祁連山島狀凍土活動層土壤氮庫對模擬凍融響應
2023-02-10 03:11:48張振華張中華王英典趙新全王文穎邵新慶杜巖功周華坤
草地學報 2023年1期
陳 哲, 徐 巍, 門 雙, 張振華, 張中華, 王英典,2,6, 趙新全, 王文穎, 孫 建, 邵新慶, 杜巖功, 周華坤*
(1.青海師范大學,青海 西寧 810016;2.高原科學與可持續發展研究院,青海 西寧 810016;3.中國科學院西北高原生物研究所青海省寒區恢復生態學重點實驗室,青海 西寧 810008;4.中國科學院青藏高原研究所,北京 100085;5.中國農業大學,北京 100083;6.青海省青藏高原生物多樣性形成機制與綜合利用重點實驗室,青海 西寧 810016;7.青海省青藏高原藥用動植物資源重點實驗室,青海 西寧 810016)
土壤凍融交替是土壤與大氣間因季節或晝夜溫差所導致的反復“凍結—融化”過程[1],全球具有凍融現象的土壤面積約占陸地總面積的70%[2]。低溫導致凍土區土壤富含有機質,其中北極和亞北極泥炭多年凍土總氮(Nitrogen,N)儲量達9.7 Pg,占全球土壤N儲量的10%[3],青藏高原凍土區0~3 m土層N儲量存約1.8 Pg[4]。而凍融過程中土體的脹縮會導致土壤團聚體結構破碎[5]、植物根系斷裂死亡[6],同時冰晶會殺死部分微生物并裂解釋放出小分子有機物等[7],這些過程會顯著改善土壤有效氮素水平[8-9]。當前,全球氣候變暖正深刻改變凍土區土壤凍融格局,例如多年凍土區土壤活動層厚度增加、表土融化期延長[10-11],冬季積雪覆蓋減少[12],這將使表土經歷更為頻繁的凍融交替[13],形成大量“有氧”“無氧”或者介于二者之間的具有獨特氧化還原條件的微域,為硝化作用和反硝化作用創造有利條件,加速凍土區土壤有機質礦化[14-15]。在全球中—高緯度不同類型高寒生態系統中,凍融作用導致土壤N2O排放量占全年總排放量的10%~80%[16],且百年尺度上N2O的輻射脅迫強度是等物質量二氧化碳(CO2)溫室效應的273倍[17]。因此,凍融過程不但導致土壤氮轉化關鍵過程發生改變,而且該時期N2O大量排放也是全球溫室氣體排放中不可忽視的“源”,對全球氣候變暖將產生正反饋效應[18]。因此,凍融交替是寒冷地區生態系統土壤養分生物地球化學循環的重要驅動力。
凍土區表層土壤作為凍融交替最為頻繁、生物化學過程最為活躍、養分最為豐富的土層,其土壤氮轉化與凍融效應密切相關。作為全球氣候變化的“敏感區”和“先兆區”,青藏高原過去幾十年正變得溫暖、濕潤。其氣溫增加幅度高于全球平均水平,達到0.16~0.67℃·(10a)-1,冬季增溫速率(0.45 ℃·(10a)-1)是夏季增幅(0.25℃·(10a)-1)的近兩倍[19]。平均溫度,特別是冬季溫度的持續上升,導致青藏高原凍土區活動層土壤開始凍結的時間以1.7 d·(10a)-1的速率延后,活動層解凍時間則以4.7 d·(10a)-1的速率提前[20],若非生長季表層土壤(0~20 cm)增溫幅度達到2.03~2.3℃時,凍土層厚度將減少14.8%,完全凍結的持續天數減少44~83天,春季表土凍融交替的天數增加37~44天[21]。另外,青藏高原近幾十年來大部分地區年均降水量增幅達3.8 mm·(10a)-1,且增加的降水主要發生在植物非生長季(11月—次年4月,3.4 mm·(10a)-1)[22]。由于水相對土壤礦質顆粒具有較大的比熱容,土壤水分含量的微小波動會引起土壤熱容量大幅改變,進而改變凍融過程中熱交換過程。因此降水時空的變化同樣影響土壤凍融格局和氮轉化。所以,氣候變化對青藏高原凍土區帶來的是溫度、降水、凍融次數的多重改變。全球范圍各類生態系統(農田、林地、草地、濕地)凍融期N2O-N通量較大波動范圍(0.7~27.2 kg·ha-1·a-1)可能與各系統中水分、溫度、凍融格局的差別有關[10,23]。因此,凍融過程中不同水、熱條件的耦合及凍融交替頻次的加劇使得青藏高原凍土區土壤氮轉化關鍵過程對氣候變化響應的不確定性增加。
盡管針對凍融過程中水分、溫度、凍融循環次數、土壤類型等因子對土壤氮循環的影響已開展多項野外原位研究或室內模擬研究,但大多以單因子試驗為主。也有多個假說試圖厘清凍融時期氮素有效性的調控機制[10,23-24],但關于長期頻繁凍融過程中氮轉化關鍵過程對不同水熱耦合響應的情況探討還比較欠缺,不利于準確評估氣候變化背景下的凍土區土壤養分轉化。基于此,本研究擬采用室內模擬試驗,監測不同梯度土壤水分和溫度組合處理在長期凍融循環過程中土壤有效氮庫組分,通過對比分析、結構方程模型等統計方法,探討青藏高原島狀凍土區高寒草甸活動層土壤氮素有效性對水、熱及凍融循環次數的響應方式,以期為近一步探討和預測氣候變化背景下,青藏高原高寒草地暖干化、暖濕化及長期凍融交替等情形可能對土壤氮循環產生的影響提供一定支撐。
1 材料與方法
1.1 研究區概況
受試土壤樣品采自祁連山脈東段大通達阪山(101°23′47.21′′ E,37°20′16.93′′ N,海拔3 705 m)。采樣點距青海海北高寒草地生態系統國家野外科學觀測研究站(簡稱“海北站”)直線距離30.7 km。該區屬典型高原大陸性氣候,冷季漫長,暖季短暫。海北站多年氣象數據表明,該地年平均氣溫—1.6℃,最暖月(7月)平均氣溫10.1℃,最冷月(1月)平均氣溫—15℃,歷史極端最高溫26.8℃,最低溫—37.1℃。年內日最低氣溫0℃以下的天數高達280天。多年平均降水量560 mm,其中80%分布于生長季(5—9月)。區域年均蒸發量1 238.0 mm。土壤樣品采集地點植被為以高山嵩草(Kobresiapygmaea)為優勢種的典型高寒草甸生態系統。植被平均高度小于10 cm,平均地上生物量210 g·m-2。采樣區土壤為斑(島)狀永久凍土區的活動層土壤,屬草氈寒凍雛形土,表層(0~20 cm)土壤有機質含量45.4 g·kg-1,容重0.90 g·cm-3,pH值7.46,含水量32.7%(6—8月)。表層土壤于10月上旬開始凍結,3月底達到最大凍結深度(187 cm),隨后表土開始融化并于6月上旬全部融通[25],全部解凍前表土持續經歷約40次晝夜凍融交替[21]。
1.2 試驗設計
本研究旨在探究長期凍融交替過程中土壤水分(W)和融化溫度(T)兩個因素不同水平下的耦合作用對土壤氮庫組分的影響。采用兩因素三水平完全隨機區組試驗設計,共9種處理。根據受試土壤持水能力,將水分梯度分別控制為體積含水量15%(W15,干燥)、30%(W30,潮濕)和45%(W45,具1 mm水層)。分別模擬降水減少導致土壤水分虧缺,正常年份生長季自然狀態和降水大量增加后土壤水分飽和三種濕度情形。鑒于海北站表層土壤10月下旬—11月中旬和3月上旬—4月中旬兩個晝夜凍融時期,近五年來此階段95%以上天數的晝夜溫度處于—13℃與13℃間波動,故我們將所有處理的凍結溫度統一設定為—20℃,將融化溫度梯度分別控制為5℃(T5,低溫)、10℃(T10,中溫)和20℃(T20,高溫),分別模擬日最高氣溫降低后低溫融化情形,正常年份的自然融化和日最高溫增加后的高溫融化情形,9個處理分別為W15T5,W15T10,W15T20(區組1),W30T5,W30T10,W30T20(區組2),W45T5,W45T10,W45T20(區組3)。各處理設3次重復,共27個受試土壤單元。以各處理在實施第一次模擬凍融交替前一天的土壤狀態為對照。
在預先設定好的三塊30 m×30 m的樣地中,采取梅花布點的方式,布設5個50 cm×50 cm的樣方框,齊地剪取植物地上生物量,帶回實驗室烘干至恒重測定生物量。用直徑10 cm根鉆在剪除地上植物后的樣方中采集0~20 cm足夠的土壤,帶回實驗室,剔除枯枝、落葉后,輕揉土塊,盡量減少土壤團聚體的人為物理破碎,過2 mm土篩,剔除植物根系、石礫等。稱取500 g過篩后的土壤,轉入體積1 L具塞玻璃廣口瓶,搖晃瓶體保證土壤與瓶體均勻接觸且上表面平整。瓶中土壤高度8~10 cm,土表距瓶口11~13 cm。采用便攜式土壤濕度計測定各受試樣平土壤體積含水量,用超純水將其調至處理所設水平。水分梯度設置好后,記錄各樣品瓶總質量,做為后期補充水分的依據。
由于土壤采樣期,研究區日最高溫近20℃,為盡可能使受試土壤與冷季初期凍結前的自然狀況相接近,對所有土壤樣品做低溫馴化處理。做法為將所有樣品先后在15℃,10℃和5℃培養箱各培養1周,再使用3臺具有制熱和制冷雙重功能的培養箱進行晝夜凍融交替模擬處理,將凍結溫度和融化溫度持續時間均設為12小時(8:00—20:00融化,20:00—次日8時凍結),利用機器溫控自動循環程序進行晝夜溫度調整。在低溫馴化和模擬凍融交替過程中每5天測定一次瓶體總質量,根據質量變化補充因蒸發導致的土壤含水量的下降,保證土壤水分穩定。共模擬149天(次)晝夜凍融交替。
1.3 樣品測定方法
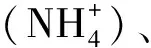
1.3.2微生物量測定 微生物量氮(Microbial biomass nitrogen,MBN)含量,主要通過氯仿熏蒸的方法完成。步驟如下:稱取經前處理的12.5 g新鮮土壤樣品,置于50 mL燒杯中,將燒杯放入盛有氯仿的真空干燥器中,用真空泵以—0.07 Mpa壓力抽真空,氯仿劇烈沸騰3~5 min停止,重復抽真空2次。關閉干燥器閥門,于25℃黑暗條件下熏蒸24 h。同時做未熏蒸對照組。熏蒸結束后用0.5 mol·L-1K2SO4溶液,按土水比1∶4(w∶v)混合液,混合液在25℃條件下,轉速200 r·min-1的震蕩器上震蕩30 min,用中速定量濾紙過濾。取10 mL過濾液,用TOC/TN分析儀(multi N/C 3100,Analytikjena,Germany)測定浸提液總氮(Total nitrogen,TN)含量[26]。
土壤微生物量氮(MBN)含量計算方法如公式:
式中EN為熏蒸土壤浸液提測定的TN與不熏蒸土壤浸提測定的TN的差值;KEN取0.54[27]。
1.4 數據統計
各處理和對照的測量指標在SPSS(IBM SPSS Statistics 20.0,SPSS Inc.,Chicago,USA)中利用單因素方差分析(ANOVA)并結合LSD法進行多重比較,顯著性水平選取α=0.05。所有數據在SPSS中經標準化轉換后,使用Amos(IBM SPSS Amos 24.0,SPSS Inc.,Chicago,USA)軟件構建微生物量與環境因子和氮庫組分間的結構方程模型,并計算各路徑系數。使用Origin(OriginPro 2020b,OriginLab Corp.,Northampton,USA)制圖。
2 結果與分析
2.1 土壤有效氮組分變化
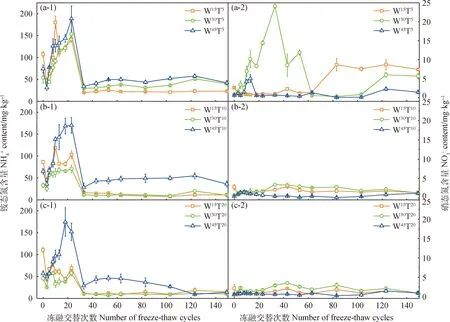
圖1 長期凍融交替過程中土壤銨態氮、硝態氮含量動態特征

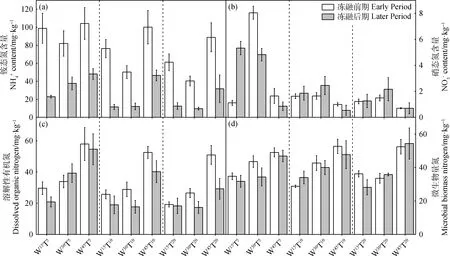
圖2 不同處理下凍融前期和后期氮庫組分均值差異性
(3)DON含量
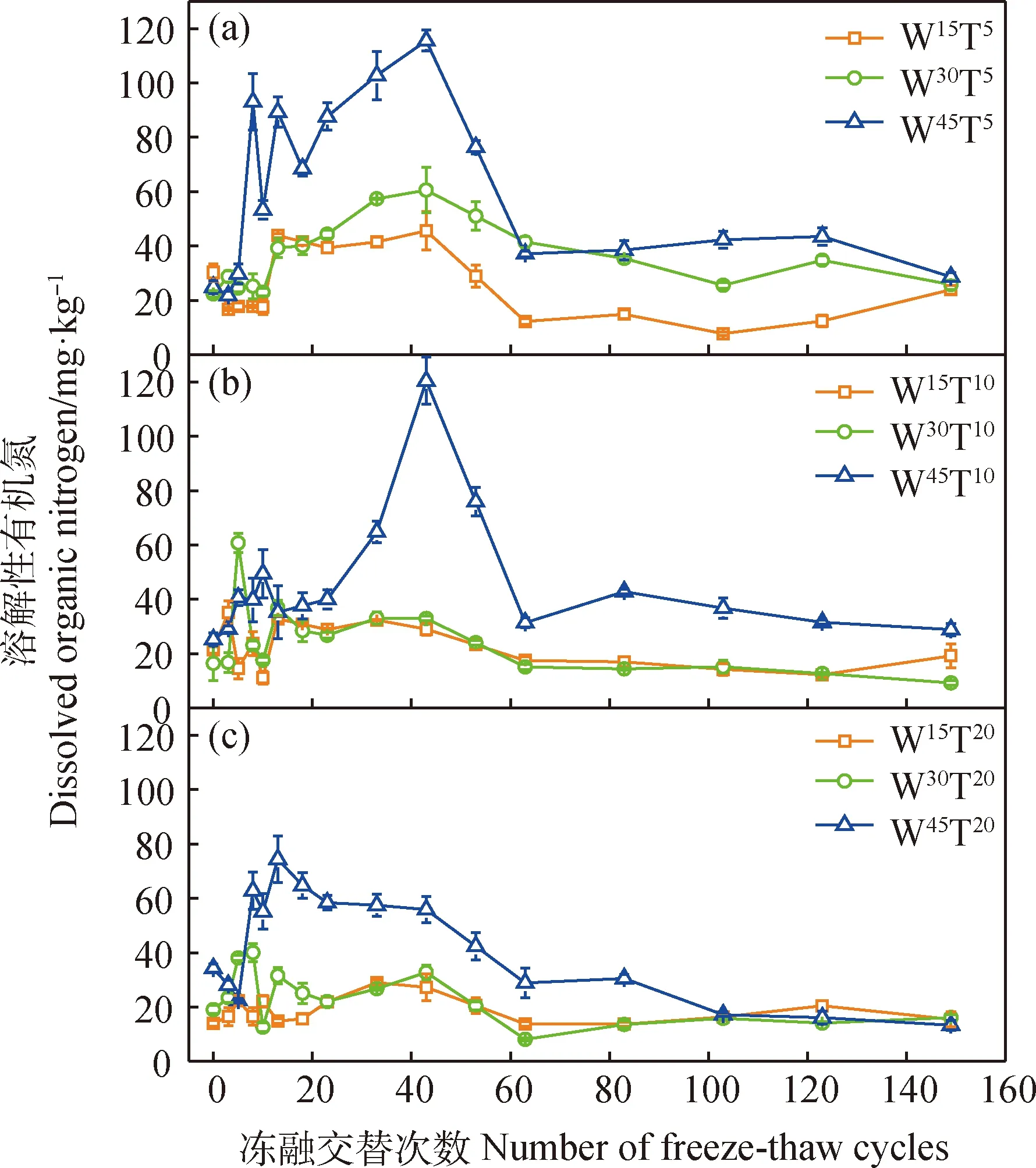
圖3 長期凍融交替過程中土壤溶解性有機氮含量變化特征
2.2 土壤微生物量變化
不同水分條件下MBN對長期凍融的響應有所區別,其中W30和W45區組MBN也與土壤速效氮一樣呈現前期增加后期降低的趨勢,峰值較凍融前對照增加1~2倍(圖4),但W15區組在整個試驗周期中均無顯著波動。W30和W45區組中除W30T20處理外,其它處理MBN前期累積量均值(42.67~52.66 mg·kg-1)顯著高于W15區組各處理(28.77~36.22 mg·kg-1)。相同水分條件下,不同融化溫度處理并未顯著改變MBN動態規律及凍融前期和后期的平均值(圖2-d)。另外,在低溫(5℃)融化環境下,長期處理反而使得MBN積累有明顯的增長趨勢,而高溫條件下凍融后期MBN含量回歸到與凍融前對照相當的水平(圖4a)。表明MBN對凍融過程中水分響應相比融化溫度更強烈,長期低溫有利于微生物氮固持。
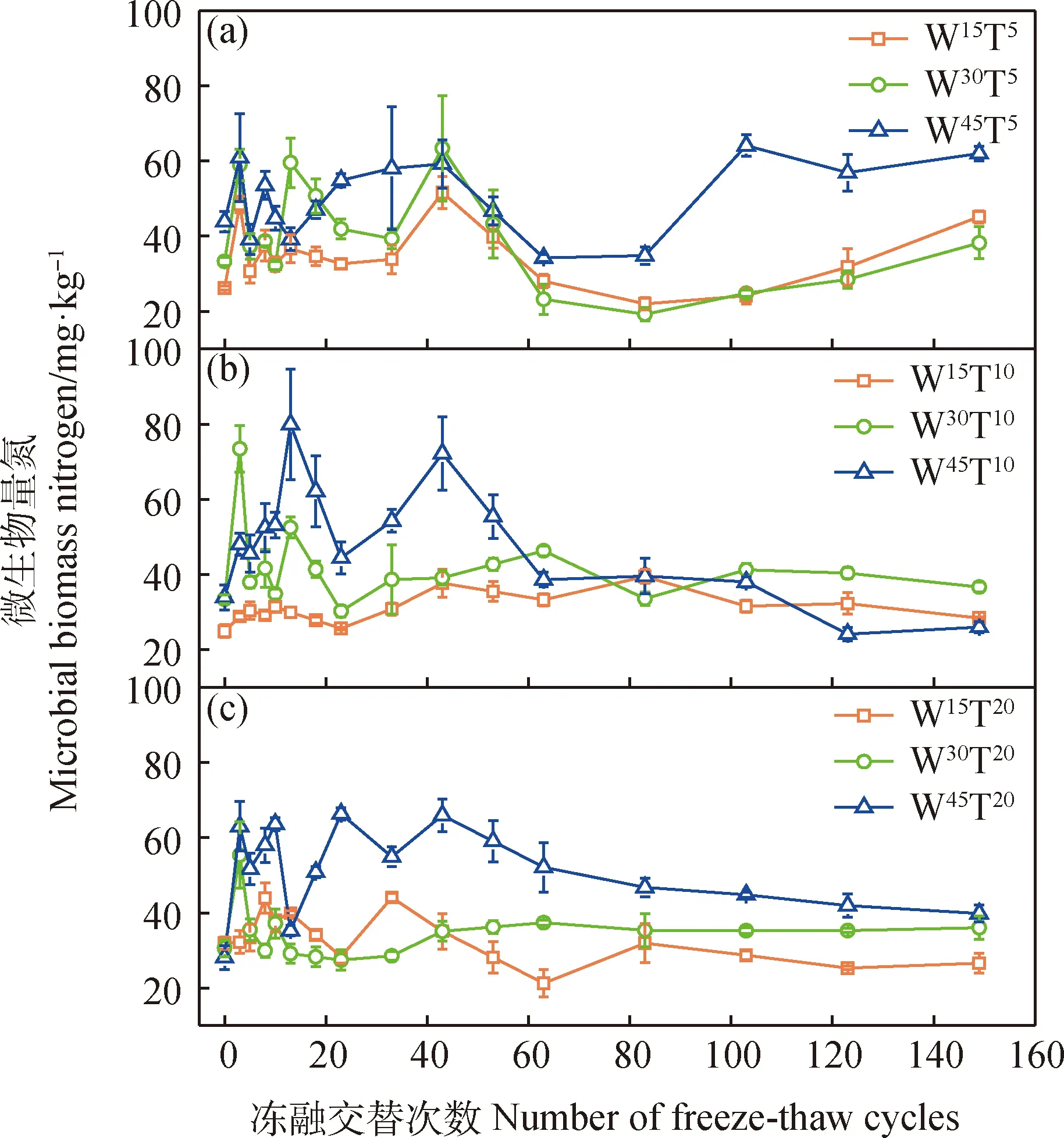
圖4 微生物量氮含量動態特征
2.3 調控土壤氮庫組分的因子分析
為探明長期凍融過程中土壤水分、凍融溫度、凍融頻次對土壤氮庫氮素有效性的影響,我們采取結構方程模型的方法量化了環境因子和不同形態氮含量對土壤氮素有效性的影響,結果表明隨著土壤水分的增加,其通過促進MBN含量(路徑系數0.328)和DON含量(路徑系數0.376),進而提高氮庫有效性,總效應為0.329;凍融的頻次和融化溫度均對土壤微生物量氮含量無顯著關系,其主要通過調節DON含量,影響氮庫有效性,但與水分不同的是,凍融頻次和融化溫度對DON含量均具有顯著負效應,路徑系數分別為—0.211和—0.229,二者對氮庫有效性的總效應分別為—0.128和—0.139(圖5)。可見隨著凍融頻次和融化溫度的增加,長期高溫條件下的頻繁凍融交替會導致土壤氮素有效性衰減。
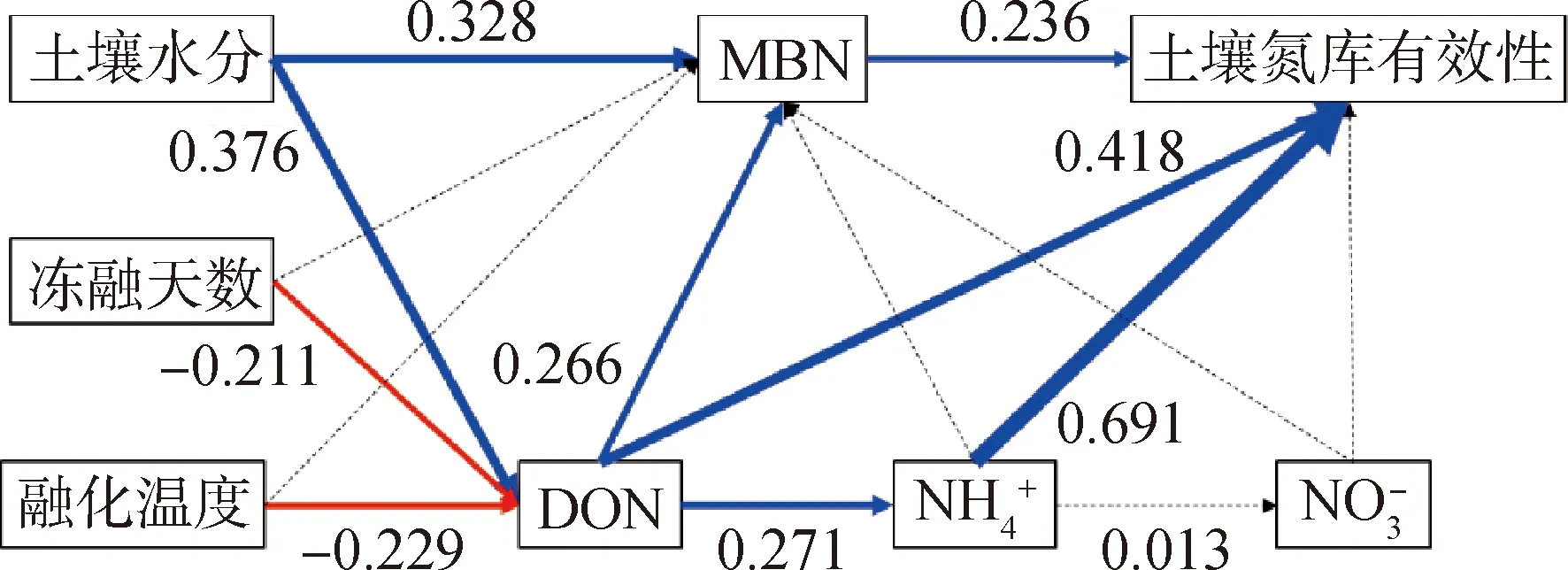
圖5 影響土壤氮庫有效性的結構方程模型分析
3 討論
3.1 凍融頻次對土壤氮庫的影響
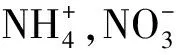
3.2 凍融溫度對土壤氮庫的影響
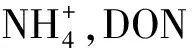
另外,W45區組的土壤氮庫對融化溫度響應不敏感的主要原因可能與高水分條件下土壤高的比熱容有關。因為水分凍融過程中發生相變會吸收或釋放大量熱量,影響土壤熱量分布和熱量傳遞過程[41]。且土壤熱傳導率和液態含水率呈現正相關關系,即在土壤凍結過程中,液態水相變會降低土壤的熱傳導率,也就是說高含水量的土壤發生凍結后將減緩大氣熱量向土體內部的傳導[42]。因此凍融過程中復雜的水熱關系導致即便在相似的融化溫度下,凍融過程中潮濕土壤與干燥土壤的溫度波動、凍結持續時間、水分狀態、土壤空隙等可能存在顯著差異[43]。故水-熱-氣條件的同步變化造成凍融過程中凍土區土壤物質轉化具有較大的不確定性。
3.3 水分對土壤氮庫的影響

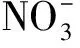
4 結論