高寒草甸土壤細菌群落多樣性和共現性網絡對牦牛覓食強度的響應
2023-02-10 02:52:40孟令旭賈彩玲許春雪張瀟文
草地學報 2023年1期
孟令旭,賈彩玲,許春雪,張瀟文,呼 和
(內蒙古大學生態與環境學院,內蒙古 呼和浩特 010000)
微生物是生態系統的重要組成成分,能夠廣泛參與生態系統的物質循環和能量流動等過程[1]。細菌群落對草地生態系統中土壤養分存儲和植物生長等過程起到了重要作用[2]。此外,細菌群落通常對環境變化比較敏感,能夠對植被和土壤等環境因素變化迅速作出響應,因此經常被用來指示生態系統變化[3]。
動物覓食能夠通過采食、踐踏和排泄等行為改變土壤理化性質,從而影響土壤細菌群落[4]。研究表明,動物采食通過影響地上植被來改變土壤碳和氮的分配[5];踐踏能夠改變土壤孔隙度以及水的導流能力;動物排泄導致的糞便沉積通過循環利用營養物質,改變了土壤氮含量[4]。這些土壤性質的改變嚴重影響了土壤細菌群落。以往動物覓食對生態系統影響的研究大多集中在地上植物群落[6-7]。例如,荒漠草原放牧通過降低植物多樣性,調節生態系統功能[8],降低草原生態系統的生產力[9];半干旱草原放牧通過改變植物特征,從而保護生物多樣性并提高植物群落恢復力[10];高寒草原放牧通過影響植物多樣性來調控草地生產力的分配[11]。然而,細菌對調控生態系統的穩定性等方面同樣有著重要作用[12],如,農業生態系統穩定性[13]和荒漠生物結皮的穩定性[14]。此外,Xun等[15]研究表明細菌群落維持了土壤氮代謝和磷酸鹽代謝等功能,有利于改善生態系統服務功能。細菌在生態系統中通常不會以單一個體的形式存在,他們之間存在競爭或互利共生等相互作用。微生物類群間相互作用不僅能夠影響土壤碳礦化速率[16],還對土壤甲烷循環等生態功能起到了重要作用[17]。Cameron等[18]研究表明土壤細菌類群間相互作用對維持草地生態系統的多功能性起到了重要作用。越來越多研究表明,共現性網絡是研究微生物類群間相互作用的重要手段[19-20]。共現性網絡不僅能夠反映物種間復雜的相互作用,還能夠更好的預測生態功能多樣性[21]。然而,我們對于動物覓食行為導致的高寒草甸土壤細菌多樣性和共生模式的改變及其調控因子的認識還不足。
青藏高原是亞歐大陸最大的地貌單元,其草地面積約占全國草地總面積的三分之一,對全球氣候變化調節、水源涵養和生態系統服務功能等方面具有重要作用[22-23]。此外,青藏高原生態系統比較脆弱,容易受到氣候變化和人為等因素影響[24],動物覓食則是影響青藏高原生態系統的一個關鍵因素。本文通過對牦牛不同覓食強度下土壤理化性質和細菌群落的變化進行分析,探究青藏高原高寒草甸土壤細菌群落組成、多樣性和共現性網絡對牦牛覓食的響應特征,明確不同覓食強度下土壤理化因子的變化規律以及對細菌群落產生的影響,以評估生態系統的變化過程,這對青藏高原的合理利用具有重要意義。
1 材料與方法
1.1 研究區域和實驗設計
研究場地位于青藏高原東緣的甘肅省甘南州瑪曲縣中部阿孜畜牧科技示范園區(33°40′ N,101°52′ E),該地區海拔3 650 m,主要土壤類型為亞高山草甸土,草地類型屬于典型高寒草甸草原,氣候為高原大陸性季風氣候,年均氣溫1.2℃,年均降水量620 mm,主要集中在5—9月,年日照時間約2 580 h,年平均霜日大于270 d,沒有四季,只有冷暖季之分。牧草每年5月初開始返青,9月開始逐漸枯黃。
2009年開始,在甘南瑪曲阿孜畜牧科技示范園區附近選擇放牧草地做為樣地,長期以來,該樣地都以自由放牧為主,且放牧時間和管理模式相同。綜合之前的研究方法[25],為了減少牦牛覓食距離造成的實驗誤差,根據草地到牛圈的距離、牦牛的行走路徑、牦牛理論采食量、采食時間、樣地面積和對應的牦牛數量將其劃分為4個不同覓食強度梯度,本實驗設計還充分考慮了牦牛行為和放牧時長等因素,依次選取距離牛圈入口250 m,500 m和1 000 m的區域分別作為重度覓食(HFI)、中度覓食(MFI)和輕度覓食(LFI)的采樣區域[25],圍封1個禁止采食區域作為對照(CK)(圖1)。相對放牧強度(yak units·hm-2)=牧戶存欄數/不同梯度區域面積(不同半徑的同心圓面積)[25-26],計算得到CK,LFI,MFI和HFI的覓食強度分別為0,2.6,3.5和6.5 yak units·hm-2,牦牛每天采食時間為8 h。
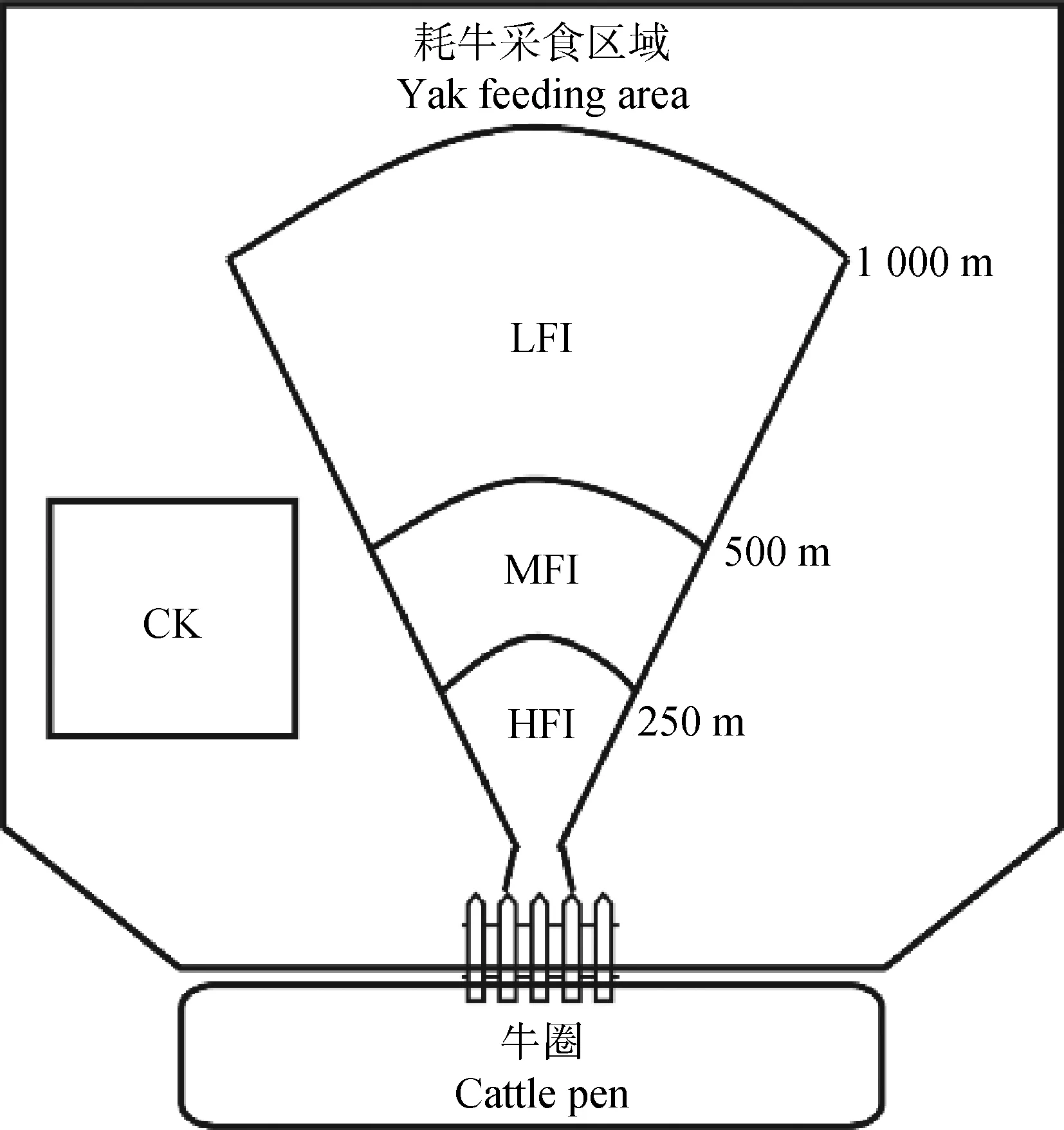
圖1 實驗設計示意圖
1.2 樣品采集及處理
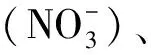
1.3 土壤理化性質測定
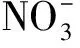
1.4 DNA提取和高通量測序
根據制造商說明,使用土壤FastDNA試劑盒(MP Biomedical,Carlsbad,CA,USA)從0.5 g土壤中提取DNA,用1%瓊脂糖凝膠電泳評估DNA質量。使用細菌通用引物338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和806R(5′-GGACTACCAGGGTATCTA-3′)完成細菌16S rRNA基因V3-V4區段序列擴增,擴增產物按照AxyPrep DNA(Axygen Biosciences,Union City,CA,USA)凝膠提取試劑盒說明書純化。最后使用Illumina Miseq平臺測序,去除不明確的堿基和低質量序列,最終堿基平均質量分數不能小于25,堿基序列長度不能低于200 bp,以97%的序列相似性將高質量的序列聚類定義為操作分類單元(OTUs)。
1.5 共現性網絡構建
OTU相對豐度被用來構建不同覓食強度下的共現性網絡,選取每個處理中至少3個樣本同時出現的OTU進行網絡分析。我們使用基于3個測量值的集成方法,包括Spearman相關性和OTU兩兩之間的Bray-Curtis和Jaccard差異,當相關閾值|r|大于0.6且P值小于0.01時,共現性網絡被認為是類群之間統計上穩健的相關[27]。利用Brown方法對3種統計值進行合并,然后利用FDR方法對P值進行校正以減少獲得假陽性結果的機會[27-28]。最終使用Gephi 0.9.2(https://gephi.org)進行網絡可視化。
1.6 統計分析
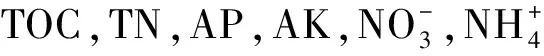
2 結果與分析
2.1 土壤理化性質

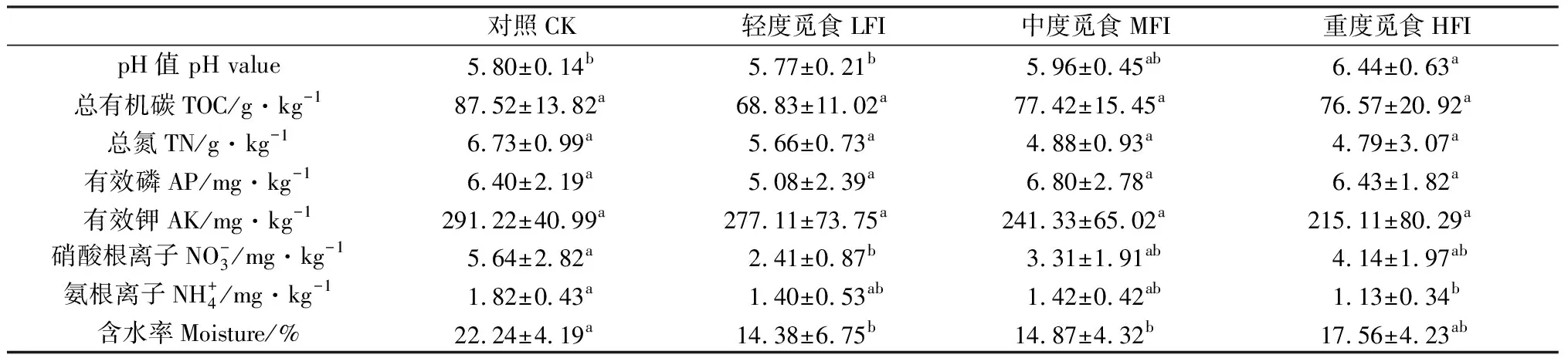
表1 不同覓食強度的土壤理化性質的ANOVA檢驗
2.2 土壤細菌群落組成
根據不同覓食強度下細菌的OTU數量繪制韋恩圖,由圖2可知,不同覓食強度處理下共檢測到3 427個細菌OTUs,CK,LFI,MFI和HFI特有的細菌OTU數分別為261,331,291和573個,可見牦牛覓食增加了土壤中特有的微生物群落。
高寒草甸草原牦牛不同覓食強度下細菌菌門組成相同,然而豐度不同。門水平相對豐度高于1%的細菌依次為變形菌門(Proteobacteria)、放線菌門(Actinobacteria)、厚壁菌門(Firmicutes)、酸桿菌門(Acidobacteria)、綠彎菌門(Chloroflexi)、擬桿菌門(Bacteroidetes)、疣微菌門(Verrucomicrobia)和芽單胞菌門(Gemmatimonadetes),其中變形菌門、放線菌門相對豐度最高,分別為30.66%~33.38%和25.04%~28.99%,2者共占土壤細菌總reads數的50%以上(圖2)。利用Kruskal Wallis對不同牦牛覓食強度下土壤細菌門水平的相對豐度進行差異顯著性檢驗,結果表明只有疣微菌門(2.46%~5.10%)和芽單胞菌門(1.91%~2.68%)存在顯著差異(P<0.05)。
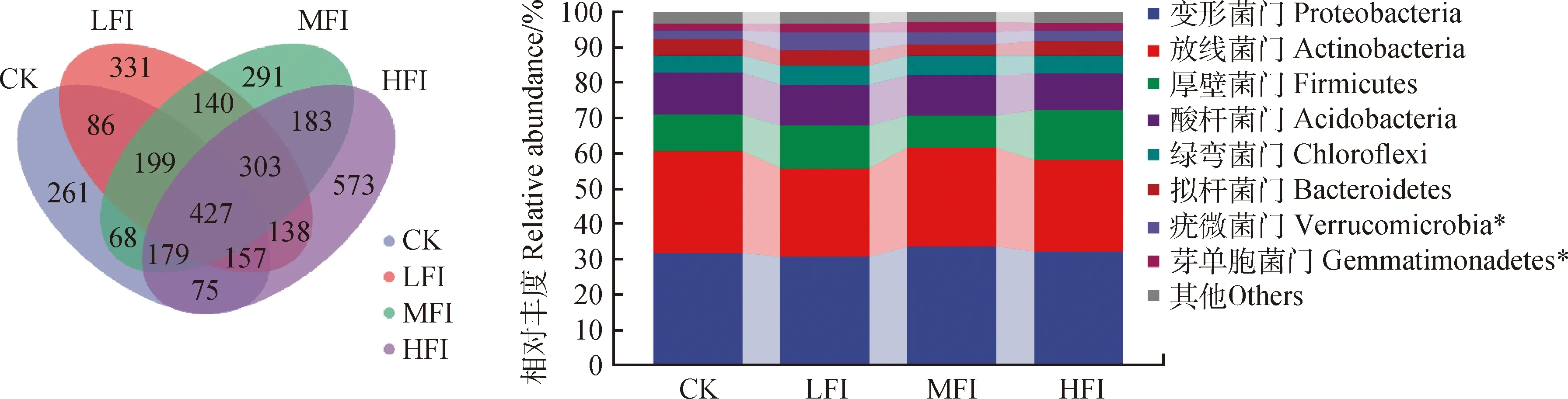
圖2 土壤細菌OTUs數量的韋恩圖和不同覓食強度細菌門水平物種組成的變化
2.3 土壤細菌群落alpha和beta多樣性
群落多樣性揭示了牦牛不同覓食強度下細菌群落的分布模式。不同覓食強度下土壤細菌Shannon-wiener多樣性指數和chao1指數均無顯著性差異(圖3),表明牦牛覓食強度對土壤細菌alpha多樣性影響較小。
基于Bray-Curtis距離的主坐標分析(PCoA)揭示了牦牛不同覓食強度下細菌群落的分布模式(圖3)。非參數的多元方差分析(PERMANOVA)表明,牦牛覓食顯著改變了細菌群落組成(P<0.05),其中,CK和HFI的細菌群落組成均與MFI呈現出顯著差異(P<0.05)。
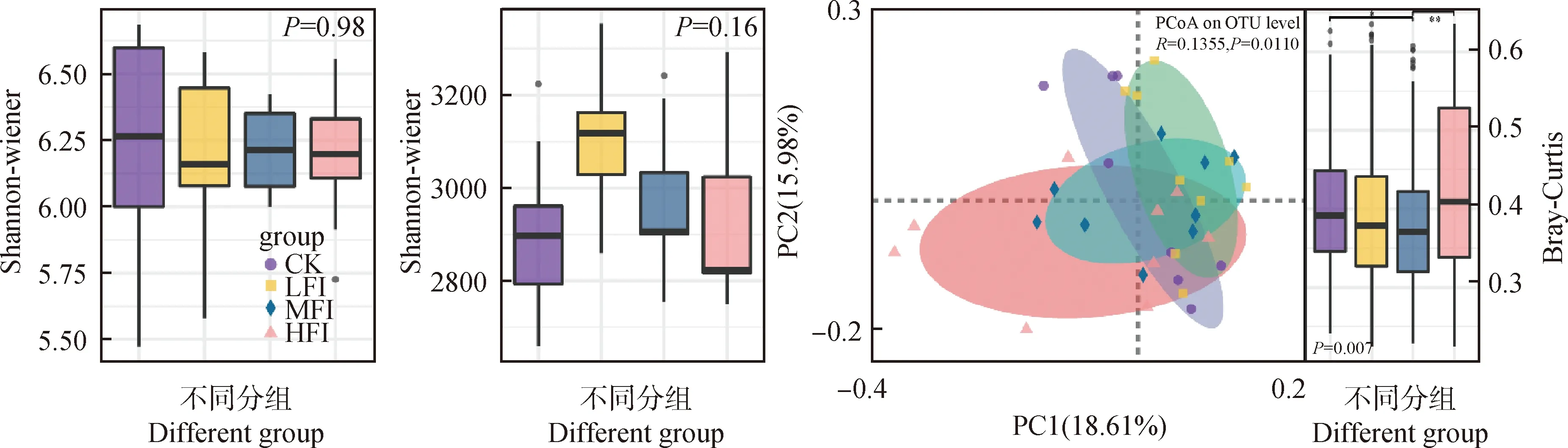
圖3 不同覓食強度細菌群落的alpha多樣性和beta多樣性
2.4 土壤細菌共現性網絡
為了研究不同覓食強度下細菌潛在的相互作用以及共發生模式,本研究基于隨機矩陣理論,采用相同的閾值(R>0.6,P<0.01),使用4種覓食強度下的細菌群落分別構建分子生態網絡(圖4a)。對各共現性網絡的節點數量進行統計發現,變形菌門(Proteobacteria)(27.13%~35.09%)、放線菌門(Actinobacteria)(21.81%~24.91%)和酸桿菌門(Acidobacteria)(15.43%~17.67%)是4個網絡中的優勢菌門(圖4a)。牦牛覓食能夠增加網絡中疣微菌門(Verrucomicrobia)和芽單胞菌門(Gemmatimonadetes)的節點數量。其中,隨著共現性網絡中疣微菌門相對豐度的增加,細菌群落beta多樣性及網絡復雜度降低(圖4b)。
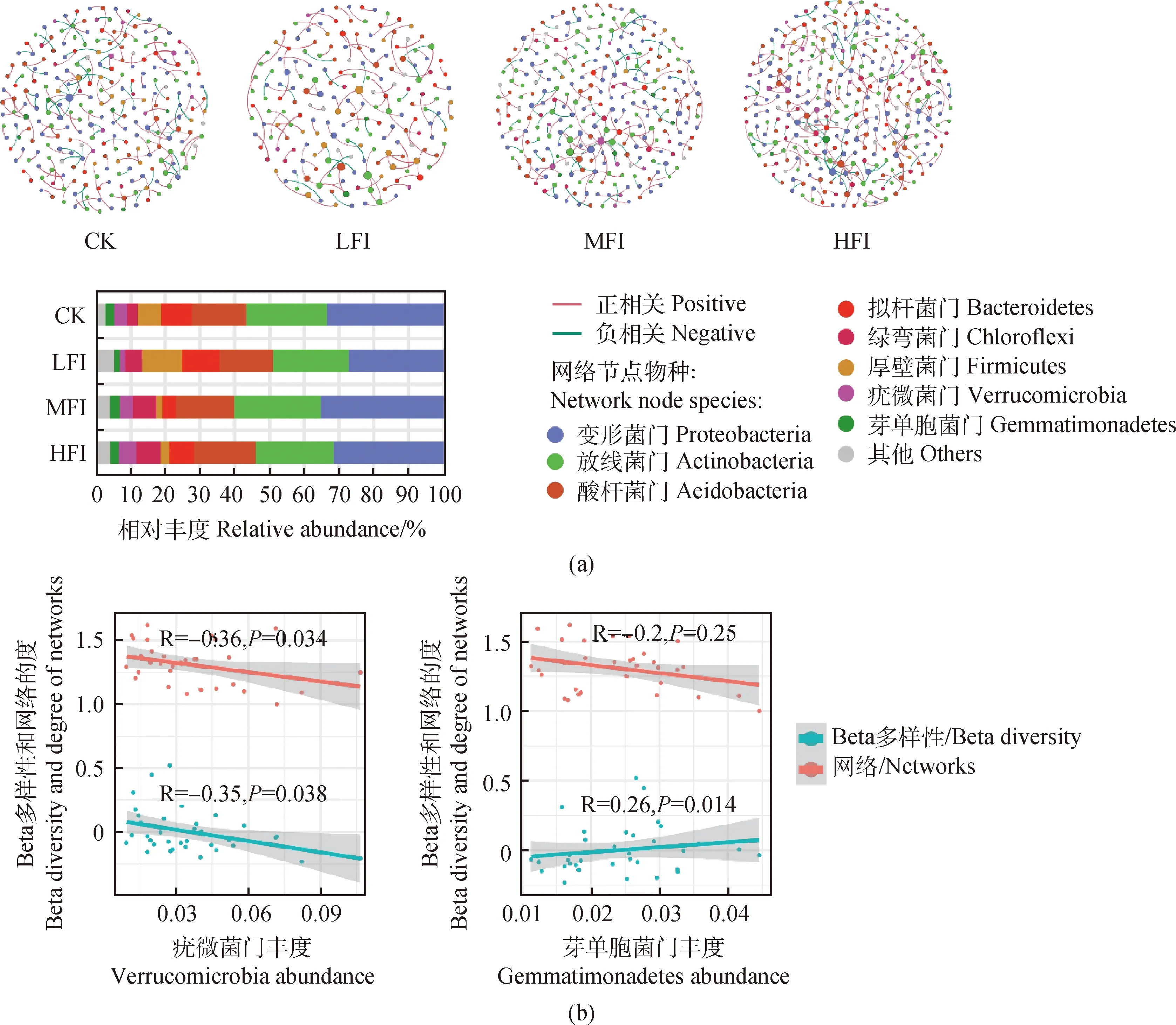
圖4 不同覓食強度對細菌共現性網絡及細菌群落多樣性的影響
牦牛覓食強度能夠顯著改變細菌共現性網絡的節點和邊的數量以及連接度等多種網絡屬性(表2)。網絡中節點、邊和度能夠代表網絡的復雜度,隨著牦牛覓食強度增加,網絡節點、邊的數量和連接度均呈現先減少后增加的趨勢,說明牦牛覓食能夠增強網絡復雜度。無論覓食強度如何改變,細菌共現性網絡節點之間正相關比例均大于負相關。模塊化程度和聚類系數反映了網絡中節點聚在一起的程度,不同處理中網絡模塊化程度(0.967~0.984)均很高,表明網絡能夠通過模塊發揮相應功能。CK處理中具有較高的平均聚類系數,因此該處理中網絡節點的聚集程度更高。我們使用Kolmogorov-Smirnov檢驗方法對網絡節點、邊和度進行差異性檢驗,結果表明牦牛覓食能夠顯著改變網絡屬性(P<0.05)(表3)。無論哪個處理,兩兩網絡之間的屬性均具有顯著差異(P<0.05),說明牦牛覓食能夠對細菌共現性網絡產生重要影響。
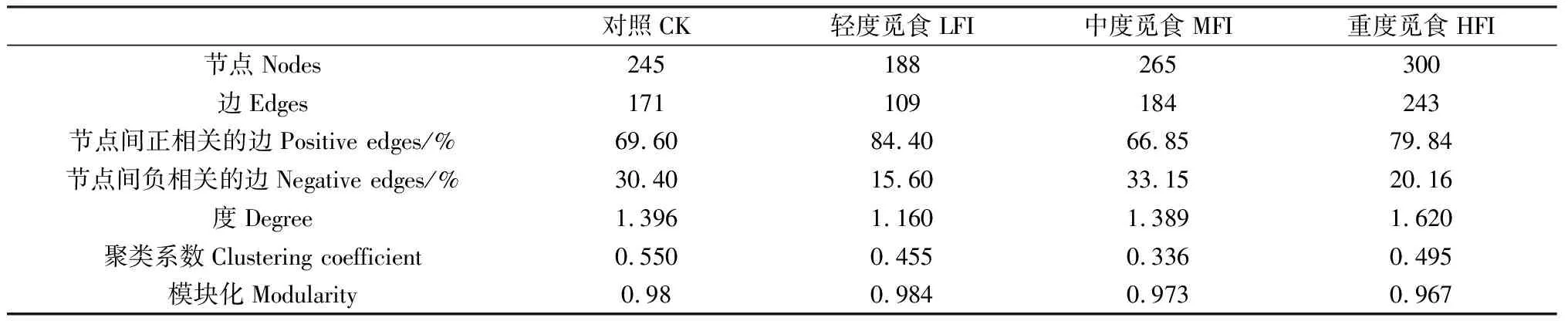
表2 細菌共現性網絡屬性

表3 共現性網絡屬性的Kolmogorov-Smirnov檢驗
2.5 影響土壤細菌群落的主要因子
為了確定牦牛覓食過程中對細菌群落特征變化發揮主要作用的環境因子,用細菌群落的Bray-Curtis距離矩陣與土壤理化參數的歐式距離矩陣進行Mantel test相關性檢驗,如圖5所示,pH值為影響細菌群落最顯著的環境因子(P<0.01)。此外,基于細菌OTU的Bray-Curtis相異度矩陣和細菌共現性網絡的度分別與pH值變化進行線性回歸分析(圖6)。結果表明,隨著土壤pH值的升高,細菌群落組成(P<0.001)和共現性網絡復雜度(P<0.05)都顯著增加,進一步驗證了上述結果。
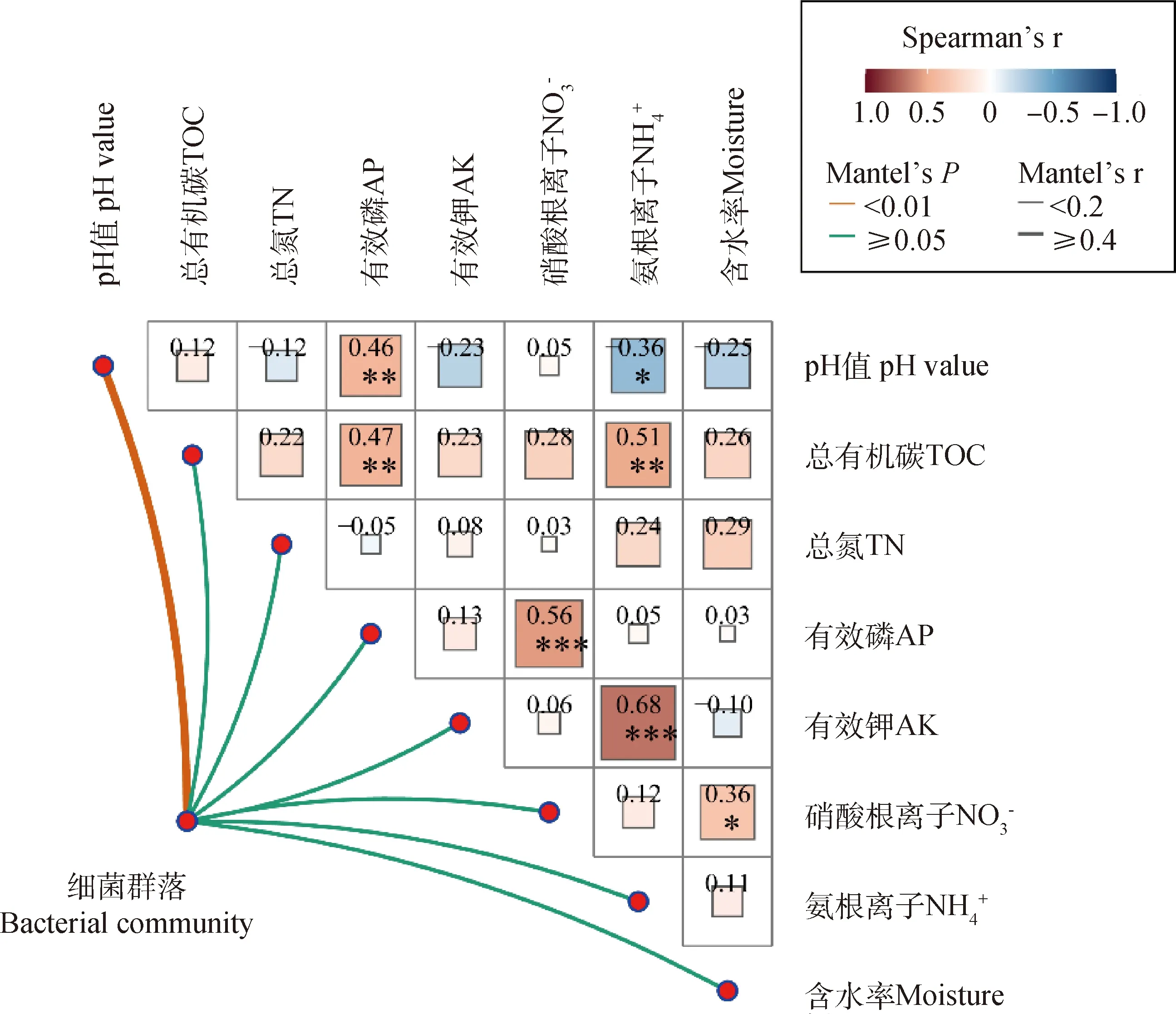
圖5 細菌群落與理化性質的相關性
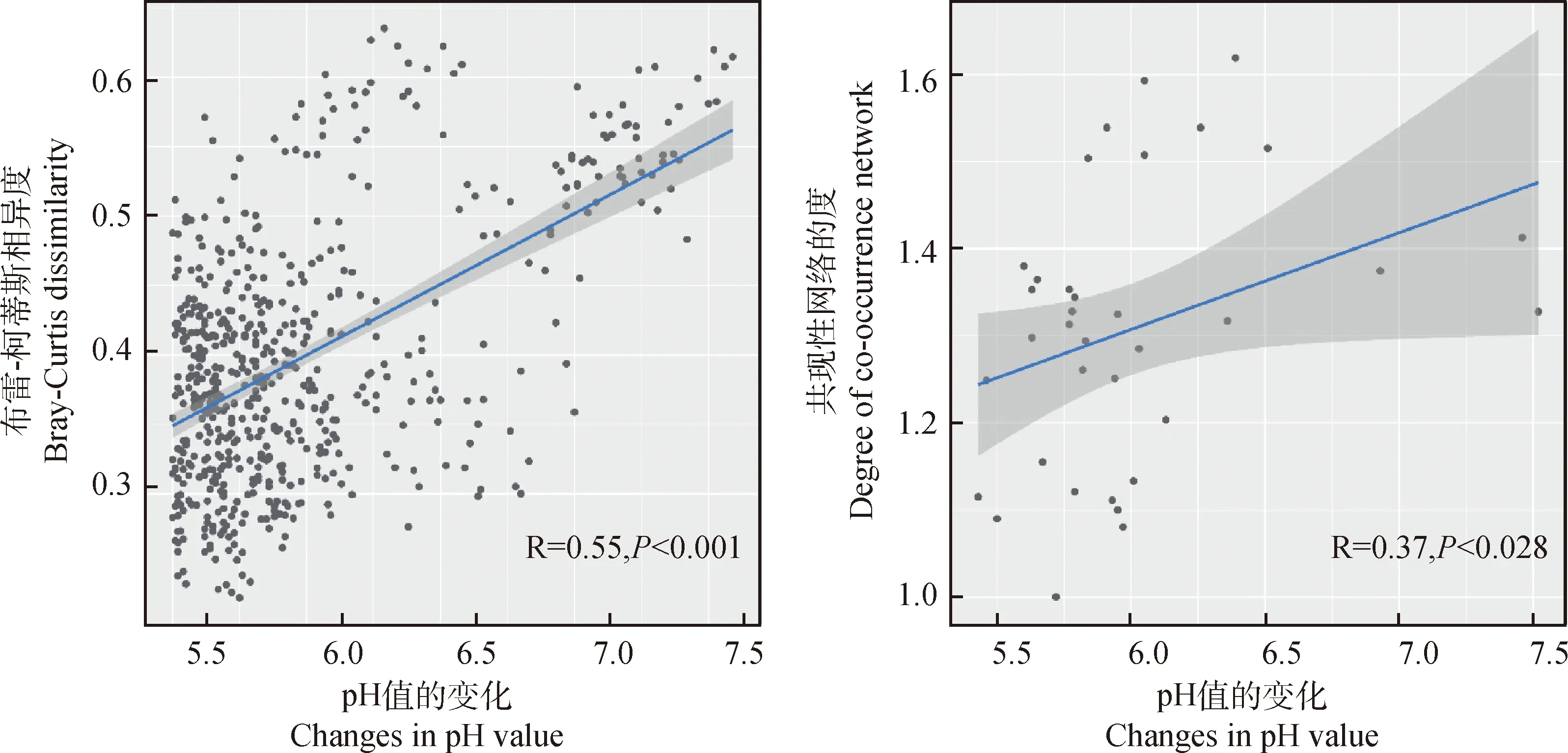
圖6 細菌群落的Bray-Curtis相異度和共現性網絡與pH之間的線性回歸
3 討論
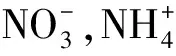
牦牛覓食行為擴大了細菌共現性網絡規模,增強了物種間的相互作用,提高了網絡復雜度。網絡復雜度的增加會導致網絡更加穩定,從而抵抗外界環境變化[43]。這可能與牦牛覓食過程中產生的排泄物有關。草地生態系統中,牲畜排泄物排泄到地表后,通過降水、踐踏和糞食性動物活動等作用進入土壤,造成土壤物理和化學等性質的改變,從而改變土壤微生物群落[44]。吳憲等[45]同樣表明向土壤中施用外源添加劑能夠擴大土壤共現性網絡規模,提高網絡復雜度與穩定性。屬于疣微菌門的節點受牦牛覓食干擾而增加,暗示著疣微菌門類群可能通過其它類群之間的相互作用調控高寒草甸土壤氮循環。此外,牦牛覓食行為增加了細菌類群之間正相關關系,表明適當的牦牛覓食行為能夠增強細菌類群間的協同作用,協同作用增強有利于形成更多營養級,使土壤養分利用更高效,從而能夠提高細菌群落物質循環、能量流動和信息傳遞的能力[46]。此外,牦牛覓食干擾導致共現性網絡平均聚類系數降低,意味著覓食干擾能夠增加土壤異質性,從而滿足不同生態位細菌的生長[47],而CK中較高的平均聚類系數代表土壤細菌功能的高度一致性。因此,合適的覓食強度有利于擴大網絡規模,提高網絡復雜性與穩定性,促使共現性網絡發揮更多的功能來適應外界環境變化。
我們發現土壤pH值是影響高寒草甸生態系統土壤細菌群落的主要因子。有研究表明,pH值的變化能夠改變土壤酶活性和離子濃度等特征,從而影響硝化和反硝化等功能細菌的代謝活性[48]。此外,土壤細菌類群更偏好偏堿性土壤,pH值升高能夠增加碳礦化和產甲烷等功能細菌的相對豐度[49]。我們發現,pH值升高引起細菌群落組成和共現性網絡復雜度顯著增加,從而可能影響高寒草甸土壤生態系統功能[50]。
4 結論
在高寒草甸,牦牛覓食改變了土壤細菌群落組成,顯著提高了疣微菌門(Verrucomicrobia)和芽單胞菌門(Gemmatimonadetes)的豐度。牦牛覓食干擾主要通過改變土壤pH值影響細菌群落組成,從而提高共現性網絡復雜度與穩定性。因此,適度的牦牛覓食干擾對改善高寒草甸土壤生態系統的可持續利用不可忽視。
