不同細胞活性物質對低溫下通草種子及幼苗的影響
2023-08-09 03:53:14龍正齡歐陽開霞
耕作與栽培 2023年3期
龍正齡, 黃 剛, 付 燕, 歐陽開霞, 趙 云
(1.黔東南民族職業技術學院, 貴州 凱里 556000; 2.黔東南生態環境監測中心, 貴州 凱里 556000)
通草[Tetrapanaxpapyrifer( Hook) K.Koch]為五加科通脫木屬通脫木的干燥莖髓,是常用的傳統中藥[1],別名大通草、花草、離南、寇脫[2],自陜西到臺灣均有分布,具有清熱利尿,通氣下乳功效[3]。近年來,通草逐漸成為帶領農村致富的鄉村產業。通草在每年秋季需將地上部植株砍掉,第二年又會萌發新芽,為多年生常綠灌木或小喬木,種一次可以延續七、八年,由于其根系有較強的繁殖能力,數量也會越來越多,產量呈逐年上漲趨勢。在種植過程中,與其他中藥材相比,通草管理相對粗放,減輕了農戶的種植工時。在生產中,可采用種子進行播種育苗,但通草種子的發芽率不高。在貴州播種育苗時容易遭受早春低溫的影響,因此提高通草種子對低溫的抗性非常重要。
硝普鈉(Sodium nitroprusside,SNP)是NO的供體,作為一種信號分子,在逆境脅迫中起到重要的信號調節作用[4]。在植物遭受非生物脅迫時,可快速合成大量滲透調節物質,同時增強抗氧化系統,減輕非生物脅迫的傷害[5-6];海藻糖(Trehalose,TH)是一種滲透調節物質,具有保護細胞內活性物質、調節細胞滲透壓,維持細胞器完整性的作用[7];赤霉素(Gibberellic acid,GA3)是一種植物生長調節劑,可以調控植物生長發育[8],能改善種子的萌發和幼苗生長狀況,緩解鹽脅迫對番茄、玉米種子及幼苗生長的傷害[9-10];種子引發是通過提高發芽率和出苗整齊度來提高種子的萌發表現,從而使幼苗更好地發育[11],同時還促進了與非生物脅迫相關的特定防御機制[11]。
本試驗從信號分子、滲透調節物質和植物生長調節劑等3個方面,采用不同細胞活性物質引發通草種子,測定種子在常溫及低溫條件下的萌發特性及幼苗生長情況,探索不同細胞活性物質引發對通草種子抗性的影響,選擇能提高種子萌發及幼苗生長特性的方法,為通草種苗繁育提供理論依據。
1 材料和方法
1.1 材 料
試驗材料為五加科通脫木通草種子,由貴州福盛天弘生態有限公司提供。
1.2 種子引發處理
硝普鈉(SNP)溶液設置50,100,200 μmol/L 3個濃度分別用A1、A2、A3表示,海藻糖(TH)溶液設置5,10,15 mol/L 3個濃度分別用B1、B2、B3表示,赤霉素(GA3)溶液設置50,100,200 μmol/L 3個濃度分別用C1、C2、C3表示。未引發的干種子記為對照1(ck1),蒸餾水引發記為對照2(ck2)。通草種子以0.5% NaClO 消毒5 min 后,用蒸餾水漂洗干凈,置于不同濃度的SNP、TH、GA3溶液中,在26 ℃黑暗條件下引發12 h。引發結束后,用蒸餾水快速沖洗種子,吸干表面水分,自然條件下晾干,備用。
1.3 種子萌發實驗
參照Khampheng等[12]的方法稍作修改。將上述SNP、TH、GA3引發的種子、未引發的干種子(ck1)及蒸餾水引發的種子(ck2)分別置于下鋪一層海綿、上墊兩張濾紙的直徑為9 cm的培養皿中,每個培養皿均勻播種100粒種子,設置3個平行重復,每個重復100粒種子,加蓋培養,發芽期間用蒸餾水保持濾紙濕潤。常溫條件設置夜溫22 ℃、晝溫28 ℃,低溫條件設置夜溫13 ℃、晝溫20 ℃,置于空氣相對濕度為75%的人工氣候箱中,每天光照13 h培養,每天記錄發芽種子數。常溫條件下發芽勢(GE)和發芽率(GP)的統計時間分別為第13天和第18天;低溫條件下發芽勢(GE)和發芽率(GP)的統計時間分別為第21天和第23天。以子葉開始展開、胚根發育正常為發芽標準。
1.4 種子各項萌發指標計算
上述各項指標種子萌發實驗結束后,統計種子的發芽勢、發芽率、發芽指數和平均發芽時間。發芽指數(GI)和平均發芽時間(MGT)計算公式如下:
GI=∑(Gt/Dt);
MGT=∑(Gt×Dt)/∑Gt,式中:Gt為第t天的發芽種子數,Dt為發芽日數[13]。
1.5 幼苗素質測定
發芽結束后(第30天)收集幼苗,用自來水沖洗干凈并用濾紙吸干表面殘余水分,測定其根長、苗高及干、鮮重。每個處理隨機選取5株,用直尺測量其根長和苗高;每個處理稱其鮮重,然后將幼苗在105 ℃殺青后在68 ℃的烘箱中烘烤24 h,稱其干重。
2 結果與分析
2.1 不同細胞活性物質引發對通草種子萌發的影響
統計在常溫條件下和低溫條件下,不同細胞活性物質引發對通草種子發芽勢、發芽率、發芽指數和平均發芽時間的影響。
2.1.1不同細胞活性物質引發對常溫條件下通草種子萌發的影響
從表1可以看出,在常溫條件下,與對照(ck1和ck2)相比,各濃度SNP、TH、GA3引發的發芽勢、發芽率、發芽指數和平均發芽時間差異不明顯,但總體上高濃度引發(200 μmol/L SNP、15 mol/L TH、200 μmol/L GA3)的發芽時間均有所縮短。其中50 μmol/L SNP處理的發芽勢最高,比ck1和ck2提高了約61%和26%;15 mol/L TH處理的發芽勢最高,比ck1和ck2提高了約39%和3%;200 μmol/L SNP處理的發芽指數最高,比ck1和ck2提高了約41%和6%;200 μmol/L GA3處理的平均發芽時間最短,比ck1和ck2縮短了0.27 d和0.28 d。
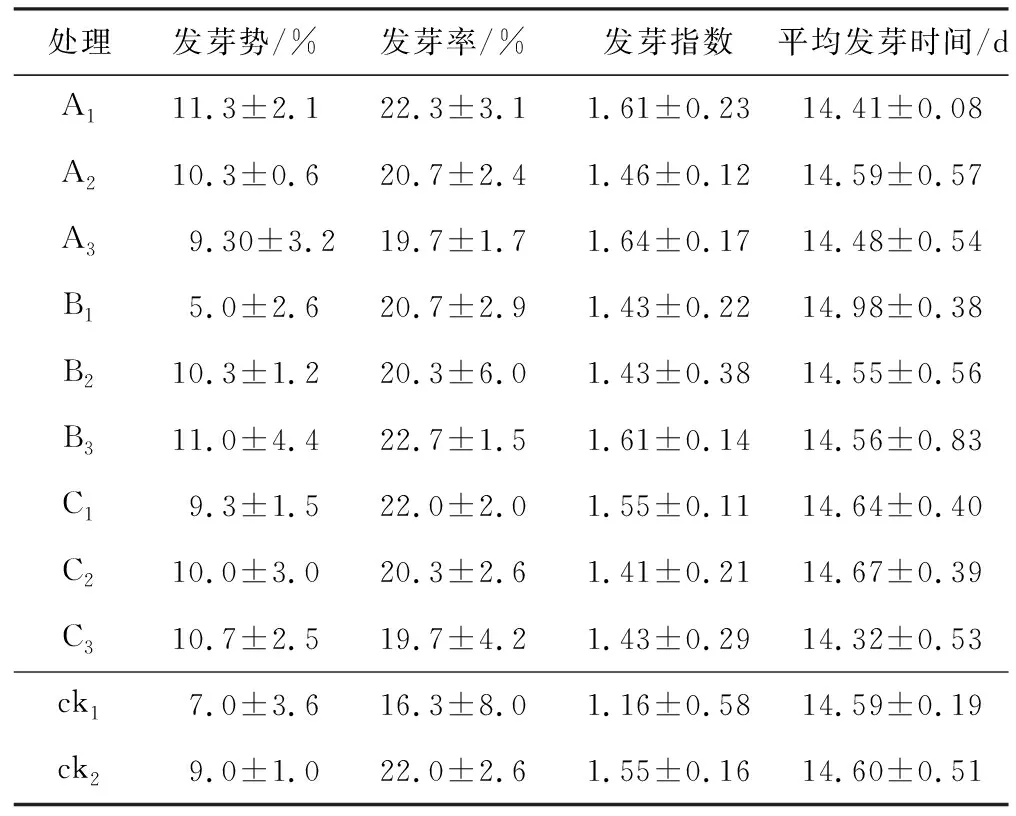
表1 不同細胞活性物質引發對常溫條件下通草種子發芽特性的影響
2.1.2不同細胞活性物質引發對低溫脅迫下通草種子萌發的影響
如表2所示,在夜溫13 ℃,晝溫20 ℃的低溫條件下,與ck1相比,200 μmol/L SNP引發明顯提高了發芽勢、發芽率、發芽指數,縮短了平均發芽時間,其次是ck2,也提高了通草種子的發芽勢、發芽率和發芽指數。除此之外,200 μmol/L GA3也在一定程度上提高了種子的發芽勢。其中經200 μmol/L SNP引發的種子其發芽勢比ck1和ck2分別提高了86%和55%,發芽率提高了41%和32%,發芽指數提高了46%和12%,平均發芽時間也縮短了約0.64 d和0.25 d(表2)。而其他處理與ck1相比,不及干種子的萌發情況。從濃度來看,每種細胞活性物質引發都是高濃度的引發效果相對較好,而低濃度不能改善在低溫脅迫下通草種子的萌發情況。
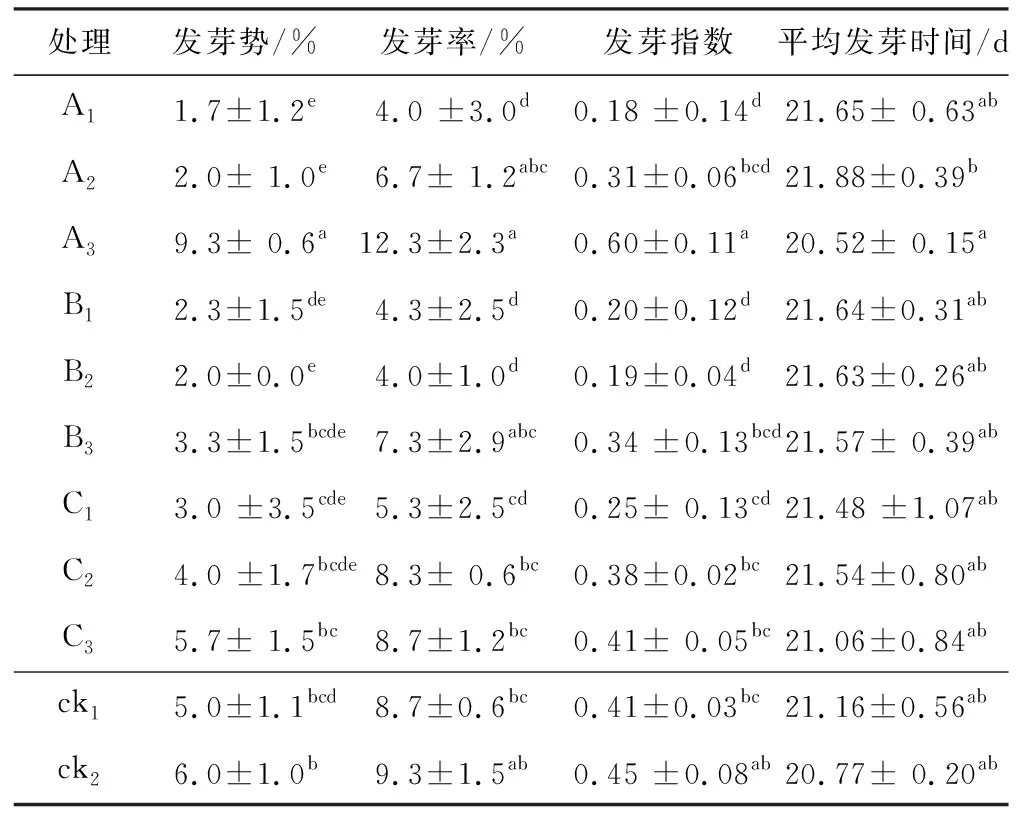
表2 不同細胞活性物質對低溫脅迫下通草種子萌發的影響
對比表1和表2可以看出,在常溫條件下,通草種子的發芽勢約在10%,發芽率在20%左右,發芽指數在1.5左右,平均發芽時間約為14 d。而在低溫脅迫下,通草種子發芽勢僅為5%左右,發芽率降低至10%,發芽指數大多降低到0.4左右,平均發芽時間也延長至21 d左右。可見低溫顯著降低了通草種子的發芽勢、發芽率、發芽指數,延長了種子的發芽時間。
進一步對各處理高濃度引發(200 μmol/L SNP、15 mol/L TH、200 μmol/L GA3)及ck1、ck2種子的發芽動態進一步監測的結果表明,與ck1相比,200 μmol/L SNP和ck2引發處理使種子在低溫脅迫下的發芽勢有顯著提高,發芽時間明顯縮短。這些結果表明,適合濃度的SNP引發能顯著提高低溫脅迫下通草種子的發芽勢、發芽率和發芽指數,縮短發芽時間,種子活力增強,顯示出較強的抗冷性(圖1)。

圖1 高濃度不同細胞活性物質引發通草種子在低溫脅迫下的發芽日變化曲線
2.2 不同細胞活性物質引發對常溫和低溫脅迫下通草幼苗素質的影響
為了進一步研究不同細胞活性物質引發在常溫和低溫脅迫下對通草幼苗生長的影響,調查了通草幼苗根長、苗高、鮮重、干重等4項生長指標。結果表明,與常溫條件下的各指標相比,低溫明顯降低了通草幼苗的干鮮重、縮短了幼苗的根長和苗高(圖2和圖3)。
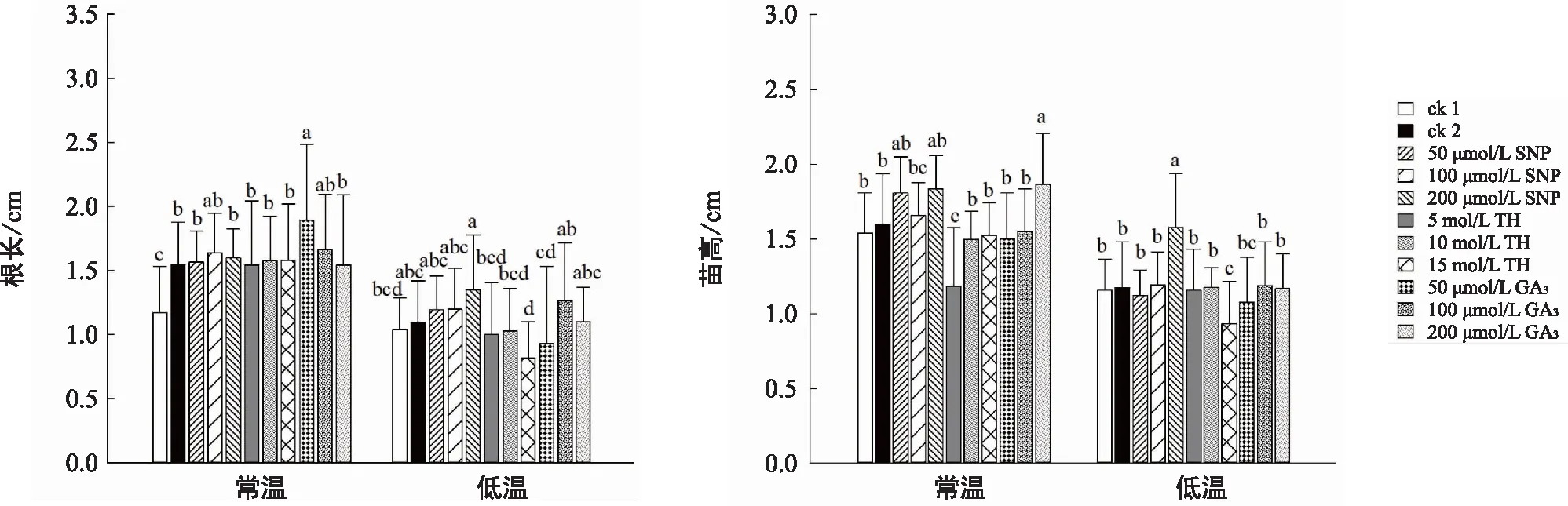
圖2 在常溫和低溫脅迫下不同細胞活性物質引發對通草幼苗根長(A)和苗高(B)的影響
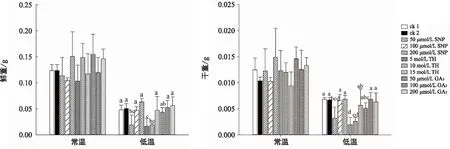
圖3 在常溫和低溫脅迫下不同細胞活性物質引發對通草幼苗鮮重(A)和干重(B)的影響
在常溫條件下,以50 μmol/L GA3處理的根長最長,比ck1和ck2提高了62%和23%。200 μmol/LGA3處理的苗高最高,比ck1和ck2提高了21%和18%,明顯高于ck1和ck2及其他處理(圖2),說明適合濃度的GA3有促進通草幼苗根莖伸長生長的作用。在常溫條件下各細胞活性物質引發的幼苗鮮重和干重與對照(ck1和ck2)相比均無明顯優勢,但50 μmol/L GA3、200 μmol/L SNP、10 mol/L TH和200 μmol/L GA3引發的鮮重高于ck1和ck2,各濃度GA3引發的干重及200 μmol/L SNP引發的干重高于ck1和ck2(圖3 B)。
在低溫脅迫下,SNP引發各處理濃度的根長都高于ck1和ck2,其中200 μmol/L SNP處理的根長比ck1和ck2分別提高了約30%和23%,其次是100 μmol/L GA3。而各濃度的TH處理的根長在低溫脅迫下都比較低,其中高濃度15 mol/L TH處理的根長最低。在苗高方面,以200 μmol/L SNP引發的苗高最高,比ck1和ck2提高了約37%和35%,而低濃度GA3和SNP處理的苗高較低,15 mol/L TH處理的苗高也最低。在鮮重方面,200 μmol/L SNP、10 μmol/L TH、200 μmol/L GA3和100 μmol/L GA3引發的鮮重均高于ck1和ck2及其他處理,其中200 μmol/L SNP引發的效果最好,比ck1和ck2提高了約32%和25%(圖2),但未達到顯著水平。而低濃度TH、SNP和GA3引發的效果較差。在干重方面,200 μmol/L SNP和100 μmol/L GA3引發處理的干重和ck1和ck2相比雖沒有明顯優勢,但仍高于其他各濃度處理,其中中高濃度處理較好,而低濃度TH、SNP和GA3引發處理效果較差。
綜合比較根長、苗高、干重、鮮重等4項幼苗生長指標,以200 μmol/L SNP引發處理的效果最好,不論在常溫還是低溫條件下都有較長的根長、苗高、干鮮重,說明經200 μmol/L SNP引發的通草幼苗對低溫脅迫有較強的抗性,能獲得較好的幼苗素質。其次是GA3引發能提高幼苗的根長與苗高,而TH引發的效果最不理想。
綜合上述,種子萌發及幼苗生長的各項指標分析表明,適宜的SNP引發能明顯提高通草的種子活力,改善在低溫脅迫下種子的萌發情況,對幼苗根長、苗高以及干鮮重也有不同程度的提高作用。而GA3和TH處理,與ck1和ck2相比,效果不是很明顯。從總體效果來看,以200 μmol/L SNP引發的通草種子及幼苗綜合素質相對較高,在低溫脅迫下仍有較強的耐受性。
3 討 論
本研究結果表明,適合濃度的SNP引發能提高通草種子在低溫脅迫下的發芽勢、發芽率、發芽指數,縮短種子發芽時間(表1、表2),提高通草幼苗根長、苗高及干鮮重(圖2、圖3),提高通草種子及幼苗的抗冷性。
通過綜合比較分析可知,與TH和GA3相比,高濃度的SNP引發能改善通草種子在低溫脅迫下種子發萌發特性和幼苗的生長情況。可能是由于SNP作為NO的供體,在低溫條件下,能作為信號分子進行一系列的植物生理調控,并且其時效性相對較長。推測在低溫脅迫時,外源一氧化氮(NO)供體硝普鈉(SNP)能作為信號分子,調節一系列的生化反應,如提高淀粉酶等活性,提高可溶性糖含量和游離氨基酸含量,為低溫脅迫下種子的萌發及幼苗的生長提供了物質來源。同時,提高抗氧化系統的酶活性來增強種子及幼苗的抗低溫能力,從而提高種子及幼苗在低溫脅迫下的抗性[9]。
而TH作為一種滲透調節物質,可能其時效性相對較短,隨著低溫脅迫時間的延長,導致其固有的對稱性穩定結構遭到破壞,其效果逐漸衰退,使其抗冷性在后期表現不明顯。GA3作為植物生長調節劑,能增加通草苗期的幼苗根長和苗高,提高幼苗的綜合素質,使得通草在低溫脅迫下仍能保持較強的抗性。而chen等[14]發現,種子引發或回干對種子來講都引起了脂質過氧化,對種子本身也是一種傷害,所以造成有些引發處理的效果不及干種子。
綜上所述,200 μmol/L SNP引發的通草種子及幼苗綜合素質相對較高,在低溫脅迫下仍有較強的耐受性,能提高低溫脅迫下通草種子的萌發情況和幼苗素質。
