, 2005)軟件對全長轉錄本進行GO (Gene Ontology)注釋與功能分類。
1.5 轉錄本的結構分析
使用上述數據庫blast 比對和ANGEL (Shimizuet al, 2006)軟件預測兩種方法獲得全長轉錄本蛋白編碼區(CDS)的核酸序列和氨基酸序列。通過hmmscan 將Isoform 的編碼序列與動物TFdb 數據庫(http://www.bioguo.org/AnimalTFDB/)進行比對, 預測轉錄因子(transcription factor, TF)家族。使用CNCI-v2 (Sunet al,2013)和CPC (Konget al, 2007)軟件預測編碼能力,并將兩個軟件判定為非編碼序列的結果合集作為最終長鏈非編碼RNA (LncRNA)的預測結果。利用Perl腳本 MISA (http://pgrc.ipk-gatersleben.de/misa/misa.html)對勞盆擬刺鎧蝦的全長轉錄組進行簡單重復序列檢測。
1.6 候選基因序列分析
使用Genedoc 軟件(Nicholaset al, 1997)對勞盆擬刺鎧蝦全長轉錄組中注釋到的谷胱甘肽 S-轉移酶(glutathione S-transferase, GST)候選序列與其他節肢動物不同亞家族的GST 氨基酸序列進行多重序列比對。基于鄰接法(neighbor-joining, NJ), 使用MEGA軟件(Kumaret al, 2016)進行系統發育樹的構建, 并設置1 000 次重復。使用Genedoc 軟件對勞盆擬刺鎧蝦GST theta 亞家族的蛋白序列與包括柯氏潛鎧蝦、家蠶(Bombyx mori) 、 美州東部熊蜂(Bombus impatiens)、二化螟(Chilo suppressalis)在內的四種節肢動物的GST-theta 蛋白序列進行比對。利用SMART(http://smart.embl-heidelberg.de/)預測勞盆擬刺鎧蝦GST-theta 蛋白的結構域。
很高興收到你的來信。真的,十六年我從來不知什么叫高興。我以為高興和我無緣了。沒想到,突然收到你的信。你想我能不高興嗎?這是我十六年來第一次擁有一個最快樂的日子,比過節還開心快樂。
2 結果與分析
2.1 全長轉錄本測序數據
經COI 序列擴增、測序與NCBI 數據庫進行同源比對后, 從形態和分子層面鑒定所采樣品為勞盆擬刺鎧蝦。采用PacBio Sequel II 平臺對勞盆擬刺鎧蝦肝胰腺、鰓、肌肉和腸組織的RNA 進行全長轉錄組測序, 共獲得55 402 300 條原始序列, 平均長度為1 506 bp, N50 長度為1 587 bp。經零模波導孔(ZMW)循環過后, 通過環型一致性序列分析, 共得到1 210 829個CCS, 平均長度為1 793 bp, 循環數為43。對從CCS 得到的FLNC 序列進行聚類后獲得36 573 條一致性序列, 經過校正后的高質量一致轉錄本為36 037條。去冗余后, 獲得28 811 條高質量非冗余isoform,平均長度為2 086 bp, N50 長度為2 275 bp (表1)。BUSCO 結果顯示勞盆擬刺鎧蝦全長轉錄組中包括節肢動物核心單拷貝同源基因數的60.51% (圖1a), 表明本研究三代全長轉錄組測序獲得的非冗余轉錄本相對完整, 可繼續進行下游的數據處理。

圖1 勞盆擬刺鎧蝦轉錄本的功能注釋Fig.1 Functional annotation of M. lauensis full-length transcripts
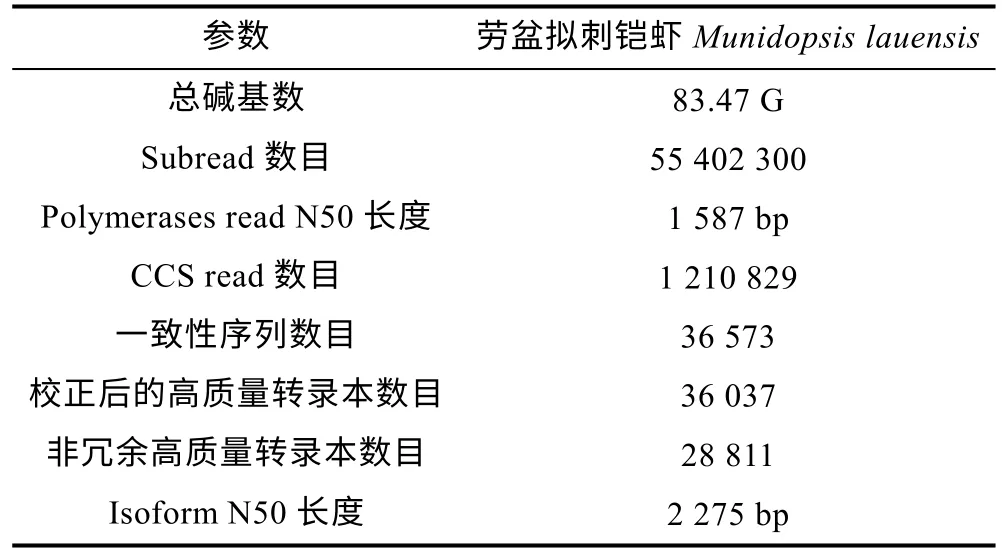
表1 基于PacBio 測序的勞盆擬刺鎧蝦全長轉錄組測序結果Tab.1 Summary for the full-length transcriptome of M.lauensis using PacBio sequencing
2.2 功能注釋
2.2.1 基本注釋 為了獲得勞盆擬刺鎧蝦更全面的遺傳信息, 將獲得的全長轉錄本在Nr、SwissProt、KOG 和 KEGG 數據庫中進行了比對分析, 共有20 616 (71.6%)條isoform 得到注釋。Nr 數據庫搜索比對顯示, 共注釋到348 個物種, 其中凡納濱對蝦(13 363; 65.01%)、克氏原螯蝦(Procambarus clarkii,881; 4.29%)和中華絨螯蟹(Eriocheir sinensis, 458;2.23%)是比對到同源序列數目前三的物種。
2.2.2 KOG 功能分類 共有17 957 條isoform 注釋到25 個KOG 分類中。其中, 占比前三位的功能分類首先是一般功能預測(general function prediction only), 具有2 990 條(16.65%)注釋信息, 其次是翻譯后修飾、蛋白質周轉、伴侶(posttranslational modification,protein turnover, chaperones)和信號轉導機制(signal transduction mechanisms), 分別有2 095 (11.67%)和1 690 (9.4%)條基因注釋信息(圖1b)。
2.2.3 GO 功能注釋 將測序獲得的基因與GO 數據庫進行匹配, 共有17 339 條isoform 被劃分到三個大類。其中基因劃分最多的類別是分子功能(molecular function, 6 669 個), 其次是生物過程(biological process, 5 955 個)和細胞成分(cellular component, 4 715 個)。分子功能類別中可以細分為18個亞類別, 其中占比最多的是結合活性(binding), 占比42.04%; 其次是催化活性(catalytic activity), 占比34.78%。生物過程類別中注釋到26 個亞類別, 占比最多的是細胞過程(cellular process), 占比 12.40%;其次是代謝過程(metabolic process), 占比11.30%。注釋到細胞成分類別中的基因被劃分為23 個亞類別,其中比例最大的為細胞(cell)和細胞組分(cell part),占比分別為 16.98%和 16.95%; 其次為細胞器(organelle), 占比為14.14% (圖1c)。
2.2.4 KEGG 功能注釋 共有20 117 條isoform 被注釋到144 條已知的KEGG 通路中, isoform 數量分布前20 條通路見表2。其中isoform 數量最多的通路是代謝途徑(metabolic pathways), 包括3 210 條isoform,其次是溶酶體(lysosome)與RNA 轉運(RNA transport),分別有551 條和439 條isoform。

表2 前20 個KEGG 代謝途徑Tab.2 Top 20 KEGG pathways
2.3 編碼序列預測與全長轉錄組結構分析
通過對勞盆擬刺鎧蝦全長轉錄本進行CDS 預測,共得到21 848 個CDS 序列。CDS 序列長度分布在102~7 719 bp, 平均值為1 049 bp; 5'UTR 的長度分布在3~4 831 bp, 平均值為266 bp; 3'UTR 的長度分布在3~7 189 bp, 平均值為970 bp。
2.3.1 轉錄因子 轉錄因子是在轉錄過程中識別和結合特定核苷酸序列的蛋白質, 通過與其他相關蛋白的相互作用介導附近基因的表達, 發揮重要的調控作用(Babu, 2010)。在勞盆擬刺鎧蝦全長轉錄組中共發現了41 個轉錄因子家族, 包含537 個轉錄因子。其中最多的轉錄因子家族是zf-C2H2 家族, 其次是轉錄因子ZBTB 和CSD 家族(圖2a)。
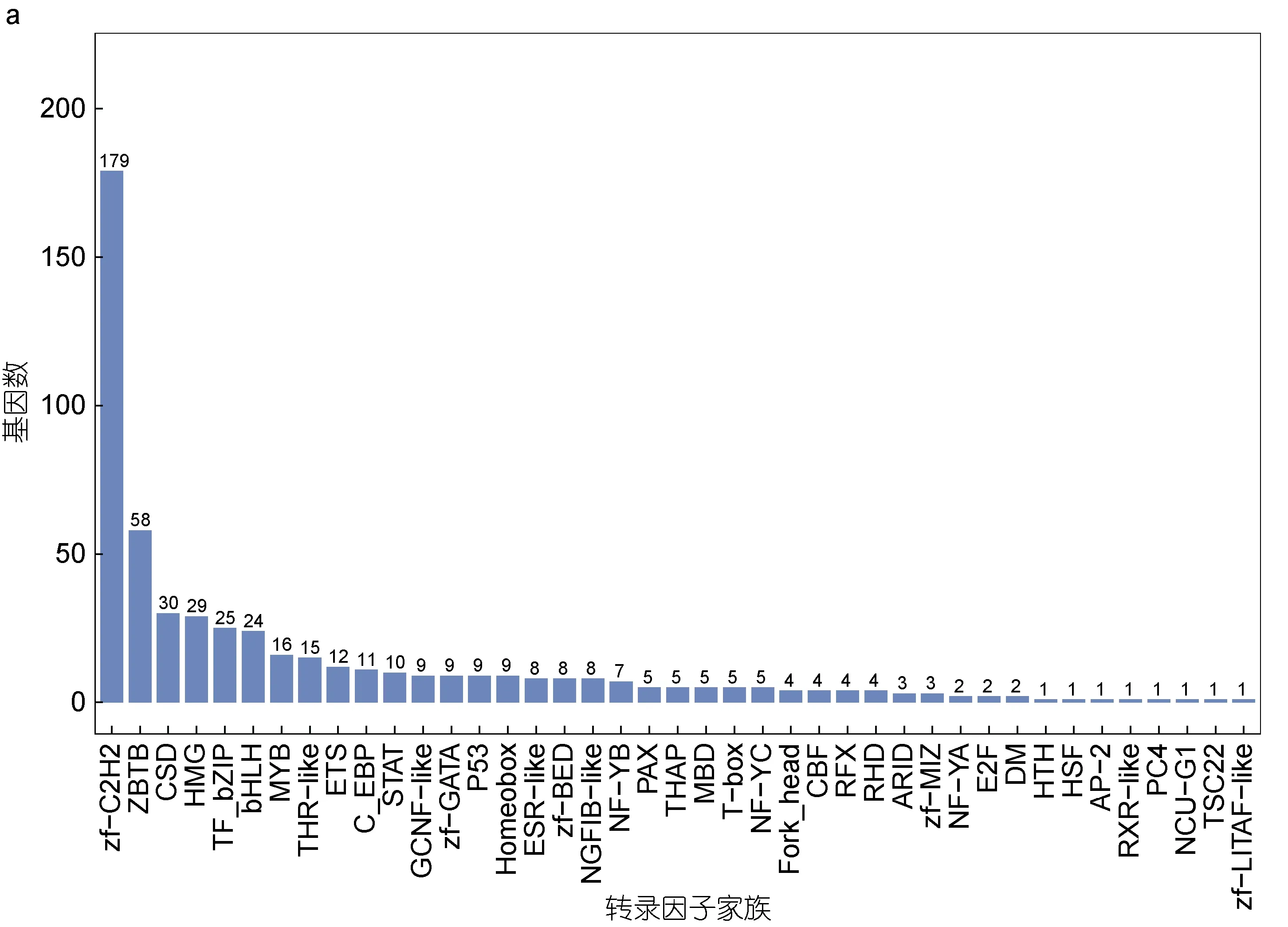
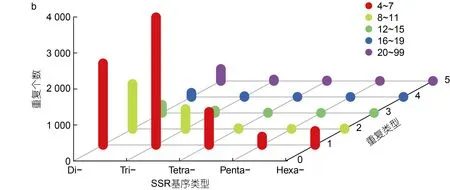
圖2 勞盆擬刺鎧蝦全長轉錄組結構分析Fig.2 Analysis of full-length transcriptome structure of M. lauensis
2.3.2 長鏈非編碼RNA LncRNA 被定義為長度超過200 個核苷酸且未被翻譯成蛋白質的RNA 分子,它們從根本上參與一些生物過程, 如轉錄、翻譯、蛋白質定位、細胞結構完整性、重編碼以及其他細胞活動(Maet al, 2013)。基于CPC 和CNCI 兩種預測方法,在勞盆擬刺鎧蝦全長轉錄組中共發現了6 430 個LncRNA。
2.3.3 簡單重復序列 簡單重復序列又稱為微衛星DNA、短串聯重復序列, 通常指以1~6 個堿基為單位組成的短串聯重復序列。本研究共檢測到6 517條包含SSR 位點的isoform, 其中1 765 條包含超過1個SSR 位點, 以復合形式存在的SSRs 數目為2 044個。進一步根據短串聯重復單元的類型對所檢測到的10 060 個SSRs 進行分類(圖2b)。依據重復核苷酸數目, SSRs 可分為5 種類型, 核苷酸重復數目為2~6 個,其中勞盆擬刺鎧蝦全長轉錄組中數量最多的是三核苷酸重復序列(4 233, 42.08%), 其次是二核苷酸重復(4 199, 41.74%)。SSRs 重復次數在4~99 次之間, 主要集中在4~7 次, 且SSRs 數量隨著重復次數的增加而降低。
2.4 候選基因及通路分析
對isoform 數量分布最多的前20 個KEGG 通路進行分析, 發現其中有兩個通路可能與勞盆擬刺鎧蝦適應深海化能極端生境相關, 分別為過氧化物酶體(Peroxisome, ko: 04146)和谷胱甘肽代謝(Glutathione metabolism, ko: 00480)。在過氧化物酶體通路中, 共有180 條isoform 編碼該通路的31 個關鍵酶, 其中脫氫酶/還原酶SDR 家族的數量最多, 為27 條; 其次是D-天冬氨酸氧化酶(21 條)和過氧化氫酶(19 條); 甲羥戊酸激酶、α-甲酰基輔酶A 消旋酶、(S)-2-羥基-酸性氧化酶、ATP 結合盒亞家族D(ALD)成員2、2-羥基酰基-CoA 裂解酶1、谷胱甘肽硫轉移酶kappa1 等數量最少, 分別有1 條(圖3a)。共有213 條isoform 參與谷胱甘肽代謝通路中19 個關鍵酶的編碼, 其中GST 的數量最多, 為59 條, 其次是前列腺素H2 D 異構酶谷胱甘肽轉移酶(25 條)和谷胱甘肽過氧化物酶(18 條), 最少的是γ 谷氨酰環化轉移酶、蛋白質二硫還原酶(谷胱甘肽)以及谷胱甘肽特異性γ-谷氨酰環轉移酶1, 數量僅為1 條(圖3b)。
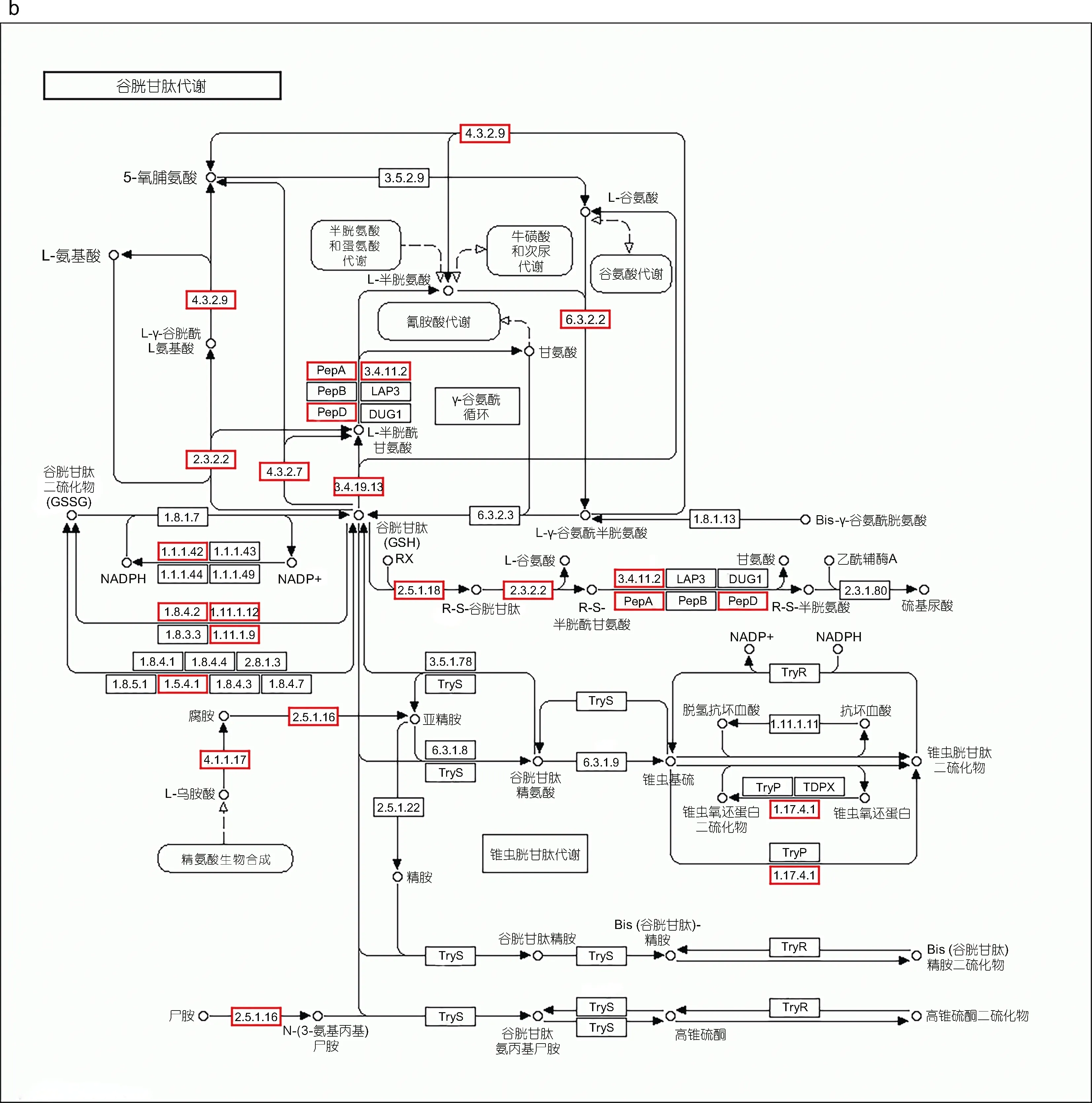
圖3 代謝通路圖Fig.3 Maps of the metabolic pathway
GSTs 屬于多功能酶, 可被分為15 個亞家族, 包括Alpha、Beta、Delta、Epsilon、Kappa、Lambda、Mu、Omega、Pi、Phi、Rho、Sigma、Theta、Tau 和Zeta (Huanget al, 2017)。勞盆擬刺鎧蝦全長轉錄組中共有20 條isoform 被分別注釋到theta、delta、mu、kappa 四個GST 亞家族中。我們利用鄰接法對勞盆擬刺鎧蝦全長轉錄組中注釋到的GST 候選序列與其他節肢動物的GST 氨基酸序列進行了系統發育樹的構建(圖4a)。結果顯示勞盆擬刺鎧蝦全長轉錄組中被注釋為不同亞家族的GSTs 分別聚類在相應亞家族的分支上。多重序列比對結果顯示勞盆擬刺鎧蝦的GST-theta 序列與柯氏潛鎧蝦的序列相似度最高, 第2~74 位氨基酸為N 端結構域, 第108~201 位氨基酸為C 端結構域(圖4b)。
3 討論
近年來, 以Pacbio 平臺為代表的第三代測序技術因其強大的讀長優勢改變了二代測序序列讀長短和碎片化的局限性, 極大地提高了獲得完整基因信息的能力(Eidet al, 2009)。全長轉錄組測序目前已成為從超大、復雜基因組物種中獲取海量基因序列信息的重要工具, 并逐步應用于水生生物生長發育、脅迫響應、物種適應性進化以及重要基因挖掘等研究(Mehjabinet al, 2019; 王菁等, 2022; Chenget al,2022; Wanget al, 2022)。本研究通過Pacbio Sequel II平臺首次對深海化能生境鎧甲蝦類的全長轉錄組進行測序和分析。與之前深海化能極端環境中的其他甲殼動物二代轉錄組結果相比(Huiet al, 2018; Chenget al, 2019), 雖然本研究所獲得的轉錄本數量(28 811 條)相對較少(143 516 條, Huiet al, 2018; 108 237 條,Chenget al, 2019), 但在轉錄本完整性和質量(N50,2 275 bp)方面卻遠高于二代(1 068 bp, Huiet al, 2018;1 125 bp, Chenget al, 2019) (圖5), 可為后續勞盆擬刺鎧蝦基因資源的挖掘提供基礎, 并為探究該物種在不同條件或不同組織的轉錄表達差異提供了參考序列。功能注釋是了解基因功能、研究基因的表達調控機制與預測蛋白功能的前提。勞盆擬刺鎧蝦各數據庫的轉錄本注釋率為71.56%, 顯著高于基于二代測序的長角阿爾文蝦(40.27%, Huiet al, 2018)和柯氏潛鎧蝦(36.78%, Chenget al, 2019)。然而仍有8 195 個轉錄本未被注釋, 推測可能是鎧甲蝦類或該物種的特有基因, 公共的數據庫中缺乏其相關的基因信息等原因造成。通過Nr 數據庫的注釋發現, 勞盆擬刺鎧蝦與凡納濱對蝦比對到的同源信息最多, 所占比例為65.01%。僅注釋到兩個擬刺鎧蝦科物種, 分別為勞盆擬刺鎧蝦(116 條)和威氏擬刺鎧蝦(Munidopsis verrilli, 2 條), 表明擬刺鎧蝦科的已有基因資源相對匱乏。GO 注釋和KEGG 注釋結果顯示勞盆擬刺鎧蝦的轉錄組功能主要集中于細胞過程與代謝過程。這些基因注釋和分類信息將有助于對勞盆擬刺鎧蝦基因功能的理解與深入研究。
轉錄因子通過特異性結合調控區域的DNA 序列來識別下游靶基因, 從而在調控各種生物過程的激活或者抑制中發揮重要作用(Charoensawanet al,2010)。本研究預測了勞盆擬刺鎧蝦的轉錄因子, 其中zf-C2H2 和ZBTB 是最豐富的轉錄因子家族。C2H2型是鋅指蛋白最常見的類型之一, 在真核生物的生長發育以及應激反應過程中起重要的調控作用(Liuet al, 2015)。ZBTB 蛋白是鋅指蛋白家族的亞家族之一, 在DNA 損傷反應和細胞發育以及先天淋巴細胞的發育和分化等過程中起重要作用(Renet al, 2019)。本結果與馬氏珠母貝(Pinctada fucata martensii,Zhanget al, 2020)、口蝦蛄(Oratosquilla oratoria,Chenget al, 2022)、大菱鲆(Scophthalmus maximus, Fuet al, 2022)的全長轉錄組解析得到的轉錄因子分析結果相似。近年來研究發現長鏈非編碼RNA 可以通過與DNA 相互作用來調節轉錄、表觀遺傳修飾、翻譯和翻譯后修飾等生物過程(Bridgeset al, 2021)。截至目前, 尚未有關于勞盆擬刺鎧蝦的長鏈非編碼RNA的報道。本研究鑒定出勞盆擬刺鎧蝦的 6 430 個LncRNA, 將為LncRNA 具體功能的進一步研究提供基礎。SSRs 廣泛存在于真核生物體內, 因其分布廣、種類多及重復單元的重復次數在個體間呈現高度特異性, 已成為檢測物種遺傳多樣性的有效工具。通過勞盆擬刺鎧蝦的全長轉錄組分析共預測到了10 060個SSRs, 結合Boyle 等(2013)對馬努斯盆地的勞盆擬刺鎧蝦開發的10 個SSRs, 可為勞盆擬刺鎧蝦在不同海盆的遺傳連通性和種群遺傳結構研究提供候選分子標記。
環境壓力可導致細胞內活性氧(ROS)水平的增高(Lushchak, 2011), 而過氧化物酶體是在ROS 的代謝中起主要作用的細胞器(Mannaertset al, 1993)。據報道, 過氧化物酶體在各種化合物包括誘導抗氧化酶的污染物刺激下在生物體內發生增殖(Orbeaet al,2002)。本研究發現過氧化物酶體通路位于isoform 數量分布最多的前20 條KEGG 通路中, 且有部分轉錄本注釋到參與過氧化物酶體生物發生的蛋白質peroxin 編碼基因上, 因此我們推測勞盆擬刺鎧蝦極有可能在深海極端環境中受到外界因子刺激后發生了過氧化物酶體增殖現象, 以應對細胞內高ROS 水平。GSTs 廣泛分布于生物體內, 主要通過協助對抗內源或者外來有害物質從而實現細胞解毒功能(楊海靈等, 2006)。真核生物中參與谷胱甘肽代謝的GSTs主要屬于MAPEG 超家族, 在內源性脂類新陳代謝以及細胞解毒方面發揮重要的生物學作用(Jakobssonet al, 1996, 1999)。鄭佩華等(2020)通過對凡納濱對蝦的GSTs 進行基因克隆, 發現MGST3 在對蝦抗逆境脅迫以及抵御病原菌感染的調控機制中發揮重要作用。Cheng 等(2019)在對深海熱液和冷泉的共有甲殼動物柯氏潛鎧蝦進行轉錄組比較分析時發現編碼GSTs 的基因在熱液個體中顯著上調表達, 推測其有助于柯氏潛鎧蝦適應理化條件更極端的熱液環境。在勞盆擬刺鎧蝦中, 我們發現了59 個編碼GSTs 的isoform, 參與谷胱甘肽代謝過程。此外還檢測到了同樣具有抗氧化作用的谷胱甘肽過氧化物酶以及參與谷胱甘肽合成的γ谷氨酰轉肽酶等。系統發育樹與多重序列比對結果顯示勞盆擬刺鎧蝦與柯氏潛鎧蝦GSTs 序列相似度高, 可能與兩物種的分類地位較近以及生存環境相似有關。不同家族的GSTs 的亞基在序列上高度進化, 但都由N 末端和C 末端兩個結構域組成。與谷胱甘肽結合的G 位點存在于N 末端結構域中, 與外源性和內源性親電子底物結合的H 位點存在于C 末端結構域中, 這兩者構成了GSTs 的催化反應活性中心(Armstrong, 1991; Ransonet al, 2005)。進一步分析發現勞盆擬刺鎧蝦的N 末端比C 末端擁有更高的序列相似性, 這種現象在 GTS 家族中也是普遍存在的(Sinninget al, 1993)。這些基因可能在勞盆擬刺鎧蝦適應高濃度硫以及重金屬等的深海化能環境過程中發揮重要作用。
4 結論
鑒于目前對深海極端環境下特殊生命過程的認識不足, 與淺海近緣物種的比較分析是解析深海生物進化與環境適應機制的關鍵。本研究利用SMRT三代測序技術對勞盆擬刺鎧蝦進行了全長轉錄組測序與分析, 不僅豐富了勞盆擬刺鎧蝦的遺傳信息, 同時為深入揭示深海化能生態系統大型甲殼動物的適應性進化機制提供了研究基礎與數據支撐。后續可基于本研究進一步開展擬刺鎧蝦科物種與其淺海近緣生物的比較轉錄組分析, 從而加深十足目甲殼動物對深海化能極端環境適應機制的理解。
